
Summary
-
1.
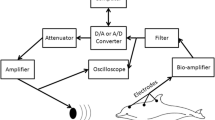
A preparation has been developed for the study of central auditory neurophysiology in cetaceans. Under N2O and Fluothane anesthesia, supplemented by succinylcholine chloride, twenty-nine specimens of four species of dolphins, most commonly Stenella caeruleo-alba, were stimulated with air-borne sound, water-borne sound, and directly by hydrophone pressed against the skin of the head. Evoked potential responses were recorded chiefly from the inferior colliculi but also from medullary auditory centers and the medial geniculate.
-
2.
The recorded potentials were onset responses of complicated waveform, usually having components with latencies of 2–4 msec. The longer latency responses probably represent heterogenous input to the inferior colliculi and, in a few cases, activity of collicular cells or input to the medial geniculate body. Shorter latency responses probably represent output of the cochlear nuclei and activity of medullary auditory nuclei.
-
3.
Although the evoked responses do not reflect increases in stimulus duration beyond a fraction of 1 msec, response threshold and waveform are sensitive to changes in rise time between 0.1 and 1.0 msec, and the small duration necessary to provide the onset response (as little as 0.1 msec) is also sufficient to provide accurate information about the frequency of the stimulus.
-
4.
Evoked potential responses in certain locations and intensity ranges were found to change markedly in amplitude or waveform with intensity changes as small as 1 db. The same response in other intensity ranges might be little altered by intensity changes as large as 10 db.
-
5.
Measurements of sensitivity as a function of frequency varied with electrode location, but averages or determinations of maximum sensitivity at each frequency at any location in a given animal produced audiograms in close agreement with that obtained behaviorally in Tursiops truncatus by Johnson (1966). Sensitivity was maximum at about 60 kc/s, and high between 20 and 70 kc/s; it typically fell about 25 db between 20 and 10 kc/s, and the same amount between 60 and 100 kc/s. The highest frequency eliciting a response was 120–140 kc/s. Data from the four species were not noticeably different.
-
6.
Response waveform and amplitude were often changed much more dramatically than was threshold by small changes in stimulus frequency, emphasizing the heterogeneity of input to a given electrode location, and the sharpness of frequency discrimination. Changes in threshold of up to 3 db/percent change in frequency were seen. Actual frequency discrimination is undoubtedly much sharper than these evoked potential measurements would indicate.
-
7.
The masking effect of background tones was shown to be confined to a relatively narrow band of frequencies surrounding that of the stimulus. Changes in masking effectiveness reached 30 db/10 kc/s change in background tone frequency. This too provides a conservative estimate of response “tuning”, since the evoked potentials represent the summed activity of such a large population of different units.
-
8.
Frequency modulated (FM) pulses of 2–5 msec duration and gradual rise and fall times were found sometimes to produce unexpectedly large responses compared with the response to any constant frequency (CF) pulse within the swept range. The response waveforms differed sharply with changes in starting frequency or range swept, even when the maximum response amplitude remained unchanged. In certain cases an FM pulse was found to be up to 10 db more effective if swept upward than if swept downward, and vice versa. It seems probable that significant populations of cells exist that are specifically responsive to sounds having certain FM characteristics, or certain combinations of frequencies in a given temporal order.
-
9.
Temporal resolution of successive sounds by cetaceans was found to be extremely rapid; virtually complete recovery of responsiveness in the fastest case took 0.8–1.0 msec between clicks; some initial recovery was often visible at approximately 0.5 msec. Following of a train of click stimuli was seen at repetition rates up to 2,000/sec. However, more typical recordings showed complete recovery of the collicular evoked potential required 3–5 msec. There was no evidence of facilitation of response to the second of two identical stimuli, as has been observed in echolocating bats.
-
10.
Distortion of the sound field by holding a sheet of paper between the source and the lower jaw or melon can in some recording sites cause dramatic changes in response waveform without significantly affecting the maximum response amplitude.
-
11.
