
Abstract
Synthetic fungicides are pesticides widely used in agriculture to control phytopathogenic fungi. The systemicity, persistency and intense application of some of these fungicides, such as boscalid, leads to long periods of exposure for honeybees via contaminated water, pollen and nectar. We exposed adult honeybees in the lab to food contaminated with boscalid for 33 days instead of the standard 10-day test. Most of the toxic effects were observed after 10 days. The median time to death (LT50) ranged from 24.9 days (lowest concentration) to 7.1 days (highest concentration) and was significantly shorter in all cases than with the control (32.0 days). The concentration and dietary doses of boscalid inducing 50% mortality (LC50 and LDD50, respectively) decreased strongly with the time of exposure: LC50 = 14,729 and 1,174 mg/l and LDD50 = 0.318 and 0.0301 mg bee−1 day−1 at days 8 and 25, respectively. We found evidence of reinforced toxicity when exposure is prolonged, but with an unusual pattern: no cumulative toxicity is observed until 17–18 days, when a point of inflexion appears that suggests a reduced capacity of bees to deal with the toxicant. Our results show the importance of time-to-death experiments rather than fixed-duration studies for evaluating chronic toxicity.
Similar content being viewed by others


Introduction
Fungicides and bactericides were the most widely used plant protection products in Europe in 2015 (41.76%), followed by herbicides (33.92%)1. Thus, it is not surprising to find fungicide residues in the environment and our food, for example in water2,3,4, air5, fruits6,7, vegetables6,7 or processed food8. Fungicide contamination has also been found in flower matrices such as nectar9,10, or pollen10,11. As a result, bees can be exposed to fungicides in flight, when collecting water, nectar or pollen or when consuming them afterwards.
One of the fungicides frequently detected in beekeeping matrices is boscalid, which is found in a range from a few μg/kg (ppb) up to 26.2 mg/kg (ppm), with pollen containing the largest amounts10,12,13,14,15,16,17,18,19,20,21,22,23,24. This molecule is authorised for a wide variety of uses. It is systemic (Kow = 2.96) and persistent in the soil (DT50 = 246 days)25. Simon-Delso et al.11,12 found that boscalid was the most frequent fungicide within beehives and in pollen pellets and contaminated samples of these pollen were found from July to October, ranging from 0.9 to 512 μg/kg. David et al.23 described the presence of boscalid residues both in oilseed rape pollen (up to 25 μg/kg) and pollen from field margins (up to 38 μg/kg), and proved that bees readily collect this pollen and bring the residues to the hive (up to 21 μg/kg). These results show that honeybees are frequently exposed to boscalid, including over extended periods.
A handful of studies have examined the eco-toxicity of this molecule. Boscalid inhibits mitochondrial respiration (succinate dehydrogenase) and blocks ATP production which affects cell respiration26. In mammals, it increases liver enzymes (alanine aminotransferase and gamma-glutamyl transferase), and is proved to induce thyroid adenomas although these reactions are considered adaptive and reversible3. Boscalid shows a low acute toxicity for honeybees, with an oral LD50 > 11 μg/bee14 and 100 μg/bee27 and a contact LD50 > 100 μg/bee27. To our knowledge, the biochemical effects on insects have not yet been described, but recently, boscalid (in co-formulation with pyraclostrobin) was found to be linked to decreased pollen consumption and digestion in bees, as well as lower ATP concentrations in the thoracic muscle tissue and higher virus titres28. A recent field study also revealed a significant positive correlation between honeybee colony disorders over the winter period and the presence of fungicides residues (mainly boscalid) in bee hives in the summer and autumn12. In addition, boscalid has been shown to interact with neonicotinoid insecticides such as thiamethoxam and clothianidin, whose toxicity to honeybees nearly double24. Disruptive effects on nest recognition of solitary bees Osmia lignaria and Megachile rotundata have been described following exposure to field doses of fungicides, including boscalid29. Around 60% of bumble bee queens died when exposed to blueberry plant material treated with Pristine® (pyraclostrobin 12.8% and boscalid 25.2%) at a rate of 1.6 kg/ha30. As is the case with chronic toxicity, hardly any of the described interactions of boscalid with other chemicals were considered when boscalid was first authorised.
A more careful examination of the long-term toxicity of boscalid for honeybees is appropriate based on the combination of these first toxicity results and the duration of the exposure of bees to boscalid. A re-examination may also be relevant for other molecules because the chronic toxicity of most pesticides entering the European market before 2014 was not evaluated within the pesticide authorisation procedure (Regulation EU 283/2013)31.
In a previous study we found only limited evidence for long-term toxicity of pure boscalid for honeybee larvae32, but its impact on adults remains to be investigated. Standard guidelines recommend a ten-day trial for chronic toxicity evaluation on adults33. Preliminary results (unpublished data) have shown that the toxicity of this product may appear only after periods longer than ten days. Repeated or prolonged exposure may also induce “time-reinforced toxicity” also called “cumulative toxicity” due to bioaccumulation of the product in the body or to other mechanisms34. Several different methodologies are described in the literature for detecting such cumulative toxicity35,36,37.
The main objectives of this study are (1) to evaluate the chronic toxicity of a commercial formulation of boscalid with a time-to-death approach: the study finishes only when the mortality rate in the control reaches 50%; (2) to check for evidences of cumulative toxicity using different methodological approaches. In addition, we carefully examined the kinetics of food consumption not only because this parameter is crucial to evaluating the real exposure, but also because it can provide insights about the physiological changes over the life of the bees.
