Effects of Different Environmental Factors on the Growth and Bioactive Substance Accumulation of Porphyridium purpureum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture
2.2. Different Growth Condition Treatments
2.3. Determination of Algae Growth
2.4. Determination of Chlorophyll Fluorescence
2.5. Determination of Polysaccharides Content
2.6. Determination of Phycoerythrin Content
2.7. Determination of Total Lipids Content
2.8. Statistical Analysis
3. Results
3.1. Algal Growth
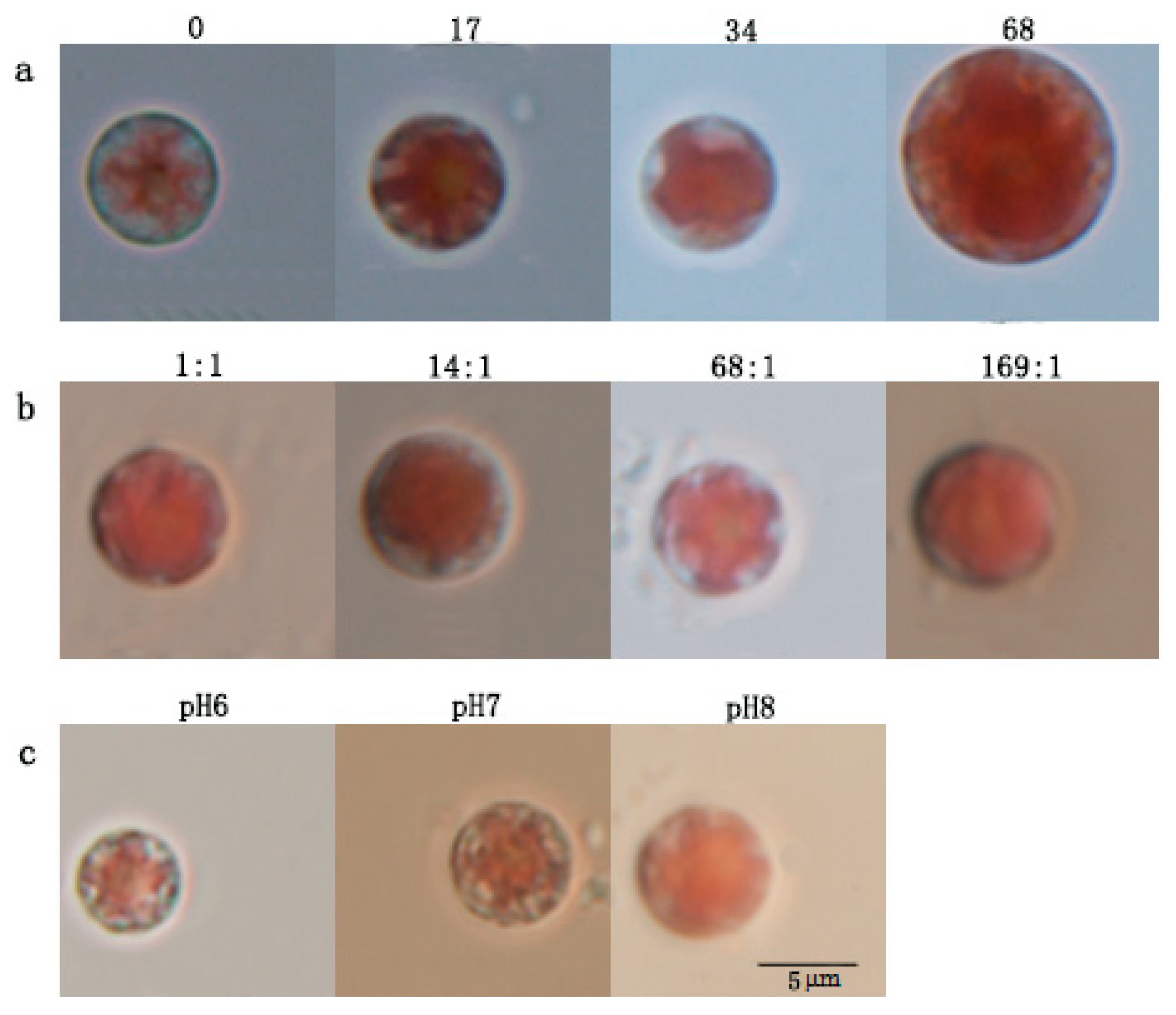
3.1.1. Microscopic Observation
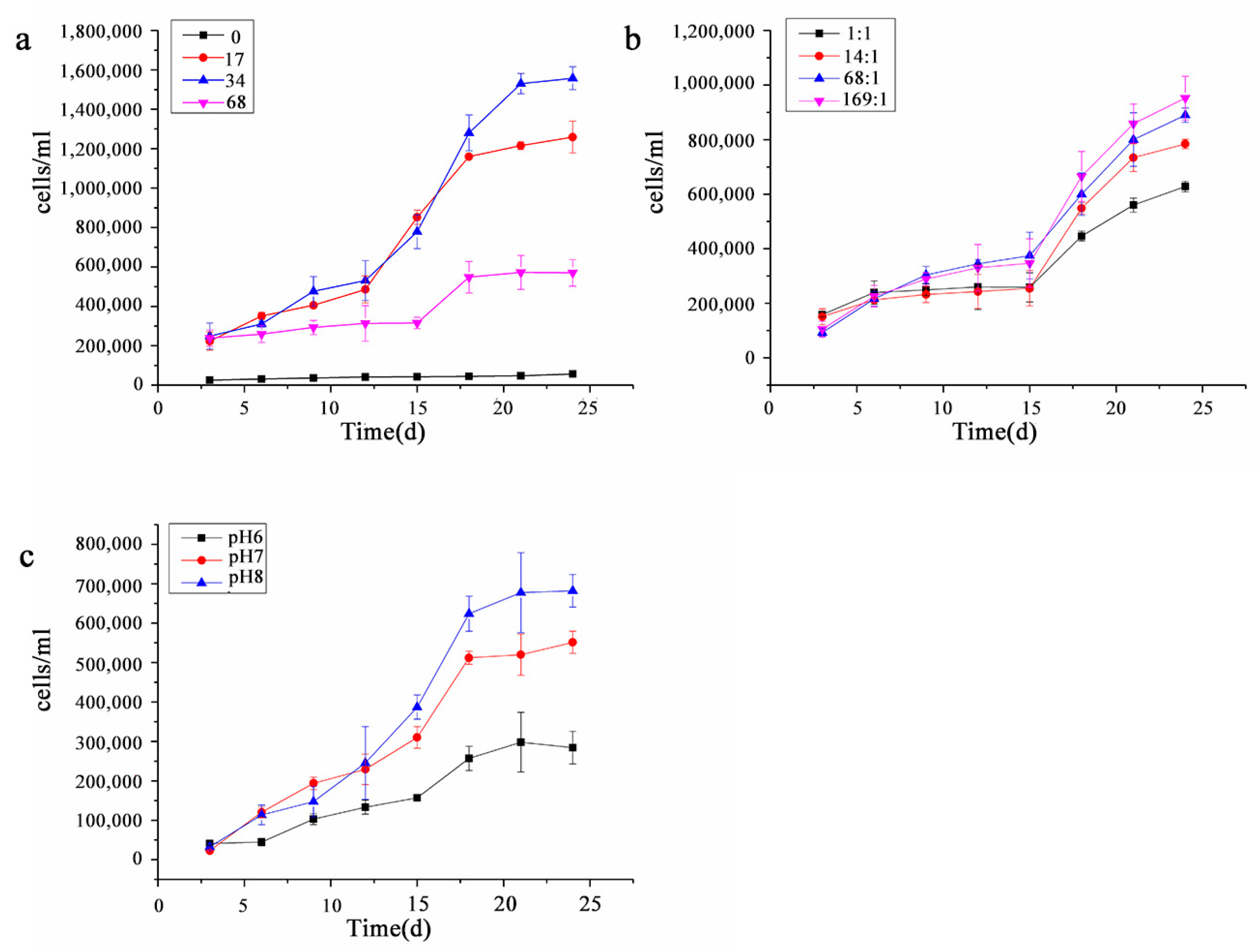
3.1.2. Growth Curve
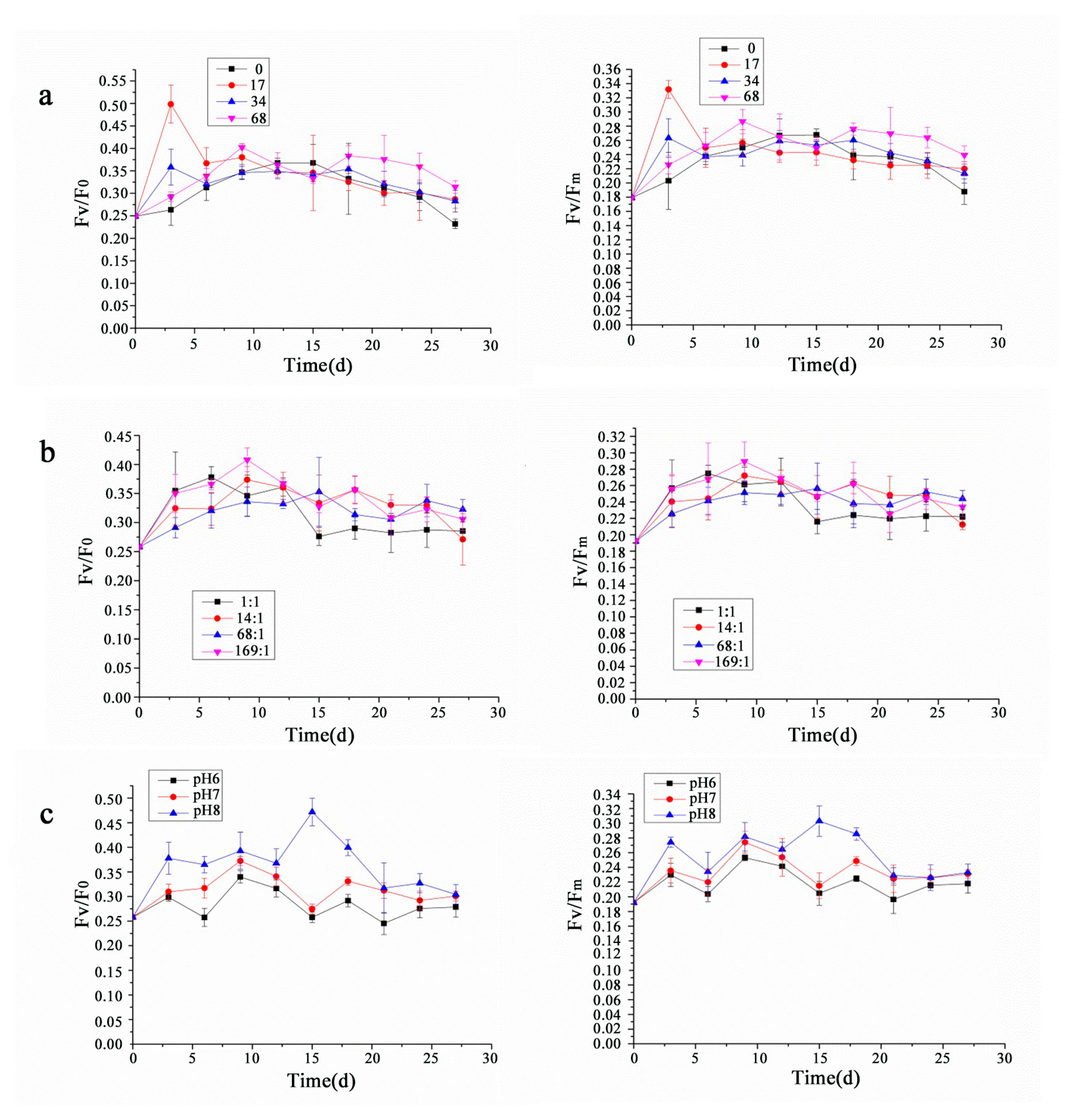
3.2. Analysis of Chlorophyll Fluorescence
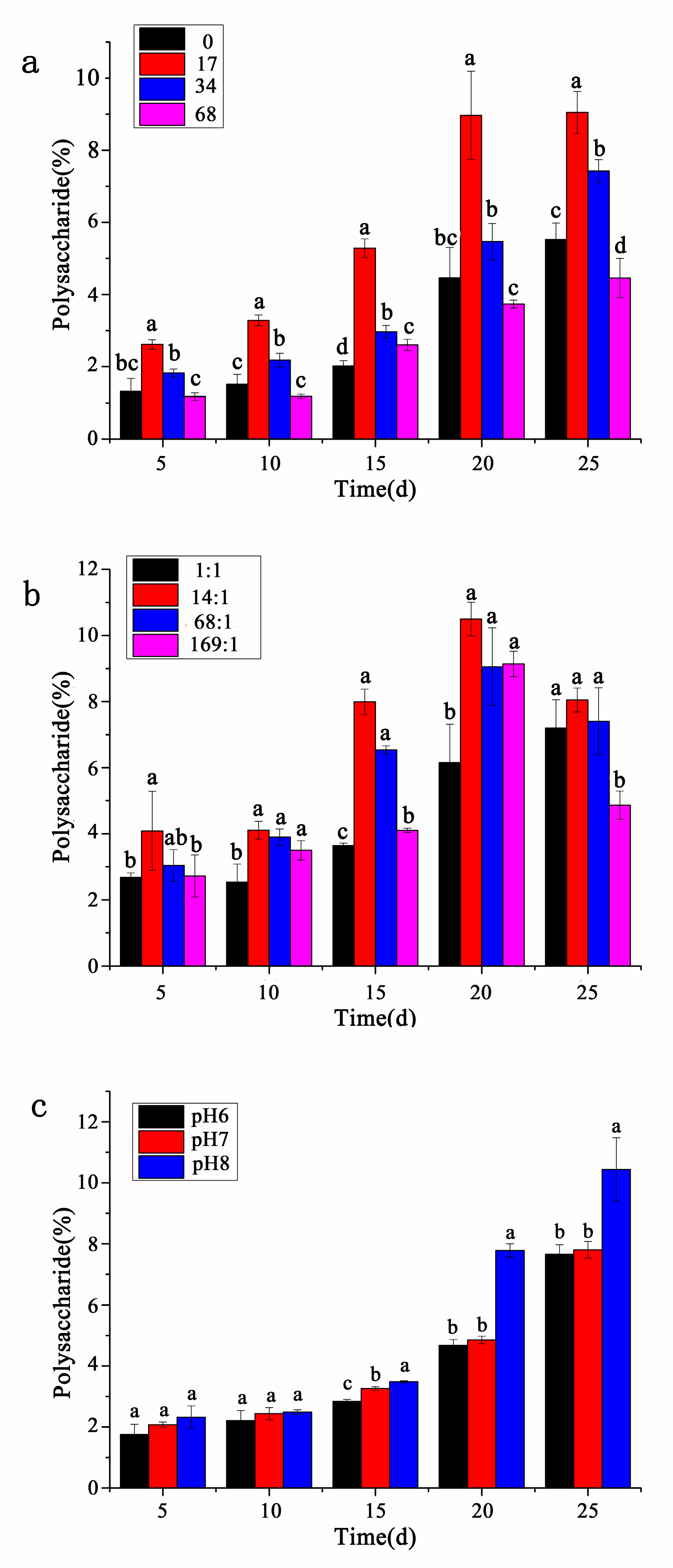
3.3. Contents of Polysaccharides
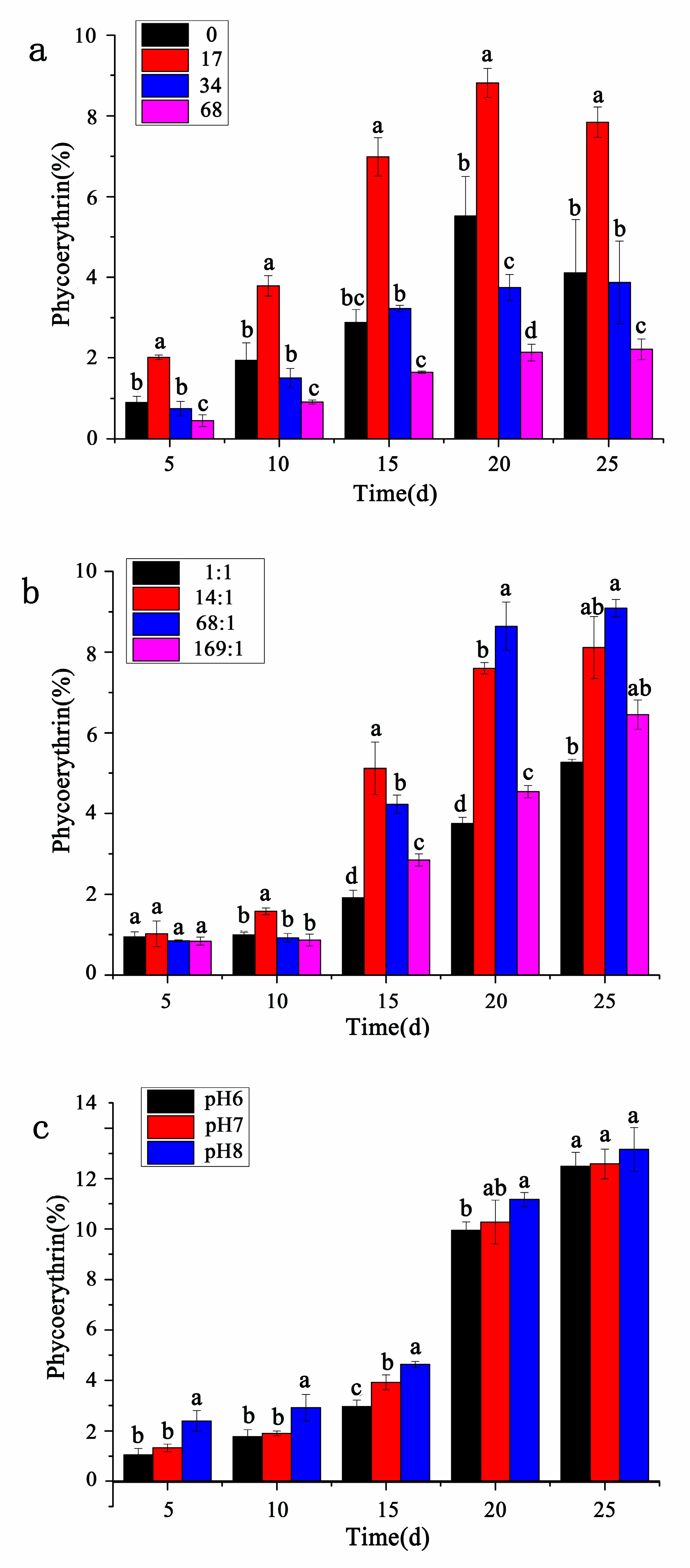
3.4. Contents of Phycoerythrin
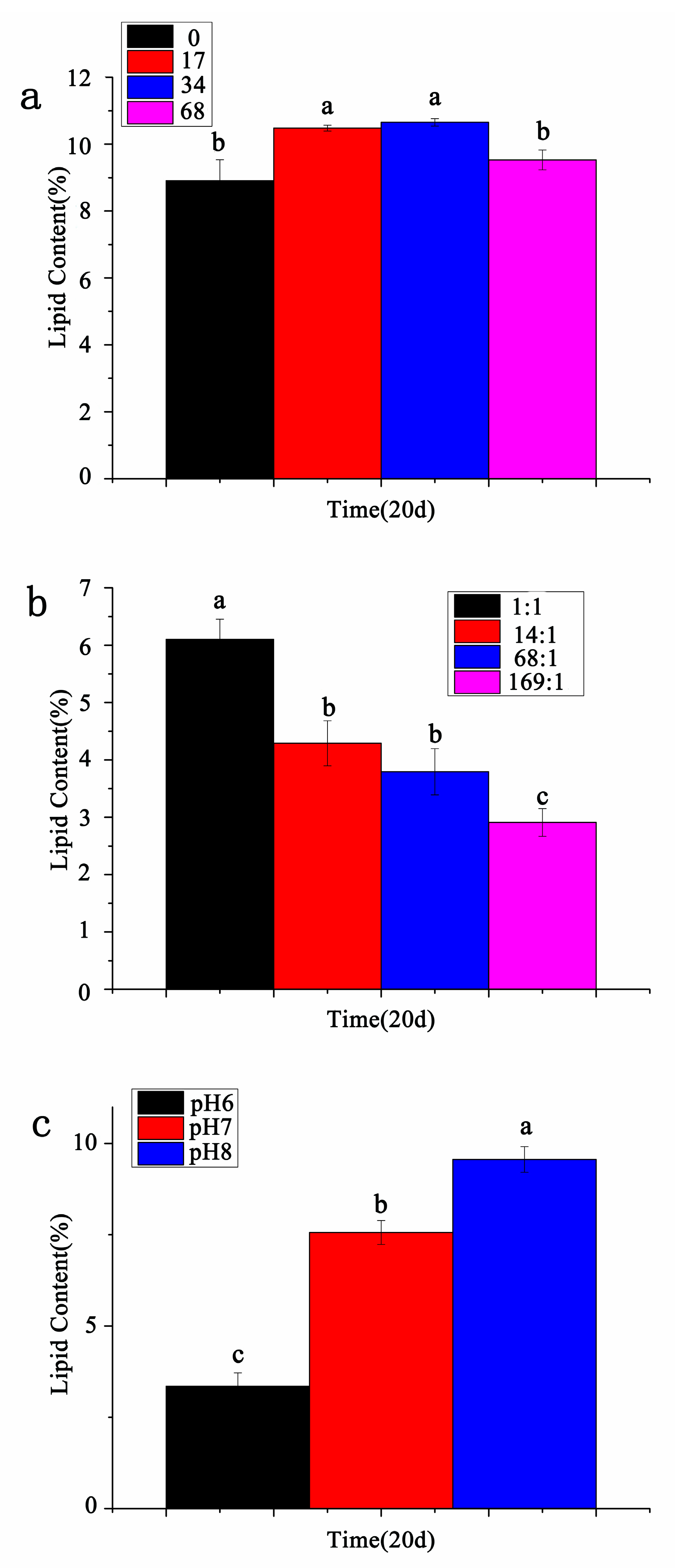
3.5. Contents of Total Lipids
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klein-Marcuschamer, D.; Chisti, Y.; Benemann, J.R.; Lewis, D. A matter of detail: Assessing the true potential of microalgal biofuels. Biotechnol. Bioeng. 2013, 110, 2317–2322. [Google Scholar]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Cheminform 2003, 83, 1–11. [Google Scholar] [CrossRef]
