Evaluation of Popcorn Hybrids for Nitrogen Use Efficiency and Responsiveness

 ,
, 
 , , , ,
, , , ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions and Genotypes
2.2. Nitrogen Availability and Studied Traits
2.3. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Variance
3.2. Mean Performance and Yield Stability of Genotypes
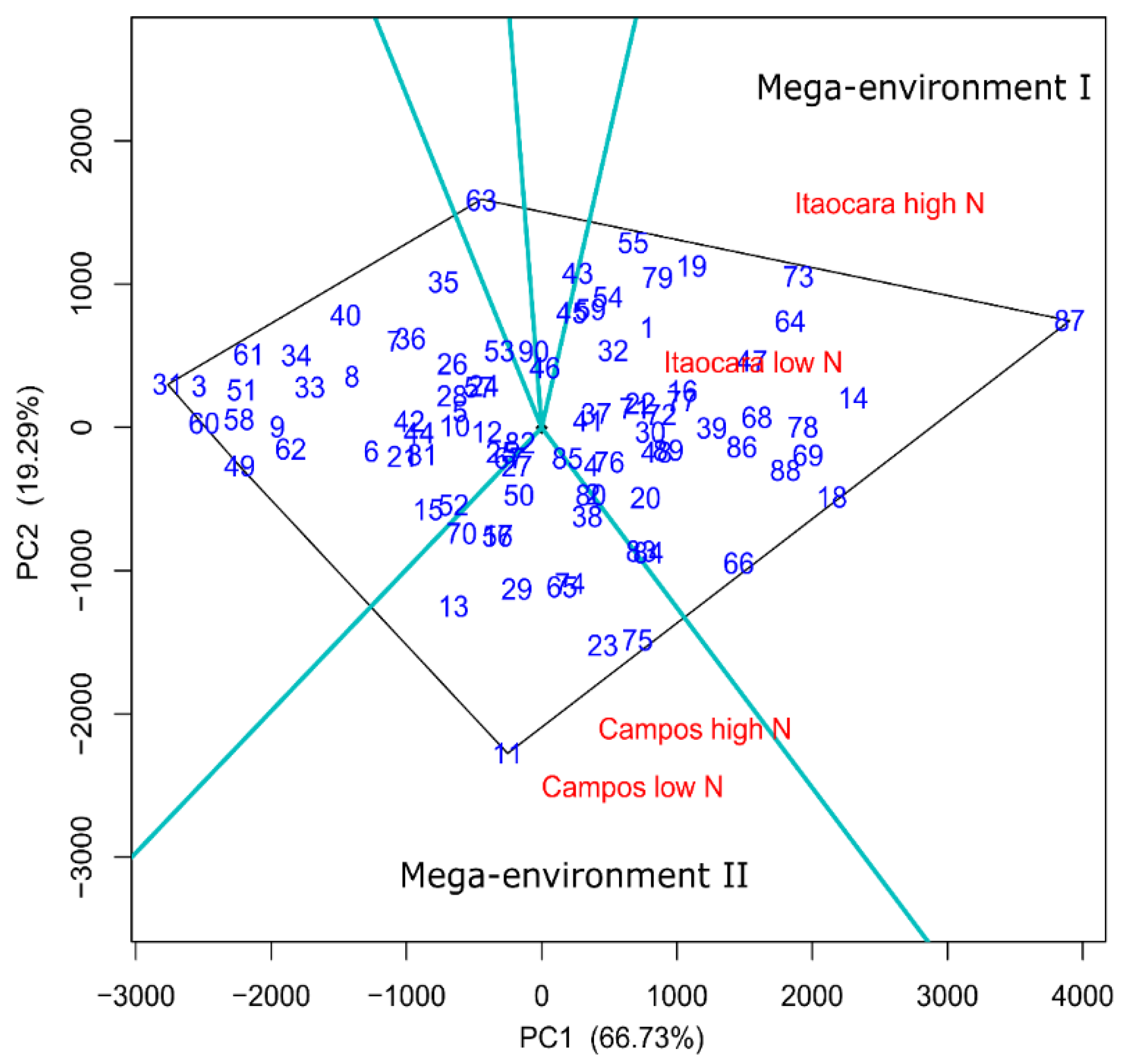
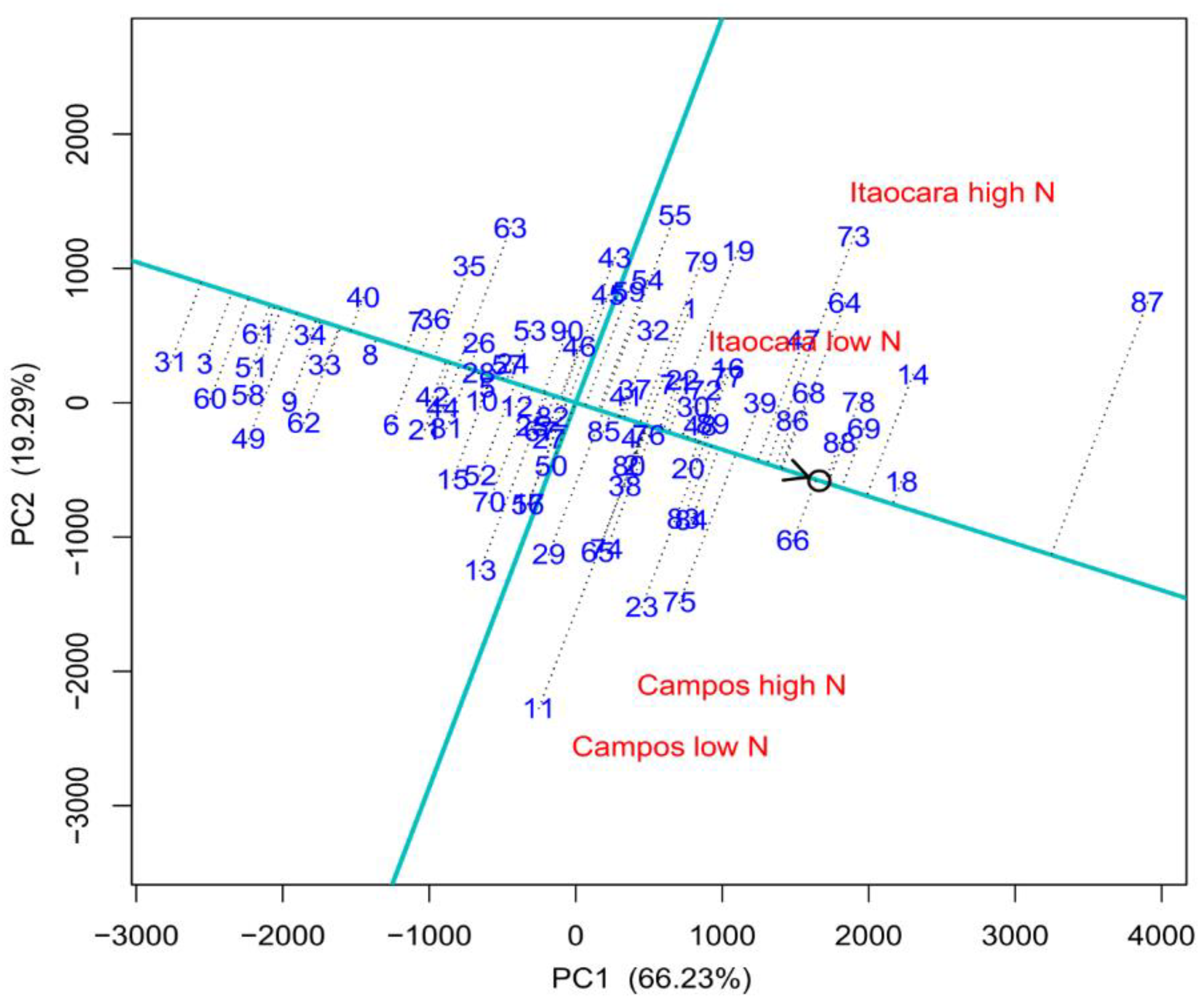
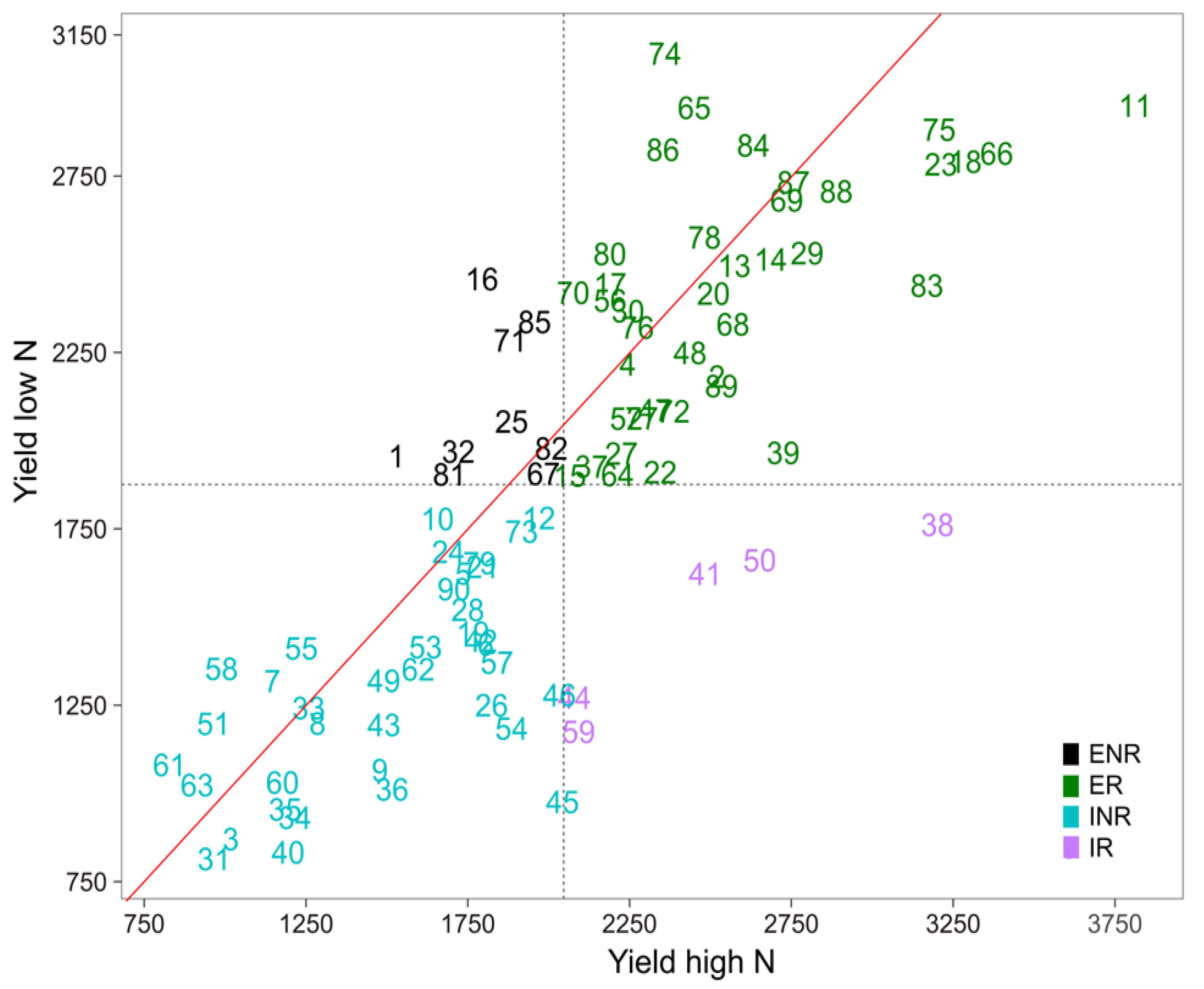
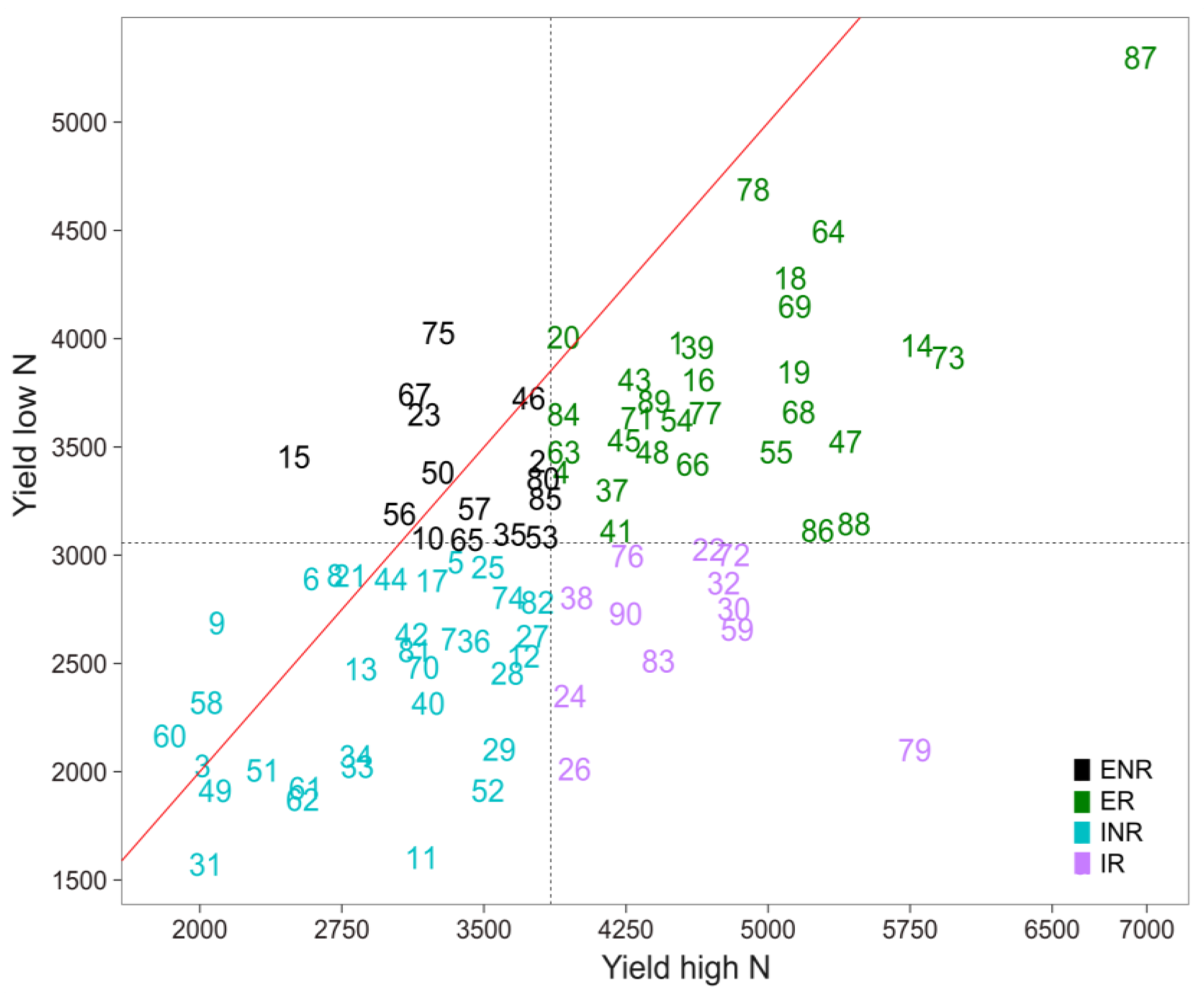
3.3. Selection for Efficient N Use
