Extending Hyperspectral Imaging for Plant Phenotyping to the UV-Range


Abstract
:1. Introduction
2. Materials and Methods
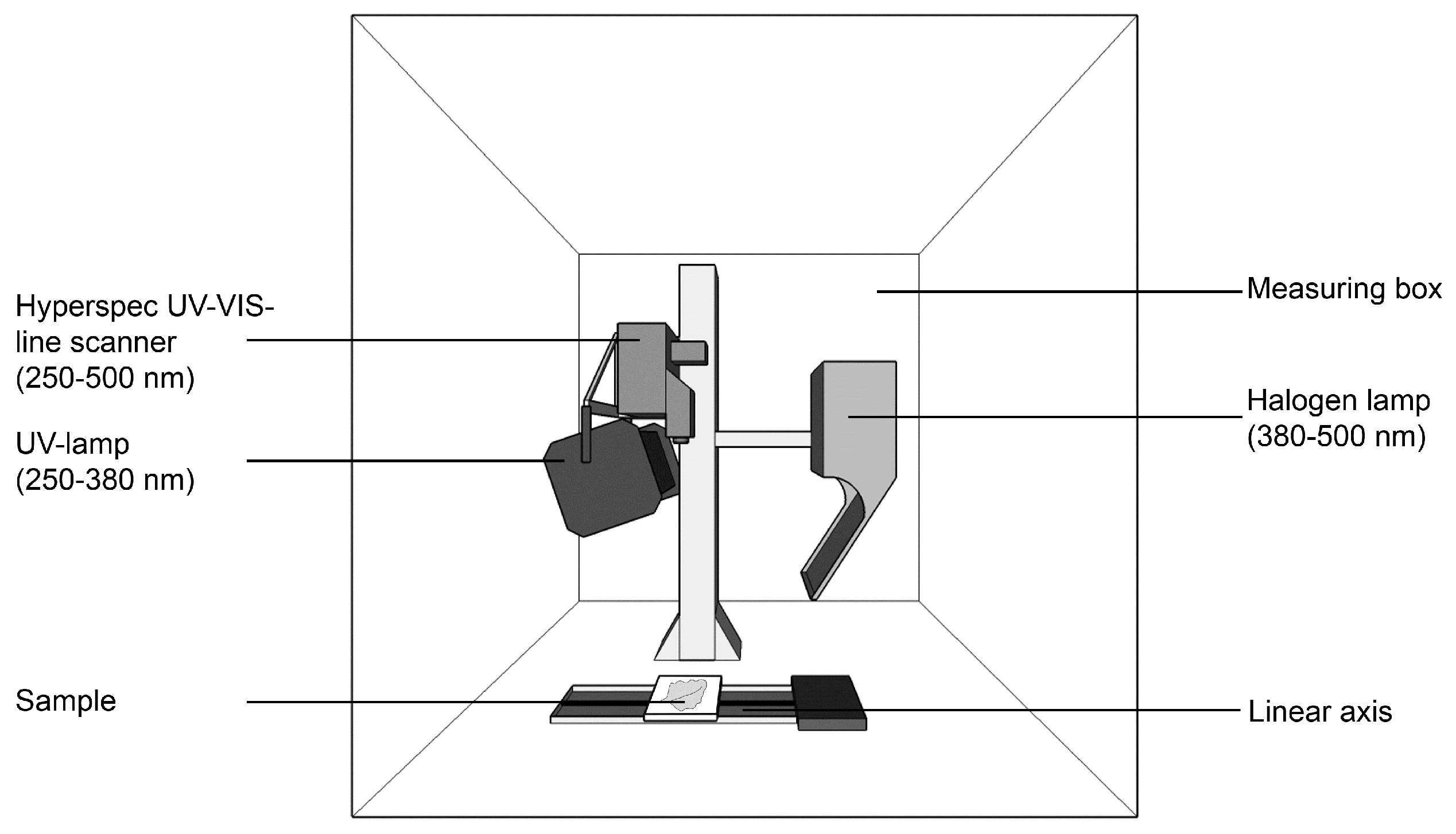
2.1. Imaging Setup and Hyperspectral Imaging
2.2. Qualitative Assessment of UV-Light Plant Tissue Interaction
2.3. Evaluation of Spectral Accuracy
2.4. Monitoring Abiotic Salt Stress of Barley Leaves in the UV-Vis Spectrum
3. Results
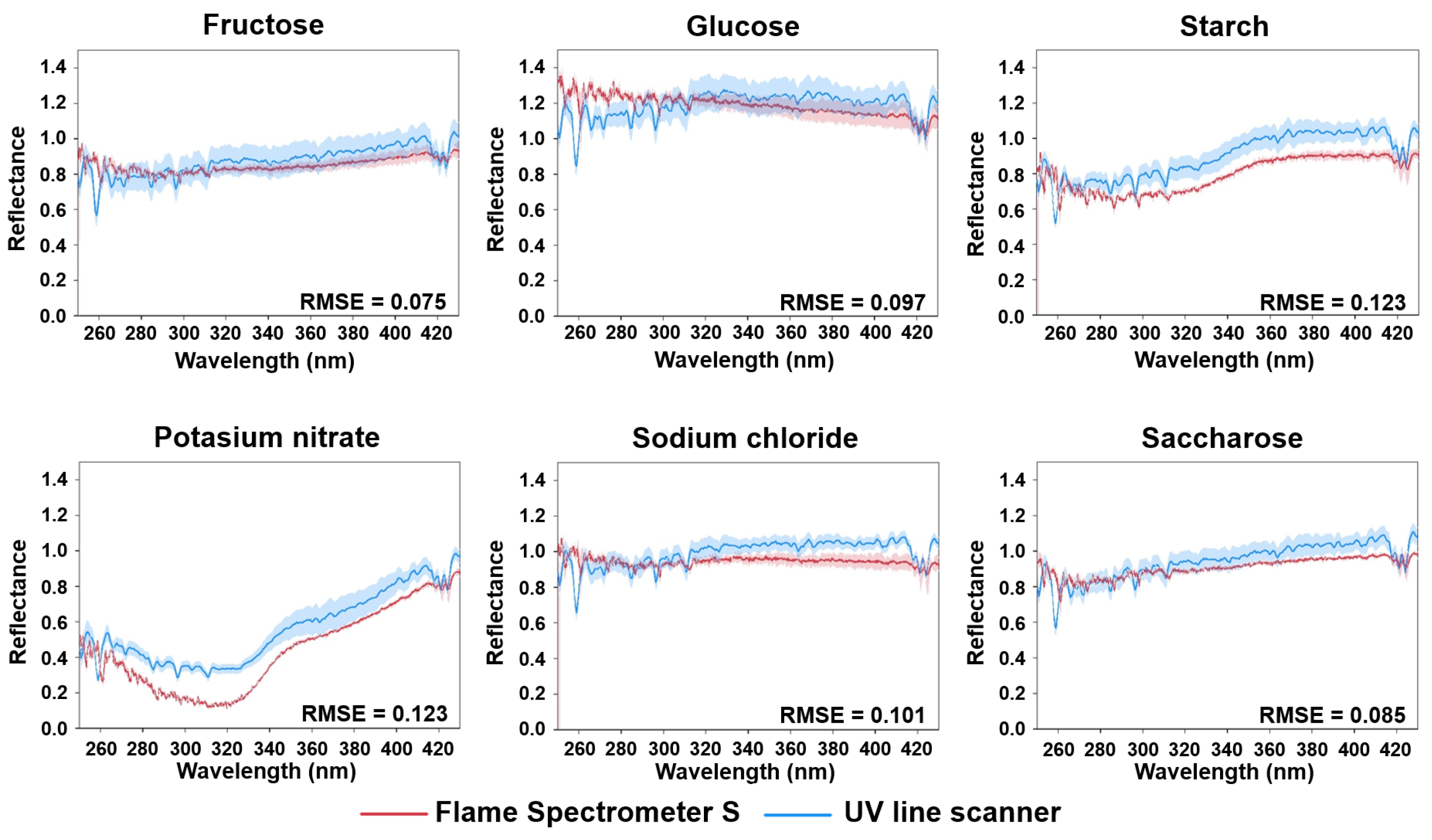
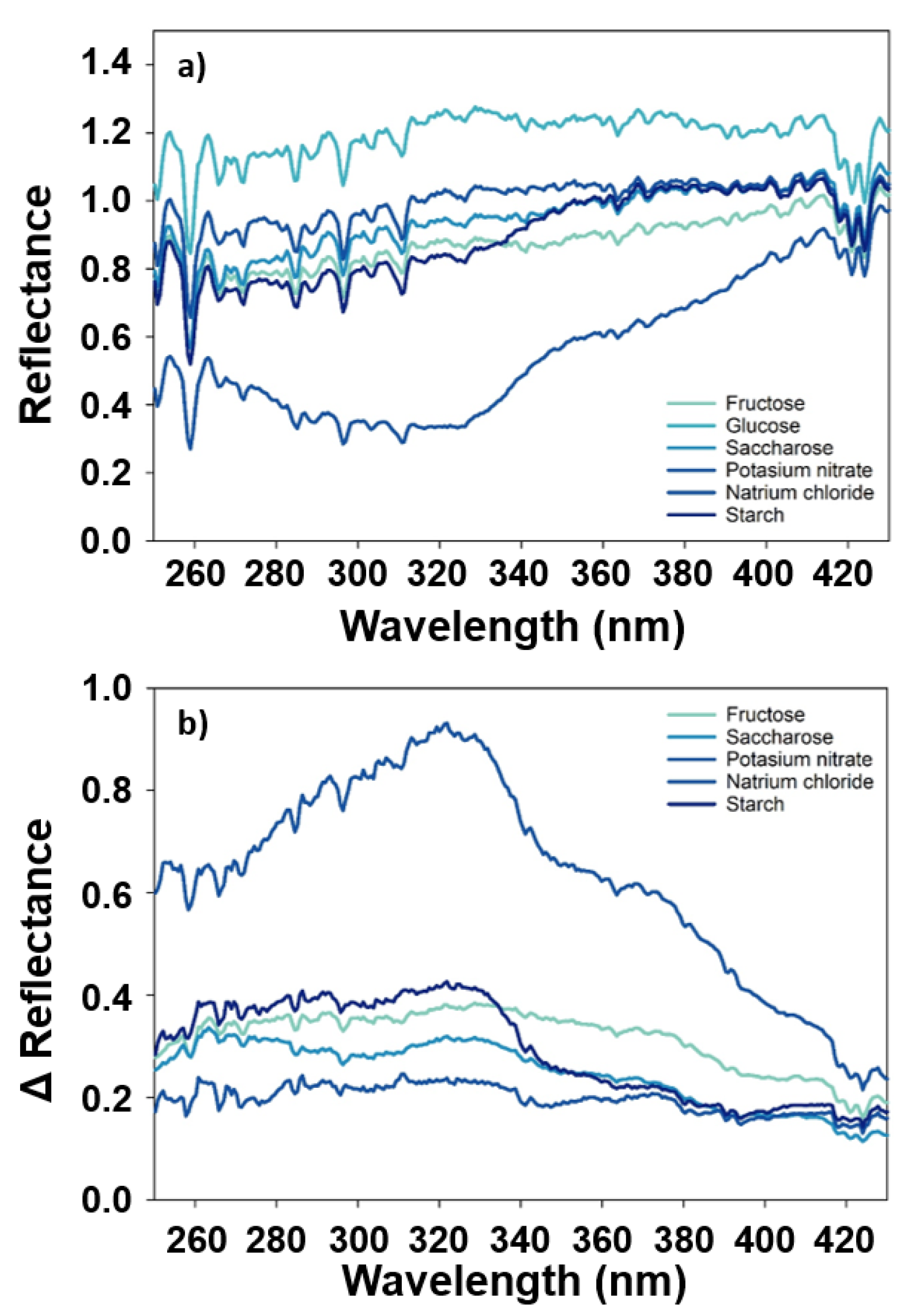
3.1. Comparison of Measurement Systems Observing Different Substances
3.2. Investigation of Phototoxicity of UV-Light on Plants
3.3. Abiotic Salt Stress of Barley Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| dai | days after incubation |
| DAB | 3,3-diaminobenzidine |
| NIR | near-infrared spectroscopy |
| RMSE | root-mean-square error |
| ROS | reactive oxygen species |
| SWIR | short-wavelength infrared |
| UV | ultraviolett |
| VIS | visual spectrum |
Appendix A

References
- Mahlein, A.K.; Kuska, M.T.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral sensors and imaging technologies in phytopathology: State of the art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Kuska, M.T.; Bohnenkamp, D.; Brugger, A.; Alisaac, E.; Wahabzada, M.; Behmann, J.; Mahlein, A.K. Benefits of hyperspectral imaging for plant disease detection and plant protection: A technical perspective. J. Plant Dis. Prot. 2018, 125, 5–20. [Google Scholar] [CrossRef]