- Kim, K.H.; Choi, I.S.; Kim, H.M.; Wi, S.G. Bioethanol production from the nutrient stress-induced microalga Chlorella vulgaris by enzymatic hydrolysis and immobilized yeast fermentation. Bioresour. Technol. 2014, 153, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Nigam, P.S.; Murphy, J.D. Mechanism and challenges in commercialisation of algal biofuels. Bioresour. Technol. 2011, 102, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Owende, P. Biofuels from microalgae: Towards meeting advanced fuel standards. In Advanced Biofuels and Bioproducts; Lee, J., Ed.; Springer: New York, NY, USA, 2013; pp. 553–599. [Google Scholar]
- Gaignard, C.; Gargouch, N.; Dubessay, P.; Delattre, C.; Pierre, G. New horizons in culture and valorization of red microalgae. Biotechnol. Adv. 2019, 37, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Thoisen, C.; Hansen, B.W.; Nielsen, S.L. A simple and fast method for extraction and quantification of cryptophyte phycoerythrin. MethodsX 2017, 4, 209–213. [Google Scholar] [CrossRef]
- Arad, S.; Levy-Ontman, O. Red microalgal cell-wall polysaccharidess: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef]
- Ramus, J.; Kenney, B.E. Shear degradation as a probe of microalgal exopolymer structure and rheological properties. Biotechnol. Bioeng. 1989, 34, 1203–1208. [Google Scholar] [CrossRef]