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Conant, R.T.; Berdanier, A.B.; Grace, P.R. Patterns and trends in nitrogen use and nitrogen recovery efficiency in world agriculture. Glob. Biogeochem. Cycles 2013, 27, 558–566. [Google Scholar] [CrossRef]
- Andrews, M.; Lea, P.J. Our nitrogen ‘footprint’: The need for increased crop nitrogen use efficiency. Ann. Appl. Biol. 2013, 163, 165–169. [Google Scholar] [CrossRef]
- Sharma, L.; Bali, S. A Review of Methods to Improve Nitrogen Use Efficiency in Agriculture. Sustainability 2017, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Raun, W.R.; Johnson, G.V. Improving Nitrogen Use Efficiency for Cereal Production. Agron. J. 1999, 91, 357. [Google Scholar] [CrossRef] [Green Version]
- Kant, S.; Bi, Y.-M.; Rothstein, S.J. Understanding plant response to nitrogen limitation for the improvement of crop nitrogen use efficiency. J. Exp. Bot. 2011, 62, 1499–1509. [Google Scholar] [CrossRef] [Green Version]
- Good, A.G.; Beatty, P.H. Fertilizing Nature: A Tragedy of Excess in the Commons. PLoS Biol. 2011, 9, e1001124. [Google Scholar] [CrossRef]
- Hirel, B.; Le Gouis, J.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef]
- Fischer, J.J.; Beatty, P.H.; Good, A.G.; Muench, D.G. Manipulation of microRNA expression to improve nitrogen use efficiency. Plant Sci. 2013, 210, 70–81. [Google Scholar] [CrossRef]
- dos Santos, A.; do Amaral Júnior, A.T.; Kurosawa, R.d.N.F.; Gerhardt, I.F.S.; Fritsche Neto, R. GGE Biplot projection in discriminating the efficiency of popcorn lines to use nitrogen. Ciência e Agrotecnologia 2017, 41, 22–31.