- Carter, G.A.; Knapp, A.K. Leaf optical properties in higher plants: Linking spectral characteristics to stress and chlorophyll concentration. Am. J. Bot. 2001, 88, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Slaton, M.R.; Raymond Hunt, E., Jr.; Smith, W.K. Estimating near-infrared leaf reflectance from leaf structural characteristics. Am. J. Bot. 2001, 88, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Jacquemoud, S.; Ustin, S.L. Leaf optical properties: A state of the art. In Proceedings of the 8th International Symposium of Physical Measurements & Signatures in Remote Sensing, CNES, Aussois, France, 8–12 January 2001; pp. 223–332. [Google Scholar]
- Kuska, M.T.; Behman, J.; Mahlein, A.K. Potential of hyperspectral imaging to detect and identify the impact of chemical warfare compounds on plant tissue. Pure Appl. Chem. 2018, 9, 1615–1624. [Google Scholar] [CrossRef]
- Curran, P.J. Remote sensing of foliar chemistry. Remote Sens. Environ. 1989, 30, 271–278. [Google Scholar] [CrossRef]
- Fischer, J. Specific detection of nucleotides, creatine phosphate, and their derivatives from tissue samples in a simple, isocratic, recycling, low-volume system. LC GC Int. 1995, 8, 254. [Google Scholar]
- Giusti, M.M.; Polit, M.F.; Ayvaz, H.; Tay, D.; Manrique, I. Characterization and quantitation of anthocyanins and other phenolics in native Andean potatoes. J. Agric. Food Chem. 2014, 62, 4408–4416. [Google Scholar] [CrossRef] [PubMed]
- Talrose, V.; Yermakov, A.; Usov, A.; Goncharova, A.; Leskin, A.; Messineva, N.; Trusova, N.; Efimkina, M. UV/visible spectra. In NIST Chemistry WebBook, NIST Standard Reference Database; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2009; p. 20899. [Google Scholar]
- Bhattacharya, A.; Sood, P.; Citovsky, V. The roles of plant phenolics in defence and communication during Agrobacterium and Rhizobium infection. Mol. Plant Pathol. 2010, 11, 705–719. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; García-Viguera, C.; Tomás-Barberán, F. On-line identification of flavonoids by HPLC coupled to diode array detection. Methods Polyphen. Anal. 2003, 92, 127. [Google Scholar]