- Dvir, I.; Chayoth, R.; Sod-Moriah, U.; Shany, S.; Nyska, A. Soluble polysaccharides and biomass of red microalga Porphyridium sp. alter intestinal morphology and reduce serum cholesterol in rats. Br. J. Nutr. 2000, 84, 469–476. [Google Scholar] [CrossRef] [Green Version]
- González, M.E.; Alarcón, B.; Carrasco, L. Polysaccharidess as antiviral agents: Antiviral activity of carrageenan. Antimicrob. Agents Chemother. 1987, 31, 1388–1393. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, I.; Itoh, W.; Kimura, S.; Mori, S.; Shimada, K. Further characterization of sulfated homopolysaccharidess as anti-HIV agents. Experientia 1989, 45, 996–998. [Google Scholar] [CrossRef]
- Talyshinsky, M.M.; Souprun, Y.Y.; Huleihel, M.M. Anti-viral activity of red microalgal polysaccharides against retroviruses. Cancer Cell Int. 2002, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, S.V.; Ghosh, T. Abiotic stresses as tools for metabolites in microalgae. Bioresour. Technol. 2017, 244, 1216–1226. [Google Scholar] [CrossRef]
- Hu, H.; Wang, H.F.; Ma, L.L.; Shen, X.F.; Zeng, J.X. Effects of nitrogen and phosphorous stress on the formation of high value LC-PUFAs in Porphyridium cruentum. Appl. Microbiol. Biotechnol. 2018, 102, 5763–5773. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.Y. Effects of Different Salinities and Nitrogen and Phosphorus Concentrations on the Growth and Metabolism of Porphyridium. Master’s Thesis, Dalian University of Technology, Dalian, China, 2006. [Google Scholar]
- Xia, L.; Rong, J.F.; Yang, H.J.; He, Q.N.; Zhang, D.L. NaCl as an effective inducer for lipid accumulation in freshwater microalgae Desmodesmus abundans. Bioresour. Technol. 2014, 161, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Yuan-Kun, L.; Hai-Meng, T.; Ching-Seng, L. Effect of salinity of medium on cellular fatty acid composition of marine algae Porphyridium cruentum (Rhodophyceae). J. Appl. Phycol. 1989, 1, 19–23. [Google Scholar]
- Gilles, R.; Pequeux, A. Effect of salinity on the free amino acids pool of the red alga Porphyridium purpureum (= P. cruentum). Comp. Biochem. Physiol. Part A Physiol. 1977, 57, 183–185. [Google Scholar] [CrossRef]
- Hasanah, H.; Setyaningsih, I.; Uju, U. Harvesting and separation technique of Porphyridium cruentum polysaccharide using ultrafiltration membrane. Jurnal Pengolahan Hasil Perikanan Indonesia 2016, 19, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Freshwater Algae Culture Collection at the Institute of Hydrobiology Home Page. Available online: http://algae.ihb.ac.cn/english/mediumDetail.aspx?id=13 (accessed on 1 June 2013).