Results
Food consumption and duration of the trial
The average daily food consumption per bee over the duration of the trial (33 days) was 39 ± 1.7 mg bee−1 day−1, and no significant difference was found between the different treatments (Gaussian Mixed model with cage as random effect: F5,12.2 = 2.289, p = 0.1104). However, syrup consumption was not constant over time and showed clear differences between treatments once time was taken into account. Firstly, we observe a strong increase of individual food consumption in most cages when there were only 1 to 3 living bees left, with extreme values above 0.08 g syrup bee−1 day−1 (see Fig. 1 and Supplementary Material (Suppl. Mat.), Section 3.2.1). These extreme values may be partially due to the measurement error induced by syrup evaporation. The difference in syrup weight due to evaporation is divided by the number of living bees. Consequently, the relative importance of evaporation increases when there are only a few living bees left. Nevertheless, after correcting consumption for evaporation (Suppl. Mat., Section 3.2.2) we still obtained some large peaks of consumption when there were only a few living bees left.

Daily syrup consumption at different boscalid concentrations. The black dots represent the mean value and the bars are the standard deviation. Each transparent coloured point represents the consumption per bee during one day in one cage. There is no significant difference of average consumption between treatments (Gaussian mixed model with cage as random effect: F5,12.2 = 2.289, p = 0.1104).
Secondly, when we removed these outliers by only considering consumption data while a minimum of 5 bees remained per cage, we observed that consumption was still not stable over the duration of the test (Fig. 2). An inverted “bell” shape was observed in the control, with individual consumption increasing to a maximum around days 17–18 of the test and then decreasing slowly until the end of the observation period (33 days). A similar pattern in consumption was observed for the bees at the different concentrations, but with differing kinetics: at higher concentrations, the maximum was reached sooner and the increase and decrease were steeper (Fig. 2). At a given point in time, bees tend to significantly reduce their consumption when Cantus® contaminates the food, the reduction being greater at higher concentrations (These visual differences were supported by statistical testing with a polynomial mixed model, see Suppl. Mat., Section 3.2.3).

Daily syrup consumption over time until 50% mortality is reached in each cage. The lines for each boscalid concentration are loess trends (locally weighted polynomial regression). See Suppl. Mat., Section 3.2.3 for detailed representation of observed consumption rates and statistical testing with a polynomial mixed model. The model show highly significant time x concentration and main concentration effects: the kinetics of consumption was different between treatments and the average consumption on a given day was also different.
Mortality rate and toxicological endpoints
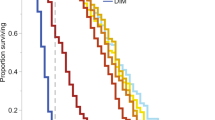
The mortality rates were in agreement with the validity criteria established by OECD draft test guidelines for the evaluation of the toxicity of pesticides over ten-days33: (1) the average mortality for the controls was lower than 15% at day 10 of exposure and remained so up to day 20; (2) the average mortality in the toxic standard was higher than 50% at day 10 (Fig. 3).

Observed mortality rate over the duration of the test. N = 30 bees per treatment, except for boscalid 9,000 mg/l, n = 29. Dashed vertical lines indicate ten days, i.e. duration of the test currently proposed by international standards, and 20 days, i.e. time at which the mortality in the control reached 15%. Current international standards (ten days) propose as validity criteria up to 15% mortality in the control group. Toxic standard was dimethoate at 1.5 mg/l.
Before day 10 - i.e. the normal duration of a chronic test - observed mortality was low. A mortality rate higher than 50% was only observed at the highest Cantus® concentration (18,000 mg boscalid/l) from day 8 onwards. On day 10, mortality had reached almost 100% at the highest concentration, but remained below 50% for all other treatments and close to 0% in most cases. On day 20, when mortality in the control had reached 15%, mortality for the two highest doses (9,000 and 4,500 mg boscalid/l) was 100%. On day 31, the mortality rate of all Cantus® treatments (including 2,250 and 1,125 mg boscalid/l) had reached 100% while that for the control was just below 50%.
The median lethal time (LT50) decreased as the concentration increased (Fig. 4a): it was 7 days (95% CI: 6.89, 7.37) at the highest concentration (18,000 mg/l) and 25 days (24.38, 25.47) at the lowest tested concentration (1,125 mg/l, Fig. 4a). The LT50 of the latter group was significantly lower than the 32 days (29.84, 34.16) of the control group. The LT50 of the toxic standard was 4 days (3.87, 4.4).

Representations of LTx vs. Concentration (a), LCx vs. Time (b), LDDx vs. Time (c) and LCDx vs. Time (d). All graphs represent the toxic endpoint for 10%, 50% and 90% mortality estimated with three parameters Weibull 2 models. See Table 1 for meaning of the abbreviations. See Suppl. Mat., “raw_results” directory to obtain the raw numbers used in these graphs.
Figure 4b–d, summarise the LC50, LDD50 and LCD50 values from day 8 to day 25 of the trial (see values in Supplementary Material and data files in the “raw_results” directory). LCx and LDDx decreased over time. LC50 started at 14,728.73 mg/l (12,055.50, 17,401.95) on day 8 and was 1,174.25 mg/l (−150.51, 2,499.01) on day 25. In terms of individual dose, LDD50 was 0.32 mg boscalid bee−1 day−1 (0.27, 0.36) on day 8, and decreased to 0.03 mg boscalid bee−1 day−1 (−0,004, 0.064) on day 25.
The median lethal cumulative dose (LCD50) showed a plateau around 2.4 mg boscalid bee−1 until day 17, when LCD50 started to decrease, reaching 0.76 mg boscalid bee−1 (0.42, 1.480) on day 25 (Fig. 4d).
Cumulative toxicity - first approach: log-log linear regressions
The slopes of all log-log linear regressions were significantly lower than −1 (Suppl. Mat., Fig. 20) except for the slope of the regression between log (concentration) and log (LT90), which was only at the margin of statistical significance (CI: −3.370, −0.995). The time-concentration relationships diverged clearly from Haber’s law and pointed to an increased toxicity for long time exposure.