- Frutos, E.; Galindo, M.P.; Leiva, V. An interactive biplot implementation in R for modeling genotype-by-environment interaction. Stoch. Environ. Res. Risk Assess. 2014, 28, 1629–1641. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Kingston, ON, Canada, 2014. [Google Scholar]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE Biplot vs. AMMI Analysis of Genotype-by-Environment Data. Crop Sci. 2007, 47, 643. [Google Scholar] [CrossRef]
- Rajcan, I.; Tollenaar, M. Source:sink ratio and leaf senescence in maize. Field Crop. Res. 1999, 60, 255–265. [Google Scholar] [CrossRef]
- Vetsch, J.A.; Randall, G.W. Corn Production as Affected by Nitrogen Application Timing and Tillage. Agron. J. 2004, 96, 502. [Google Scholar] [CrossRef] [Green Version]
- Miranda, G.V.; Godoy, C.L.; Galvão, J.C.C.; Santos, I.C.; Eckert, F.R.; Souza, L.V. Selection of discrepant maize genotypes for nitrogen use efficiency by a chlorophyll meter. Crop. Breed. Appl. Biotechnol. 2005, 5, 451–459. [Google Scholar] [CrossRef]
- Bullock, D.G.; Anderson, D.S. Evaluation of the Minolta SPAD-502 chlorophyll meter for nitrogen management in corn. J. Plant Nutr. 1998, 21, 741–755. [Google Scholar] [CrossRef]
- Soares, M.O.; Miranda, G.V.; Guimarães, L.J.M.; Marriel, I.E.; Guimarães, C.T. Parâmetros genéticos de uma população de milho em níveis contrastantes de nitrogênio. Rev. Ciência Agronômica 2011, 42, 168–174. [Google Scholar] [CrossRef]
- Udo, E.F.; Ajala, S.O.; Olaniyan, A.B. Physiological and morphological changes associated with recurrent selection for low nitrogen tolerance in maize. Euphytica 2017, 213, 140. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crop. Res. 1996, 48, 65–80. [Google Scholar] [CrossRef]
- Ngirazi, S.; Manjeru, P.; Ncube, O. Pod yield stability and adaptation of groundnut (Arachis hypogaea L.) genotypes evaluated in multi-environmental trials in Zimbabwe. Afr. J. Plant Sci. 2017, 11, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Bornhofen, E.; Benin, G.; Storck, L.; Woyann, L.G.; Duarte, T.; Stoco, M.G.; Marchioro, S.V. Statistical methods to study adaptability and stability of wheat genotypes. Bragantia 2017, 76, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Milioli, A.S.; Zdziarski, A.D.; Woyann, L.G.; dos Santos, R.; Rosa, A.C.; Madureira, A.; Benin, G. Yield stability and relationships among stability parameters in soybean genotypes across years. Chil. J. Agric. Res. 2018, 78, 299–309. [Google Scholar] [CrossRef]
- Almeida, V.C.; Viana, J.M.S.; Risso, L.A.; Ribeiro, C.; DeLima, R.O. Generation mean analysis for nitrogen and phosphorus uptake, utilization, and translocation indexes at vegetative stage in tropical popcorn. Euphytica 2018, 214, 103. [Google Scholar] [CrossRef]