- Ali, R.; Abbas, H. Response of salt stressed barley seedlings to phenylurea. Plant Soil Environ. 2003. [Google Scholar] [CrossRef]
- Hollósy, F. Effects of ultraviolet radiation on plant cells. Micron 2002, 33, 179–197. [Google Scholar] [CrossRef]
- A.-H.-Mackerness, S.; John, C.F.; Jordan, B.; Thomas, B. Early signaling components in ultraviolet-B responses: Distinct roles for different reactive oxygen species and nitric oxide. FEBS Lett. 2001, 489, 237–242. [Google Scholar] [CrossRef]
- Van Breusegem, F.; Dat, J.F. Reactive oxygen species in plant cell death. Plant Physiol. 2006, 141, 384–390. [Google Scholar] [CrossRef]
- Tsormpatsidis, E.; Henbest, R.; Battey, N.; Hadley, P. The influence of ultraviolet radiation on growth, photosynthesis and phenolic levels of green and red lettuce: Potential for exploiting effects of ultraviolet radiation in a production system. Ann. Appl. Biol. 2010, 156, 357–366. [Google Scholar] [CrossRef]
- Witzenberger, A.; Hack, H.; Van Den Boom, T. Erläuterungen zum BBCH-Dezimal-Code für die Entwicklungsstadien des Getreides—Mit Abbildungen. Gesunde Pflanze 1989, 41, 384–388. [Google Scholar]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—Powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Wolf, G.; Fric, F. A rapid staining method for Erysiphe Graminis F. sp. hordei Whole Barley Leaves A Protein-Specif Dye. Phytopathology 1981, 71, 596–598. [Google Scholar]