However the slope is only a rough summary of this relationship. Plotting the data revealed that the log-log relationship was not always linear and showed interesting patterns (see for example Fig. 5 and Suppl. Mat., Section 5.1 for more details).

Log-log regression of LDDx vs. Time compared to the regression line expected in the absence of cumulative toxicity. The dashed lines show regression lines with a fixed −1 slope (the models estimate only the intercepts). This is the expected relationship under Haber’s law i.e. without cumulative toxicity. The slopes of the observed linear regression are significantly lower than −1, which implies some level of cumulative toxicity. However the log-log relationship is clearly not linear at least for the lowest mortality rates. The representation of the log-log regressions for the other toxic endpoints can be found in Suppl. Mat., Section 5.1. along with confidence intervals of the slopes (Fig. S20).
More precisely, the log (LDDx) vs log (Time) relationship seemed to follow Haber’s rule closely up to day 17–18 with a slope around −1. At this point the slope abruptly decreased and the relationship clearly deviated from Haber’s rule (Fig. 5). This pattern was more obvious for lower levels of mortality (LDD10, LDD20, etc) and tended to disappear at higher levels of mortality (LDD90). For the LDD90, the points were almost perfectly aligned on a straight line without any inflection point. However, even for the LDD90, where the observed regression line seemed to be close to the theoretical Haber’s line, the slope was significantly smaller than the expected −1 (estimate = −1.359 with a 95% confidence interval of (−1.492, 1.226)).
This pattern was less marked on the log (LCx) vs log (Time) graph (Suppl. Mat., Figs 24 and 25) and not visible on the log (Concentration) vs log (LTx) graphs (Suppl. Mat., Figs 22 and 23); however, the later graphs were based on five points only, which was probably not enough to visualise this non-linear relationship.
Cumulative toxicity - second approach: cumulative doses
The cumulative dose necessary to reach 50% mortality was similar (approximatively 2.5 mg a.i./bee) for the three highest-concentration treatments (18,000; 9,000 and 4,500 mg a.i./l syrup – see Suppl. Mat., Fig. 6). The same level of mortality was reached with significantly lower cumulative doses for the two lowest-concentration treatments: 1.38 mg a.i./bee for a concentration of 2,250 mg a.i./l syrup and 0.84 mg a.i./bee for a concentration of 1,125 mg a.i./l syrup (Fig. 6 and Suppl. Mat., Section 5.2). While the time to reach 50% mortality (Fig. 6: black dots - LT50) was longer for the lowest concentrations, the cumulative dose necessary was much lower than what we observed for higher-concentration treatments.

Cumulative dose consumed per bee for each treatment (concentration) over time. The black dots indicate the estimated values of LT50 (with 95% confidence intervals) for each concentration. Without cumulative toxicity, the black dots should appear horizontally at approximately the same level of cumulative dose or aligned vertically above the control LT50 for the two lowest concentrations.
Without cumulative toxicity (under Haber’s rule) the bees should be exposed to the same cumulative dose to reach the same level of mortality. Again, these results point to an increased toxicity in the context of a long-term exposure to Cantus®.
Discussion
Syrup consumption
Despite having used a standardised methodology, syrup consumption was highly variable within the cages, between the cages and over time. We observed a non-constant individual daily syrup consumption over the duration of the trial, with high consumption values when only a few bees were left in the cage (1 to 3 bees). These extreme values may be in part due to syrup evaporation, which is why it would be worthwhile to include this parameter as a measurement to be performed during chronic toxicity testing. In contrast to our results, Arnold38 observed lower levels of consumption when bees were isolated. We also clearly observed a non-constant consumption level over time and a difference between the treatments both in terms of kinetics (the first peak of consumption appeared at a different time) and in term of absolute consumption (the food intake was lower when the concentration was higher whatever the time). In contrast to our observations, a constant daily consumption per bee and no treatment effect were reported in a similar experiment testing chronic mortality rates of imidacloprid and deltamethrin39. However, our results show that real consumption differences may be masked when the data are analysed with simple statistical approaches because of complex non-linear consumption vs time relationships and differences in kinetics (time x treatment interactions). In addition, the difference in food consumption found between the different doses and over time, mainly at the beginning of the test, could be linked to the perception/taste of the substance by the bees as is the case with other products40,41,42. To our knowledge, no repellent effect has been described for boscalid, but additional trials could be performed to evaluate the preferences of bees between diets containing boscalid or without it.
The variation of syrup consumption over the trial and the food distribution among bees are realities whose impact needs to be considered during the interpretation of toxicity results, as they will have an impact on dose calculation, but also on the implementation of methodologies proposed for regulatory purposes, i.e. cumulative toxicity36.
Toxicity of Cantus® for bees
The mortality rate of the bees in the Cantus® treatments was significantly higher than that of the control group. The chronic toxicity of this pesticide in terms of concentration (LC50) was 1,174 mg boscalid/l on day 25. This concentration is far from the highest residue value described in the literature for boscalid in pollen (26 mg/kg)16, but lies in the lower range of concentrations applied in the field. In addition, the effects measured here are mortality rates; we could expect sublethal effects at even lower concentrations. Both LC50 and LDD50 decreased in time, their values being reduced by approximately 90% from day 8 to day 25. Similar results have been found in other studies, mainly with insecticides39,43,44,45,46,47,48,49,50,51,52. The main differences between our study and others were the level of toxicity described and the order of magnitude of the concentrations tested. These tested concentrations are a reflection of the amount of residues bees are exposed to in natural conditions.