- Stromberger, J.A.; Tsai, C.Y.; Huber, D.M. Interactions of potassium with nitrogen and their influence on growth and yield potential in maize. J. Plant Nutr. 1994, 17, 19–37. [Google Scholar] [CrossRef]
- McCullough, D.E.; Mihajlovic, M.; Aguilera, A.; Tollenaar, M.; Girardin, P. Influence of N supply on development and dry matter accumulation of an old and a new maize hybrid. Can. J. Plant Sci. 1994, 74, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Lithourgidis, A.; Dordas, C.; Damalas, C.; Vlachostergios, D. Annual intercrops: An alternative pathway for sustainable agriculture. Aust. J. Crop Sci. 2011, 5, 396–410. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Itaocara, RJ | ||||||||||
| Layer | pH | P | K | Ca | Mg | Al | H + Al | Na | C | OM |
| H2O | Mg dm−3 | ............................mmol dm−3............................ | g dm−3 | |||||||
| 0–10 cm | 5.1 | 5.0 | 3.7 | 11.9 | 8.0 | 1.2 | 18.2 | 1.1 | 8.9 | 15.3 |
| 10–20 cm | 4.9 | 2.0 | 1.9 | 11.9 | 7.2 | 1.8 | 17.8 | 0.6 | 9.3 | 16.0 |
| Campos dos Goytacazes, RJ | ||||||||||
| Layer | pH | P | K | Ca | Mg | Al | H + Al | Na | C | OM |
| H2O | mg dm−3 | ............................mmol dm−3............................ | g dm−3 | |||||||
| 0–10 cm | 4.8 | 22.0 | 2.6 | 16.3 | 8.5 | 1.2 | 21.4 | 12.3 | 8.9 | 15.3 |
| 10–20 cm | 5.0 | 10.0 | 1.7 | 15.6 | 7.4 | 1.5 | 18.3 | 10.3 | 9.3 | 16.0 |
| Hybrid | Parents | Hybrid | Parents | Hybrid | Parents |
|---|---|---|---|---|---|
| 1 | L54 × L59 | 31 | L75 × L76 | 61 | L80 × P2 |
| 2 | L54 × L61 | 32 | L75 × L77 | 62 | L80 × P6 |
| 3 | L54 × L75 | 33 | L75 × L80 | 63 | L80 × P7 |
| 4 | L54 × L76 | 34 | L75 × P2 | 64 | P2 × L54 |
| 5 | L54 × L77 | 35 | L75 × P6 | 65 | P2 × L59 |
| 6 | L54 × L80 | 36 | L75 × P7 | 66 | P2 × L61 |
| 7 | L54 × P2 | 37 | L76 × L54 | 67 | P2 × L75 |
| 8 | L54 × P6 | 38 | L76 × L59 | 68 | P2 × L76 |
| 9 | L54 × P7 | 39 | L76 × L61 | 69 | P2 × L77 |
| 10 | L59 × L54 | 40 | L76 × L75 | 70 | P2 × L80 |
| 11 | L59 × L61 | 41 | L76 × L77 | 71 | P2 × P6 |
| 12 | L59 × L75 | 42 | L76 × L80 | 72 | P2 × P7 |
| 13 | L59 × L76 | 43 | L76 × P2 | 73 | P6 × L54 |
| 14 | L59 × L77 | 44 | L76 × P6 | 74 | P6 × L59 |
| 15 | L59 × L80 | 45 | L76 × P7 | 75 | P6 × L61 |
| 16 | L59 × P2 | 46 | L77 × L54 | 76 | P6 × L75 |
| 17 | L59 × P6 | 47 | L77 × L59 | 77 | P6 × L76 |
| 18 | L59 × P7 | 48 | L77 × L61 | 78 | P6 × L77 |
| 19 | L61 × L54 | 49 | L77 × L75 | 79 | P6 × L80 |
| 20 | L61 × L59 | 50 | L77 × L76 | 80 | P6 × P2 |
| 21 | L61 × L75 | 51 | L77 × L80 | 81 | P6 × P7 |
| 22 | L61 × L76 | 52 | L77 × P2 | 82 | P7 × L54 |
| 23 | L61 × L77 | 53 | L77 × P6 | 83 | P7 × L59 |
| 24 | L61 × L80 | 54 | L77 × P7 | 84 | P7 × L61 |
| 25 | L61 × P2 | 55 | L80 × L54 | 85 | P7 × L75 |
| 26 | L61 × P6 | 56 | L80 × L59 | 86 | P7 × L76 |