- Kuska, M.; Wahabzada, M.; Leucker, M.; Dehne, H.W.; Kersting, K.; Oerke, E.C.; Steiner, U.; Mahlein, A.K. Hyperspectral phenotyping on the microscopic scale: Towards automated characterization of plant-pathogen interactions. Plant Methods 2015, 11, 28. [Google Scholar] [CrossRef]
- Nawkar, G.; Maibam, P.; Park, J.; Sahi, V.; Lee, S.; Kang, C. UV-induced cell death in plants. Int. J. Mol. Sci. 2013, 14, 1608–1628. [Google Scholar] [CrossRef] [PubMed]
- Gadjev, I.; Stone, J.M.; Gechev, T.S. Programmed cell death in plants: New insights into redox regulation and the role of hydrogen peroxide. Int. Rev. Cell Mol. Biol. 2008, 270, 87–144. [Google Scholar]
- Gao, C.; Xing, D.; Li, L.; Zhang, L. Implication of reactive oxygen species and mitochondrial dysfunction in the early stages of plant programmed cell death induced by ultraviolet-C overexposure. Planta 2008, 227, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Mahlein, A.K.; Hammersley, S.; Oerke, E.C.; Dehne, H.W.; Goldbach, H.; Grieve, B. Supplemental Blue LED Lighting Array to Improve the Signal Quality in Hyperspectral Imaging of Plants. Sensors 2015, 15, 12834–12840. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Prášil, I.; Vítámvás, P. Protein contribution to plant salinity response and tolerance acquisition. Int. J. Mol. Sci. 2013, 14, 6757–6789. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Cai, S.; Chen, M.; Ye, L.; Chen, Z.; Zhang, H.; Dai, F.; Wu, F.; Zhang, G. Tissue metabolic responses to salt stress in wild and cultivated barley. PLoS ONE 2013, 8, e55431. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Annunziata, M.G.; Pontecorvo, G.; Fuggi, A.; Woodrow, P. Salinity stress and salt tolerance. In Abiotic Stress in Plants-Mechanisms and Adaptations; InTech: Vienna, Austria, 2011. [Google Scholar]