- Markou, G.; Muylaert, K. Effect of light intensity on the degree of ammonia toxicity on PSII activity of Arthrospira platensis and Chlorella vulgaris. Bioresour. Technol. 2016, 216, 453–461. [Google Scholar] [CrossRef]
- Lutzu, G.A.; Zhang, L.; Zhang, Z.; Liu, T.Z. Feasibility of attached cultivation for polysaccharidess production by Porphyridium cruentum. Bioprocess Biosyst. Eng. 2016, 40, 1–11. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ficner, R.; Lobeck, K.; Schmidt, G.; Huber, R. Isolation, crystallization, crystal structure analysis and refinement of B-phycoerythrin from the red alga Porphyridium sordidum at 2.2 Å resolution. J. Mol. Biol. 1992, 228, 935–950. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Liu, T.; Zhang, W.; Chen, X.L.; Wang, J.F. Biodiesel production from algae oil high in free fatty acids by two-step catalytic conversion. Bioresour. Technol. 2012, 111, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Golueke, C.G.; Oswald, W.J. The mass culture of Porphyridium cruentum. Appl. Microbiol. 1962, 10, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.M.; Oh, C.H.; Bae, H.J. Comparison of red microalgae (Porphyridium cruentum) culture conditions for bioethanol production. Bioresour. Technol. 2017, 233, 44–50. [Google Scholar] [CrossRef]