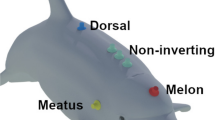
The primary pathway of sound to the cochlea is via the lower jaw. When stimuli were applied either with a loudspeaker in air, a hydrophone pressed against various points on the head, or a hydrophone underwater, sensitivity was consistently greatest over a limited portion of the side of the contralateral lower jaw, nearly as high over the ipsilateral side of the melon, markedly lower over the side of the head including the external ear orifice, and almost nil at the rostrum. This map was confirmed by using a distant sound source and local acoustical shields of 10×12 cm pieces of foam rubber or paper; held over the sensitive part of the lower jaw or melon this caused marked attenuation of response. At low frequencies (30 kc/s) the same pattern was found, but sensitivity was somewhat more uniform, relatively greater over the side of the head including the external auditory meatus and reduced on the melon, compared with the high-frequency pattern (60 kc/s).
-
12.
Gentle stroking of the side of the jaw or splashing of water on the jaw masked response to sounds. The effective input is acoustic. The effective area is circumscribed and identical to that for sound. Electrical stimulation of the skin of the jaw elicited no response in auditory centers and did not mask responses to sounds.
-
13.
A cone of best reception is directed forward and downward, some 5-30° from the midline in the horizontal plane (the higher the frequency the closer to the midline), and 5–20° below the horizontal in the optimum vertical plane. Sensitivity fell sharply (up to 1.5 db/degree) at more lateral angles. Sound coming from above the horizontal is probably much reduced in effectiveness.
-
14.
None of the response parameters studied was found to be affected by the anesthetics used, or by anesthetic doses of Nembutal.
-
15.
The evoked response characteristics are evaluated from the standpoint of adaptations for echolocation, and compared with analagous responses in bats and non-echolocating mammals. Possible mechanisms of distance measurement are considered. Target localization is discussed in light of the patterns of angular sensitivity resulting from use of the lower jaws as primary pathways of sound to the ears. The question is raised: In view of the extreme sensitivity of the head surface to mechanical disturbance, especially around the lower jaw, is echolocation usable during natural activity, e.g. rapid swimming, pursuit of prey, or food mastication ? If so, what adaptations have occurred to help overcome the potential masking noise ?
Similar content being viewed by others
References
Busnel, R. G., et A. Dziedzic: Resultats metrologiques experimentaux de l'echolocation chez le Phocaena phocaena et leur comparison avec ceux de certaines chauves-souris. In: Animal Sonar Systems, ed. R. G. Busnel, p. 307–335. Gap, France: Imprimerie Louis-Jean 1967.
Evans, W. E., and B. A. Powell: Discrimination of different metallic plates by an echolocating Delphinid. In: animal Sonar Systems, ed. R. G. Busnel, p. 363–383. Gap, France: Imprimerie Louis-Jean 1967.
Evans, W. E., W. W. Sutherland, and R. G. Beil: The directional characteristics of delphinid sounds. In: Marine Bioacoustics, ed. W. N. Tavolga, New York: Pergamon Press 1964.
Friend, J. H., N. Suga, and R. A. Suthers: Neural response in the inferior colliculus of echolocating bats to artificial orientation sounds and echoes. J. Cell Physiol. 67, 319–332 (1966).
Grinnell, A. D.: The neurophysiology of audition in bats: intensity and frequency parameters. J. Physiol. (Lond.) 167, 38–66 (1963a).
—: The neurophysiology of audition in bats: directionality and binaural interaction. J. Physiol. (Lond.) 167, 67–96 (1963b).
—: The neurophysiology of audition in bats: directionality and binaural interaction. J. Physiol. (Lond.) 167, 97–113 (1963c).
—: The neurophysiology of audition in bats: resistance to interference. J. Physiol. (Lond) 167, 114–127 (1963d).
—: Mechanisms of overcoming interference in echolocating animals. In: Animal Sonar Systems, ed. R. G. Busnel, p. 451–480. Gap, France: Imprimerie Louis-Jean 1967.
—, and V. S. Grinnell: Neural correlates of vertical localization by echo-locating bats. J. Physiol. (Lond.) 181, 830–851 (1965).
Haan, F. de, and W. Reysenbach: Listening underwater: thoughts on sound and cetacean hearing. In: Whales, Dolphins and Porpoises, ed. K. S. Norris, p. 583–596. Berkeley: University of California Press 1966.
Henson jr., O. W.: The activity and function of the middle ear muscles in echolocating bats. J. Physiol. (Lond.) 180, 871–887 (1965).
: The perception and analysis of biosonar signals by bats. In: Animal Sonar System, ed. R. G. Busnel, p. 949–1003. Gap, France: Imprimerie Louis-Jean 1967.