Standard methodologies currently prescribe the testing of chronic toxicity over a period of ten days33. These standard methodologies are themselves inspired by previous toxicological studies evaluating chronic effects on bees after 10–14 days exposure44,50,51. This duration seems adequate for testing pesticides with high to moderate acute toxicity for bees. The effects we observed here, however, would not have been detected over a duration of ten days. Pesticide exposure longer than ten days is not an unrealistic situation for bees, given that boscalid has been found to contaminate food resources over months11. This is not surprising considering that boscalid is systemic, persistent and authorised for a large list of uses, even during the flowering period of many crops53. For this reason, we think that a time-to-death approach might be more appropriate for evaluating the risk of chronic exposure to a pesticide. The estimation of the LTx as toxicological endpoint is useful for comparing the effect of chronic exposure to different doses and makes in possible to evaluate the impact of stressors on the lifespan of bees. In our case, even the lowest concentration tested significantly reduced the time to reach 50% mortality. An additional advantage of such an approach is that it can help detect a potential cumulative toxicity.
Cumulative toxicity
A first clear indication of the existence of cumulative toxicity of Cantus® can be inferred from the relationship between the Lethal Cumulative Dose (LCDx) and Time. The plateau observed in Fig. 4d until day 17–18 indicates that a certain number of bees managed to detoxify the ingested pesticide: whatever the duration of repeated exposure, the same cumulative amount of pesticide was necessary to kill 50% of the bees. From days 17–18 onwards, a decreasing total amount of pesticide was sufficient to kill the same number of bees. Their detoxifying capacity seemed to be exceeded, or the accumulation of boscalid in the bees’ bodies was enough to trigger toxic effects which increased with time. This time-increased-toxicity and sudden change in toxicity profile around 17–18 days were confirmed by several other statistical approaches: (1) the log-log regressions testing whether the relationships between concentration or dose and time followed Haber’s rule54 and (2) the so-called EFSA protocol (comparison of cumulative doses to reach 50% of corrected mortality between concentrations)36.
Moncharmont et al.39 observed a delayed toxicity of the insecticide imidacloprid in bees and argued that the effects could be due to age-dependent sensitivity or to the accumulation of the compound in the body. The dynamic of syrup consumption in the control also suggested a naturally occurring change in the physiology of the bees even without consumption of Cantus®: at days 18–19 of their life, bees in the control achieved a maximum syrup consumption, while the mortality rate was slowly increasing. The sudden change of toxicity around 17–18 days could then be explained by higher sensitivity due to the natural ageing process, by physiological fatigue due to repeated exposure to the pesticide during their life, or by a combination of the two processes. The similar patterns of food consumption observed for the different concentrations, but with maximums at earlier dates for higher concentrations, might indicate faster physiological ageing for bees exposed to the pesticide and suggest that ageing alone is not a sufficient hypothesis to explain the patterns observed. The effects of xenobiotics on bees involve responses of detoxification and oxidative and general stress leading to an increase in the insect’s energetic metabolism55. These may lead to physiological wear-out and reduction of vitellogenin production, thus shortening the lifespan and immuno-competence of bees17,56,57,58,59. We did not supply any dietary protein during the test. The already limited detoxification capacity of bees60 may therefore have been even more restricted when the bees exhausted their capacity for protein synthesis due to a lack of essential aminoacids61.
The toxicity and the sudden change in toxicity could also be explained by mechanisms other than ageing and physiological fatigue. For example, as boscalid is moderately lipophilic (Kow = 2.96)25, it could have a certain tendency to accumulate in the bees’ fatty tissue (i.e. bioconcentration). The products contained in Cantus® or their metabolites could accumulate in the bees’ fatty tissue from the onset of exposure up to a level of saturation, when these chemicals could target other tissues, accelerating the toxic effect of these compounds if exposure is prolonged. Unfortunately we did not evaluate the residues of boscalid and metabolites in dead bees. This would have provided very useful information about contamination in the bees and the accumulation process in their bodies according to the concentrations used47. The lower food intake when the syrup was contaminated by the pesticide may also have contributed to the weakening of the bees and decreased their capacity to handle the toxicity in the long term. Finally, it must be stressed that the toxicity observed here may have been induced by boscalid itself but also by the co-formulants present in Cantus® or interactions between all these xenobiotics62. Limited information is available on co-formulates because few ingredients are revealed in the description of the product. Phyto-pharmaceutical companies regard the composition of their formulation as trade-secret.
Cumulative toxicity - methodological discussion
The time-to-death approach used here generated a large amount of data that allowed us to explore different methodologies for testing for cumulative toxicity. The main advantage of the protocol proposed by EFSA is that it is less costly: after an acute toxicity test of 48 h, one is required to perform a new test and monitor mortality for only two concentrations (the LC50 and ¼ of this concentration) up to 50% mortality and then test whether the cumulative lethal doses are equal. However, in our case, the toxicity after 48 h was too low to estimate the LC50 (no acute toxicity). After ten days, the estimated LC50 was approximately 10,000 mg/l and could have been compared with a 2,500 mg/l treatment. Instead of running a new test, we monitored mortality for all our initial concentrations until the control reached 50% mortality. Using these data, we were able to show that the cumulative doses to kill 50% of the bees at a concentration of 9,000 mg/l (the concentration closest to the LC50) and 2,250 mg/l are significantly different, supporting the hypothesis of cumulative toxicity with an approach close to the simple EFSA protocol. However, using only two concentrations would have masked some of the most interesting patterns observed in this study, which could help us to understand the toxicological effect of the pesticide. The log-log regression between Lethal Dose and Time revealed a clear breaking point in the toxicological effect after 17–18 days which would have gone unnoticed with simpler approaches. The log-log regression between Concentration and Lethal Time was also less interesting for the same reason: the lower number of points (i.e. the number of different concentrations tested) makes such subtle patterns difficult to spot. We also showed that simply estimating the slope of the log-log regression, while useful, was only a rough summary: it was necessary to plot the data to check if we had a simple, constant, deviation from Haber’s rule or a more complex relationship between toxicity and time. The Lethal Dose should also be preferred to the Lethal Concentration in log-log regressions, because the latter does not take into account the differences in food intake between treatments. However, Lethal Concentration estimates are still useful for comparing the lab results with the concentrations observed in beehive matrices or applied in the field by farmers.