- Xu, X.M.; Ye, H.C.; Li, G.F. Research advances in plant salt tolerance mechanism. Chin. J. Appl. Environ. Biol. 2000, 6, 379–387. [Google Scholar]
- Ho, S.H.; Nakanishi, A.; Ye, X.T.; Chang, J.S.; Hara, K. Optimizing biodiesel production in marine Chlamydomona ssp. JSC4 through metabolic profiling and an innovative salinity-gradient strategy. Biotechnol. Biofuels 2015, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Huang, J.; Sandmann, G.; Chen, F. High-light and sodium chloride stress differentially regulate the biosynthesis of astaxanthin in Chlorella zofingiensis (Chlorophyceae). J. Phycol. 2009, 45, 635–641. [Google Scholar] [CrossRef]
- Redfield, A.C. The biological control of chemical factors in the environment. Sci. Prog. 1960, 11, 150–170. [Google Scholar]
- Razaghi, A.; Godhe, A.; Albers, E. Effects of nitrogen on growth and carbohydrate formation in Porphyridium cruentum. Cent. Eur. J. Biol. 2014, 9, 156–162. [Google Scholar]
- Adda, M.; Merchuk, J.C.; Arad, S. Effect of nitrate on growth and production of cell-wall polysaccharides by the unicellular red alga Porphyridium. Biomass 1986, 10, 131–140. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Maurya, R.; Trivedi, K.; Kumar-Patidar, S. Salinity induced oxidative stress enhanced biofuel production potential of microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 189, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Speer, H.; Kuyr, W. Studies on the growth of the red alga Porphyridium cruentum. Physiol. Plant. 1963, 16, 636–643. [Google Scholar] [CrossRef]
- Singh, S.; Arad, S.; Richmond, A. Extracellular polysaccharides production in outdoor mass cultures of Porphyridium sp. in flat plate glass reactors. J. Appl. Phycol. 2000, 12, 269–275. [Google Scholar] [CrossRef]
- Wang, J.; Chen, B.; Rao, X.; Huang, J.; Li, M. Optimization of culturing conditions of Porphyridium cruentum using uniform design. World J. Microbiol. Biotechnol. 2007, 23, 1345–1350. [Google Scholar] [CrossRef]
- Cohen, Z.; Vonshak, A.; Richmond, A. Effect of environmental conditions on fatty acid composition of the red alga Porphyridium cruentum: Correlation to growth rate. J. Phycol. 1988, 24, 328–332. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | Volume | Mother Liquor Concentration |
|---|---|---|
| KNO3 | 10 mL/L | 7.5 g/100ml dH2O |
| KH2PO4 | 1 mL/L | 2.5 g/100ml dH2O |
| MgSO4.7H2O | 10 mL/L | 2.0 g/100ml dH2O |
| Ferric citrate | 1 mL/L | 0.25 g/100ml dH2O |
| Soil extract | 10 mL/L | |