Johnson, C. S.: Auditory thresholds of the bottlenosed porpoise (Tursiops truncatus, Montagu). NOTS Technical Publication 4178 (1966).
Kramer, M. O.: The dolphin's secret. New Scientist 7, 1118–1120 (1960a).
— Boundary layer stabilization by distributed damping. J. Amer. Soc. Naval Engin. Inc., Feb. 1960, 25–33 (1960b).
Kuhl, W., G. R. Sohodder, and F. K. Schröder: Condenser transmitters and microphones with solid dielectric for airborne ultrasonics. Acoustica 4, 519–532 (1954).
Nagel, E. L., P. J. Morgane, and W. L. McFarland: Anesthesia for the bottle nose dolphin, Tursiops truncatus. Science 146, 1591–1593 (1964).
Nelson, P. G., S. D. Erulkar, and J. S. Bryan: Responses of units of the inferior colliculus to time-varying acoustic stimuli. J. Neurophysiol. 29, 834–860 (1966).
Nordmark, J.: Perception of distance in animal echolocation. Nature (Lond.) 188, 1009–1010 (1960).
Norris, K. S.: Some problems of echolocation in cetaceans. In: Marine Bioacoustics, ed. W. N. Tavolga, p. 317–336. New York: Pergamon Press 1964.
— The evolution of acoustic mechanisms in odontocete cetaceans. (In press) (1967).
—, W. E. Evans, and R. N. Turner: Echolocation in an Atlantic bottlenos porpoise during discrimination. In: Animal Sonar Systems, ed. R. G. Busnel, p. 409–437. Gap, France: Imprimerie Louis-Jean 1967.
Ogawa, T., and S. Arifuku: On the acoustic system in the cetacean brains. Sci. Reps. Whale Res. Inst. 2, 1–20 (1948).
Palmer, E., and G. Weddell: The relationship between structure, innervation and function of the skin of the bottlenose dolphin (Tursiops truncatus). Proc. zool. Soc. London 143, 553–567 (1964).
Peake, W. T.: An analytical study of electrical responses at the periphery of the auditory system. Tech. Rep. 365, MIT Research Laboratory of Electronics (1960).
Pumphrey, R. J.: The sense organs of birds. The Ibis 90, 171–199 (1948).
Schevill, W. E., and B. Lawrence: Auditory response of a bottlenosed porpoise, Tursiops truncatus, to frequencies above 100 kc. J. exp. Zool. 124, 147–165 (1953).
Suga, N.: Single unit activity in cochlear nucleus and inferior colliculus of echolocating bats. J. Physiol. (Lond.) 172, 449–474 (1964a).
—: Recovery cycles and responses to frequency modulated tones pulses in auditory neurones of echo-locating bats. J. Physiol. (Lond.) 175, 50–80 (1964b).
—: Analysis of frequency-modulated sounds by auditory neurones of echo-locating bats. J. Physiol. (Lond.) 179, 26–53 (1965a).
—: Functional properties of auditory neurones in the cortex of echo-locating bats. J. Physiol. (Lond.) 181, 671–700 (1965b).
Turner, R. N., and K. S. Norris: Discriminating echolocating in a porpoise. J. exp. Anal. Behav. 9, 535–544 (1966).
Wainwright, W. N.: Comparison of hearing thresholds in air and water. J. acoust. Soc. Amer. (1958) 30, 1025–1029.
Whitfield, I. C., and E. F. Evans: Responses of auditory cortical neurons to stimuli of changing frequency. J. Neurophysiol. 28, 655–672 (1965).
Yamada, M.: On the hearing organ of cetacea. Juzen Igakukkai Zasshi 66, 1–15 (1960).
Yanagisawa, K., O. Sato, M. Nomoto, Y. Katsuki, E. Ikezono, A. D. Grinnell, and T. H. Bullock: Auditory evoked potentials from brain stem in cetaceans. Fed. Proc. 25, 464 (Abs.) (1966).
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Bullock, T.H., Grinnell, A.D., Ikezono, E. et al. Electrophysiological studies of central auditory mechanisms in cetaceans. Zeitschrift für vergleichende Physiologie 59, 117–156 (1968). https://doi.org/10.1007/BF00339347
Received:
Issue Date:
DOI: https://doi.org/10.1007/BF00339347