Conclusion
In conclusion, we showed that at field application rates, Cantus® (500 g boscalid/kg) leads to chronic toxicity in honeybees that would have remained undetected with current proposed methodologies for pesticide risk assessment. This fungicide significantly reduced the lethal time for all concentrations tested. Furthermore, a cumulative toxicity potential was detected particularly after 17–18 days of exposure. All in all, we recommend a time-to-death approach rather than fixed-duration studies for exploring the chronic toxicity effects of pollutants that are present over long periods in field conditions.
Materials and Methods
Test substances
We tested five concentrations of boscalid: 18,000, 9,000, 4,500, 2,250 and 1,125 mg a.i./l. The test substances were dissolved in a 50% w/v sucrose solution provided as food to the bees ad libitum. We used the commercial formulation Cantus® (BASF, 500 g/kg solid boscalid) because the pure active substance has a low water solubility (4.6 mg/l). We had tried to perform earlier experiments with the pure active ingredient, but could not be sure of the exposure due to its low solubility and its flocculation. The concentrations were chosen based on previous studies and the potential concentrations used in the field. Application rates in the field range from 250 to 1,880 g a.i./ha27. Based on the maximum application rate and on the typical spray rate per hectare in Belgium (100–200 l/ha), a concentration of 18,800–9,400 mg/l would be in the upper range of concentration expected to be found in the field, while 1,125 mg/l would be in the lower range. Dimethoate (Perfektion®, BASF, 400 g/l dimethoate) was used as toxic standard at 1.5 mg a.i./l as recommended by OECD standards33 to ensure that bees were well exposed to the toxicants.
Bees
Three frames containing capped cells with emerging bees were collected from three healthy queen-right colonies with queens of different origin. They were left in an incubator at 32.8 °C ± 1 °C and 60% ± 20% relative humidity. One-day-old worker bees were transferred without anaesthesia into cages in groups of ten bees per cage.
Experimental conditions
The bees were kept in cardboard cages with a mobile plastic window, in darkness, within an incubator at 32.8 °C ± 1 °C and 60% ± 20% relative humidity for the duration of the test. They were fed ad libitum with a syrup (sucrose solution with different concentrations of test substances) provided through 2 ml plastic syringes with the tip removed.
Treatments and data gathering
At the beginning of the test, each treatment group was composed of three replicates each containing ten bees. In one cage one of the ten bees died before the start of the experiment (i.e. before the contaminated syrup was provided). The initial number of bees was considered to be nine, instead of ten, in this cage. Bee mortality was recorded every day at approximately the same time as the syringes were changed. Dead bees were removed at each assessment. Syringes containing freshly prepared syrup were replaced every day and weighed just before administration and again the following day to estimate the quantity of syrup consumed by the bees. This consumption value was divided by the number of living bees to obtain the consumption per bee and per day. The test lasted until mortality in the control group reached 50%, instead of stopping the test after ten days as recommended by OECD guidelines33.
Data analysis
All analyses were performed with R63 mainly with drc package64 for dose response curves (Weibull models) and toxicological endpoints estimations, the lme4 package65 for mixed models and the multcomp package66 for post-hoc multiple comparisons. All datasets, R code and detailed data analyses are provided as a public figshare repository (https://figshare.com/s/865e87feaad34c095bbd).
Data analysis – Syrup consumption
We first compared the daily syrup consumption per bee between the different treatments (different Cantus® concentrations) without taking the time into account. We used a Gaussian mixed model with the cage as random effect and the treatment as fixed effect. We then analysed how syrup consumption changed over time for each treatment. We used graphical tools such as loess (locally weighted polynomial regression) to visualise the trends in the noisy data and used mixed models to check if the observed patterns were statistically significant. We used the cage as random effect and the treatment, time (second-order polynomial) and their interactions as fixed effects. The evaporation of syrup was also measured for a subset of the days in cages without bees or with only dead bees (see Suppl. Mat., Section 3.1).
Data analysis – Toxicological endpoints
We computed four toxicological endpoints (and their 95% confidence intervals): Lethal Time (LTx), Lethal Concentration (LCx), Lethal Dietary Dose (LDDx) and Lethal Cumulative Dose (LCDx) (Table 1). The LTx was computed for each concentration and the LCx, LDDx and LCDx were computed for each day of the test between day 8 and day 25 (outside this range, the mortality was either too low or too high). All “x” levels of mortality between 10% and 90% in 10% increments were calculated.
To estimate these toxicological endpoints, we chose a three-parameters Weibull 2 model (sensu Ritz)67 to model the relationship between uncorrected mortality and time, concentration or dose. This three-parameters-sigmoid model fixed the higher asymptote (i.e. the mortality is 100% for an infinite dose) and estimated the lower asymptote (i.e. the mortality in the control). The toxicological endpoints were then calculated relative to these two asymptotes. The LCx, LDDx and LCDx estimates were therefore corrected for mortality in the control. For LTx we used the same approach but the lower asymptote in this case estimated the mortality at D0, which was zero. Hence, the LTx values as we calculated them here were not corrected for mortality in the control. See Suppl. Mat., Section 7, for more details.