| dH2O | 484 mL | |
| Seawater | 484 mL |
| Components | Dosage |
|---|---|
| NaCl | 27.0 g |
| MgSO4.7H2O | 6.6 g |
| MgCl2.6H2O | 5.6 g |
| CaCl2.2H2O | 1.5 g |
| KNO3 | 1.0 g |
| KH2PO4 | 0.07 g |
| NaHCO3 | 0.04 g |
| 1M Tris-HCl | 20 mL |
| Trace element mother liquor | 1 mL |
| Fe-EDTA (Ethylene Diamine Tetraacetic Acid) | 1 mL |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Nan, F.; Feng, J.; Lv, J.; Liu, Q.; Liu, X.; Xie, S. Effects of Different Environmental Factors on the Growth and Bioactive Substance Accumulation of Porphyridium purpureum. Int. J. Environ. Res. Public Health 2020, 17, 2221. https://doi.org/10.3390/ijerph17072221
Lu X, Nan F, Feng J, Lv J, Liu Q, Liu X, Xie S. Effects of Different Environmental Factors on the Growth and Bioactive Substance Accumulation of Porphyridium purpureum. International Journal of Environmental Research and Public Health. 2020; 17(7):2221. https://doi.org/10.3390/ijerph17072221
Chicago/Turabian StyleLu, Xudan, Fangru Nan, Jia Feng, Junping Lv, Qi Liu, Xudong Liu, and Shulian Xie. 2020. "Effects of Different Environmental Factors on the Growth and Bioactive Substance Accumulation of Porphyridium purpureum" International Journal of Environmental Research and Public Health 17, no. 7: 2221. https://doi.org/10.3390/ijerph17072221