| 27 | L61 × P7 | 57 | L80 × L61 | 87 | P7 × L77 |
| 28 | L75 × L54 | 58 | L80 × L75 | 88 | P7 × L80 |
| 29 | L75 × L59 | 59 | L80 × L76 | 89 | P7 × P2 |
| 30 | L75 × L61 | 60 | L80 × L77 | 90 | P7 × P6 |
| SV | DF | Mean Square | |||
|---|---|---|---|---|---|
| FF | FI | SPAD | GY | ||
| Block/Rep/N/L | 108 | 4.5 | 5.0 | 47.0 | 42,442.0 |
| Rep/N/Location | 8 | 16.5 | 6.0 | 59.0 | 55,746.0 |
| Location (L) | 1 | 853.3 ** | 3499.0 ** | 6932.0 ** | 603,308,664.0 ** |
| Nitrogen (N) | 1 | 0.1 | 58.0 * | 1387.0 ** | 62,989,885.0 ** |
| L × N | 1 | 16.6 * | 58.0 ** | 128.0 ** | 26,597,501.0 ** |
| Hybrid (H) | 89 | 45.9 ** | 15.0 ** | 99.0 ** | 4,222,957.0 ** |
| L × H | 89 | 6.9 ** | 7.0 ** | 36.0 | 1,564,010.0 ** |
| N × H | 89 | 5.9 ** | 4.0 * | 54.0 * | 628,677.0 ** |
| L × N × H | 89 | 3.9 | 4.0 * | 35.0 | 567,165.0 ** |
| Error | 604 | 3.1 | 3.0 | 35.0 | 37,787.0 |
| Accuracy | - | 0.965 | 0.895 | 0.803 | 0.996 |
| Traits | Campos dos Goytacazes | Itaocara | |
|---|---|---|---|
| FF | High N | 54.84 aA | 52.81 Ab |
| Low N | 54.6 aA | 53.07 aA | |
| % Reduction | 0.43 | −0.5 | |
| FI | High N | −0.70 aB | −3.84 bA |
| Low N | −0.72 aB | −4.77 aA | |
| % Reduction | −2.86 | −24.11 | |
| SPAD | High N | 52.18 aB | 56.56 aA |
| Low N | 49.22 bB | 54.98 bA | |
| % Reduction | 5.66 | 2.79 | |
| GY | High N | 2296.48 aB | 3855.16 aA |
| Low N | 1827.34 bB | 3058.29 bA | |
| % Reduction | 20.43 | 20.67 |
| Principal Component | Singular Value | % Cumulative | % Explained | IR |
|---|---|---|---|---|
| 1 | 16,756.03 | 66.73 | 66.73 | 2.67 |
| 2 | 4843.76 | 86.02 | 19.29 | 0.77 |
| 3 | 2249.87 | 94.98 | 8.96 | 0.36 |
| 4 | 1260.53 | 100.00 | 5.02 | 0.20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, A.d.; Amaral Júnior, A.T.d.; Kamphorst, S.H.; Gonçalves, G.M.B.; Santos, P.H.A.D.; Vivas, J.M.S.; Mafra, G.S.; Khan, S.; Oliveira, F.T.d.; Schmitt, K.F.M.; et al. Evaluation of Popcorn Hybrids for Nitrogen Use Efficiency and Responsiveness. Agronomy 2020, 10, 485. https://doi.org/10.3390/agronomy10040485
Santos Ad, Amaral Júnior ATd, Kamphorst SH, Gonçalves GMB, Santos PHAD, Vivas JMS, Mafra GS, Khan S, Oliveira FTd, Schmitt KFM, et al. Evaluation of Popcorn Hybrids for Nitrogen Use Efficiency and Responsiveness. Agronomy. 2020; 10(4):485. https://doi.org/10.3390/agronomy10040485
Chicago/Turabian StyleSantos, Adriano dos, Antônio Teixeira do Amaral Júnior, Samuel Henrique Kamphorst, Gabriel Moreno Bernardo Gonçalves, Pedro Henrique Araújo Diniz Santos, Janieli Maganha Silva Vivas, Gabrielle Sousa Mafra, Shahid Khan, Fabio Tomaz de Oliveira, Kátia Fabiane Medeiros Schmitt, and et al. 2020. "Evaluation of Popcorn Hybrids for Nitrogen Use Efficiency and Responsiveness" Agronomy 10, no. 4: 485. https://doi.org/10.3390/agronomy10040485