Data analyses – Cumulative toxicity estimation
Most of the methods used to evaluate the potential of cumulative toxicity (or time reinforced toxicity) were based on Haber’s rule54, which states that the product of exposure concentration (C) and exposure duration (t) leads to a constant toxic effect. In other words, if there is no cumulative toxicity, when the concentration (or dose) is divided by two the time of exposure to reach the same level of mortality should be doubled. Haber’s rule is a specific case of the Druckrey-Küpfmüller model, for which the exponent (b) has an absolute value of 134.
We used two different approaches to test if Cantus® induced time-reinforced toxicity: (1) log-log linear regression between concentration or dose and time to estimate the slope b from Eq. 1 (Eq. 1)35; (2) comparison between treatments of the cumulative dose needed to reach 50% mortality as proposed by the European Food Safety Authority (EFSA)36.
For the first approach, we estimated the slope (and 95% Confidence Intervals) of three types of simple linear regressions: (1) log (Concentration) vs log (LTx); (2) log (LCx) vs log (Time); (3) log (LDx) vs log (Time). Should cumulative toxicity exist, these slopes would be significantly smaller than −1. This was repeated for each level of mortality between 10% and 90% in 10% increments.
For the second approach, the EFSA36 has suggested evaluating the LC50 at 48 h and then running a test with two treatments: one corresponding to the estimated LC50 value and another with ¼ of this concentration. The cumulative dose needed to reach 50% mortality in each treatment is then compared. These cumulative doses should be equal if there is no cumulative toxicity.
We did not observe mortality in any treatment at 48 h, but we used a similar idea by comparing the cumulative dose needed to reach 50% mortality in our five concentrations (with two-fold dilutions). We used a one-way ANOVA (each cage in a dose was a replicate) and all pair-wise post-hoc comparisons (multcomp package66 to test for differences. For this part only we used corrected mortalities based on Abbott’s formula68.
A third approach proposed by Miller35 provided unconvincing results which are not detailed here (See Section 5.3 in the Suppl. Mat. for the details).
Data availability
All datasets, R code and detailed data analyses are provided as a public figshare repository (https://figshare.com/s/865e87feaad34c095bbd) and the Supplementary Material contain enough detail to understand the analyses performed.
References
Eurostat. Data on pesticide sales. Available at: http://ec.europa.eu/eurostat/data/database. Accessed: 25th September 2017 (2017).
Kreuger, J., Graaf, S., Patring, J. & Adielsson, S. Pesticides in surface water in areas with open ground and greenhouse horticultural crops in Sweden 2008 (2010).
Elskus, A. A. Toxicity, Sublethal Effects, and Potential Modes of Action of Select Fungicides on Freshwater Fish and Invertebrates: U.S. Geological Survey. 42 p (2012).
Samson-Robert, O., Labrie, G., Chagnon, M. & Fournier, V. Neonicotinoid-Contaminated Puddles of Water Represent a Risk of Intoxication for Honey Bee. Plos One 9, e108443 (2014).
Woodrow, J. E., Seiber, J. N. & Fitzell, D. Analytical method for the dithiocarbamate fungicides ziram and mancozeb in air: preliminary field results. J. Agric. Food Chem. 43, 1524–1529 (1995).
Caldas, E. D., Conceiçao, M. H., Miranda, M. C. C., de Souza, L. C. K. & Lima, J. F. Determination of dithiocarbamate fungicide residues in food by a spectrophotometric method using a vertical disulfide reaction system. J. Agric. Food Chem. 49, 4521–4525 (2001).
EFSA. The 2014 European Union Report On Pesticide Residues In Food. EFSA J. 14, 139 pp (2016).
Cabras, P. et al. Fate of Some New Fungicides (Cyprodinil, Fludioxonil, Pyrimethanil, and Tebuconazole) from Vine to Wine. J. Agric. Food Chem. 45, 2708–2710 (1997).
Kubik, M. et al. Pesticide residues in bee products collected from cherry trees protected during blooming period with contact and systemic fungicides. Apidologie 30, 521–532 (1999).
Johnson, R. M., Ellis, M. D., Mullin, C. A. & Frazier, M. Pesticides and toxicity – USA. Apidologie 41, 312–331 (2010).
Simon-Delso, N., San Martin, G., Bruneau, E., Delcourt, C. & Hautier, L. The challenges of predicting pesticide exposure of honey bees at landscape level. Sci. Rep. 7, 3801 (2017).
Simon-Delso, N. et al. Honeybee Colony Disorder in Crop Areas: The Role of Pesticides and Viruses. Plos One 9, e103073 (2014).
Illies, I., Büchler, R. & Wallner, K. Strategies to reduce the input of pesticides in colonies from the fungicide Cantus. Apidologie 38, 482–504 (2007).
Aubee, C. & Lieu, D. Environmental Fate and Ecological Risk Assessment for Boscalid New Use on Rapeseed, Including Canola (Seed Treatment). 1–52 (U.S. Environmental Protection Agency, 2010).
Mullin, C. A. et al. High Levels of Miticides and Agrochemicals in North American Apiaries: Implications for Health. Plos One 5, e9754 (2010).
Wallner, K. Sprayed and seed dressed pesticides in pollen, nectar and honey of oilseed rape. Julius-Kühn-Archiv 423, 148 pp (2010).
Wu, J. Y., Anelli, C. M. & Sheppard, W. S. Sub-lethal effects of pesticide residues in brood comb on worker Honey Bee (Apis mellifera) development and longevity. Plos One 6, e14720 (2011).
Pohorecka, K. et al. Residues of neonicotinoid insecticides in bee collected plant materials from oilseed rape crops and their effect on bee colonies. J. Apic. Sci. 56, 115–134 (2012).
Stoner, K. A. & Eitzer, B. D. Using a hazard quotient to evaluate pesticide residues detected in pollen trapped from honey bees (Apis mellifera) in Connecticut. Plos One 8, e77550 (2013).
Greenpeace. The bees’ burden. An analysis of pesticide residues in comb pollen (beebread) and trapped pollen from honey bees (Apis mellifera) (2014).
Jabot, C. et al. Trace-level determination of pyrethroid, neonicotinoid and carboxamide pesticides in beeswax using dispersive solid-phase extraction followed by ultra-high-performance liquid chromatography-tandem mass spectrometry. Int. J. Environ. Anal. Chem. 95, 240–257 (2015).
Ravoet, J., Reybroeck, W. & de Graaf, D. C. Pesticides for Apicultural and/or Agricultural Application Found in Belgian Wax Combs. Bull. Environ. Contam. Toxicol. 94, 543–548 (2015).
David, A. et al. Widespread contamination of wildflower and bee-collected pollen with complex mixtures of neonicotinoids and fungicides commonly applied to crops. Environ. Int. 88, 169–178 (2016).
Tsvetkov, N. et al. Chronic exposure to neonicotinoids reduces honey bee health near corn crops. Science 356, 1395–1397 (2017).
IUPAC. Pesticide Properties Database (PPDB) - Boscalid (Ref: BAS 510F) (2014). Available at: http://sitem.herts.ac.uk/aeru/ppdb/en/86.htm. (Accessed: 3rd January 2016).
Stammler, G. et al. Studies on the biological performance of boscalid and its mode of action. In Modern fungicides and antifungal compounds V: 15th International Reinhardsbrunn Symposium, Friedrichroda, Germany, May 6–10, 2007 45–51 (Deutsche Phytomedizinische Gesellschaft e.V. Verlag, 2008).
European Commission. Review report for the active substance boscalid. Finalised in the Standing Committee on the Food Chain and Animal Health at its meeting on 22 January 2008 in view of the inclusion of boscalid in Annex I of Council Directive 91/414/EEC. SANCO/3919/2007-rev. 5, 21 January 2008 (2008).
Degrandi-Hoffman, G., Chen, Y., Dejong, E. W., Chambers, M. L. & Hidalgo, G. Effects of Oral Exposure to Fungicides on Honey Bee Nutrition and Virus Levels. J. Econ. Entomol. 1–11 (2015).
Artz, D. R. & Pitts-Singer, T. L. Effects of Fungicide and Adjuvant Sprays on Nesting Behavior in Two Managed Solitary Bees, Osmia lignaria and Megachile rotundata. Plos One 10 (2015).
Skyrm, K. M. & Rao, S. Impact of Pesticide Residues on a Native Bumble Bee Pollinator, Bombus Vosnesenskii (Hymenoptera: Apidae). Proceedings of the 69th Annual Pacific Northwest Insect Management Conference, held January 11th and 12th, 2010 at the Hilton Hotel, Portland, Oregon (2010).
European Commission. CommissionRegulation (EU) No. 283/2013 of 1 March 2013 setting out the data requirements for active substances, in accordance withRegulation (EC) No. 1107/2009 of the European Parliament and of the Council concerning the placing of plant protection products on the market. OJ L 93, 3.4.2013, pp. 1–84 (2013).
Simon-Delso, N., San Martin, G., Bruneau, E., Hautier, L. & Medrzycki, P. Toxicity assessment on honey bee larvae of a repeated exposition of a systemic fungicide, boscalid. Bull. Insectology 70, 83–89 (2017).
OECD. Honey bee (Apis mellifera L.), chronic oral toxicity test (10 day feeding test in the laboratory). Revised by the International OECD Ring Test Group on 08 December 2014 (2014).
Tennekes, H. A. & Sánchez-Bayo, F. Time-Dependent Toxicity of Neonicotinoids and Other Toxicants: Implications for a New Approach to Risk Assessment. J. Environment. Analytic. Toxicol. S4, 001, https://doi.org/10.4172/2161-0525.S4-001 (2011).
Miller, F. J., Schlosser, P. M. & Janszen, D. B. Haber’s rule: a special case in a family of curves relating concentration and duration of exposure to a fixed level of response for a given endpoint. Toxicology 149, 21–34 (2000).
EFSA. Guidance on the risk assessment of plant protection products on bees (Apis mellifera, Bombus spp. and solitary bees). EFSA J. 11, 3295 [266 pp.] (2013).
Sánchez-Bayo, F. From simple toxicological models to prediction of toxic effects in time. Ecotoxicology 18, 343–354 (2009).
Arnold, G. Influence de l’isolement sur la consommation alimentaire des ouvrières d’abeille. Apidologie 10, 29–34 (1979).
Moncharmont, F.-X. D., Decourtye, A., Hennequet-Hantier, C., Pons, O. & Pham-Delègue, M.-H. Statistical analysis of honeybee survival after chronic exposure to insecticides. Environ. Toxicol. Chem. 22, 3088–3094 (2003).
Kessler, S. C. et al. Bees prefer foods containing neonicotinoid pesticides. Nature 521, 74–76 (2015).
Tison, L. et al. Honey bees’ behavior is impaired by chronic exposure to the neonicotinoid thiacloprid in the field. Environ. Sci. Technol. 50, 7218–7227 (2016).
Wright, G. A. To feed or not to feed: circuits involved in the control of feeding in insects. Curr. Opin. Neurobiol. 41, 87–91 (2016).
Pham-Delègue, M.-H. et al. Long-term effects of soybean protease inhibitors on digestive enzymes, survival and learning abilities of honeybees. Entomol. Exp. Appl. 95, 21–29 (2000).
Suchail, S., Guez, D. & Belzunces, L. P. Discrepancy between acute and chronic toxicity induced by imidacloprid and its metabolites in Apis mellifera. Environ. Toxicol. Chem. SETAC 20, 2482–6 (2001).
Schmuck, R., Schöning, R., Stork, A. & Schramel, O. Risk posed to honeybees (Apis mellifera L., Hymenoptera) by an imidacloprid seed dressing of sunflowers. Pest Manag. Sci. 57, 225–238 (2001).
Mommaerts, V. et al. Risk assessment for side-effects of neonicotinoids against bumblebees with and without impairing foraging behavior. Ecotoxicology 19, 207 (2010).
Sánchez-Bayo, F., Belzunces, L. & Bonmatin, J.-M. Lethal and sublethal effects, and incomplete clearance of ingested imidacloprid in honey bees (Apis mellifera). Ecotoxicology 26(9), 1199–1206 (2017).
Rondeau, G. et al. Delayed and time-cumulative toxicity of imidacloprid in bees, ants and termites. Sci. Rep. 4, 5566 (2014).
Ramirez-Romero, R., Chaufaux, J. & Pham-Delègue, M.-H. Effects of Cry1Ab protoxin, deltamethrin and imidacloprid on the foraging activity and the learning performances of the honeybee Apis mellifera, a comparative approach. Apidologie 36, 601–611 (2005).
Decourtye, A. et al. Comparative sublethal toxicity of nine pesticides on olfactory learning performances of the honeybee Apis mellifera. Arch. Environ. Contam. Toxicol. 48, 242–250 (2005).
Illarionov, A. I. Toxic effects of some insecticides on the honeybee. Agrokhimiya 8, 121–125 (1991).
Cresswell, J. E., Robert, F.-X. L., Florance, H. & Smirnoff, N. Clearance of ingested neonicotinoid pesticide (imidacloprid) in honeybees (Apis mellifera) and bumblebees (Bombus terrestris). Pest Manag. Sci. 70, 332–337 (2014).
Fytoweb. Consulter autorisations de produits phytopharmaceutiques. Available at: https://fytoweb.be/fr/produits-phytopharmaceutiques/consulter-autorisations-de-produits-phytopharmaceutiques (Accessed: 9th January 2015).
Haber, F. Zur Geschichte des Gaskrieges. In Fünf Vorträge aus den Jahren 1920–192376–92, https://doi.org/10.1007/978-3-662-33987-9_5 (Springer Berlin Heidelberg 1924).
Rand, E. E. et al. Detoxification mechanisms of honey bees (Apis mellifera) resulting in tolerance of dietary nicotine. Sci. Rep. 5, 11779 (2015).
Amdam, G. V. et al. Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Exp. Gerontol. 39, 767–773 (2004).
Corona, M. et al. Vitellogenin, juvenile hormone, insulin signaling, and queen longevity. Proc. Natl. Acad. Sci. 104, 7128–7133 (2007).
Brandt, A., Gorenflo, A., Siede, R., Meixner, M. & Büchler, R. The neonicotinoids thiacloprid, imidacloprid, and clothianidin affect the immunocompetence of honey bees (Apis mellifera L.). J. Insect Physiol. 86, 40–47 (2016).
Christen, V., Mittner, F. & Fent, K. Molecular effects of neonicotinoids in honey bees (Apis mellifera). Environ. Sci. Technol. 50, 4071–4081 (2016).
Claudianos, C. et al. A deficit of detoxification enzymes: pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 15, 615–636 (2006).
Groot, A. P. de. Protein and amino acid requirements of the honeybee (Apis mellifica L.). (Utrecht., 1953).
Mullin, C. A., Chen, J., Fine, J. D., Frazier, M. T. & Frazier, J. L. The formulation makes the poison. Pestic. Biochem. Physiol. 120, 27–35 (2015).
R Core Team. R: A Language and Environment for Statistical Computing (2017).
Ritz, C., Baty, F., Streibig, J. C. & Gerhard, D. Dose-Response Analysis Using R. Plos One 10, e0146021 (2015).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 67 (2015).
Hothorn, T., Bretz, F. & Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 50, 346–363 (2008).
Ritz, C. Toward a unified approach to dose-response modeling in ecotoxicology. Environ. Toxicol. Chem. 29, 220–229 (2010).
Abbot, W. S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18, 265–267 (1925).
Acknowledgements
We would like to thank the Walloon Region for their funding (Bee Wallonie project) of the data analyses and drafting of this publication. We thank Lucie Mesrine for her critical and constructive review of the article and the reviewers for their input.
Author information
Authors and Affiliations
Contributions
N.S. performed the experimental part and G.S.M. performed the analyses with the help of N.S. and L.H., N.S. and G.S.M. wrote the main manuscript text. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Simon-Delso, N., San Martin, G., Bruneau, E. et al. Time-to-death approach to reveal chronic and cumulative toxicity of a fungicide for honeybees not revealed with the standard ten-day test. Sci Rep 8, 7241 (2018). https://doi.org/10.1038/s41598-018-24746-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-24746-9
This article is cited by
-
Sublethal effects of herbicides clethodim, haloxyfop-P-methyl, and their mixture on honey bee health
Apidologie (2023)
-
Combined transcriptome and metabolite profiling analyses provide insights into the chronic toxicity of carbaryl and acetamiprid to Apis mellifera larvae
Scientific Reports (2022)
-
Is acetamiprid really not that harmful to bumblebees (Apidae: Bombus spp.)?
Apidologie (2022)
-
Long-term field-realistic exposure to a next-generation pesticide, flupyradifurone, impairs honey bee behaviour and survival
Communications Biology (2021)
-
A natural deep eutectic solvent as a novel dispersive solvent in dispersive liquid-liquid microextraction based on solidification of floating organic droplet for the determination of pesticide residues
Analytical and Bioanalytical Chemistry (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