- Parida, A.; Das, A. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Clark, R.N.; Roush, T.L. Reflectance spectroscopy: Quantitative analysis techniques for remote sensing applications. J. Geophys. Res. 1984, 89, 6329–6340. [Google Scholar] [CrossRef]
- Wang, H.; Ma, F.; L, C. Metabolism of organic acids, nitrogen and amino acids in chlorotic leaves of ’Honeycrisp’ apple (Malus Domest. Borkh) Excessive Accumul. Carbohydrates. Planta 2010, 232, 511–522. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, M.; Ruan, J. Metabolomics analysis reveals the metabolic and functional roles of flavonoids in light-sensitive tea leaves. BMC Plant Biology 2017. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, A.; Takahashi, T. Procyanidin B-3, isolated from barley and identified as a hair-growth stimulant, has the potential to counteract inhibitory regulation by TGF-beta1. Exp. Dermatol. 2002, 11, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Gausman, H. Leaf reflectance vs. leaf chlorophyll and carotenoid concentrations for eight crops. Agron. J. 1977, 69, 799–802. [Google Scholar] [CrossRef]
- Okoronkwo, N.E.; Kalu, C.M.; Nnorom, I.C. Estimation of protein content and amino acid compositions in selected plant aamples using UV-Vis spectrophotometeric method. Am. J. Food Sci. Health 2017, 3, 41–46. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Flame Spectrometer S | UV Line Scanner |
|---|---|---|
| manufacturer | Ocean Optics | Headwall Photonics |
| sensor type | non-imaging | imaging |
| wavelength range | 190–1100 nm | 250–500 nm |
| spectral resolution | 0.11 nm | 14 nm (961 bands) |
| spatial resolution | - | 1392 px |
| frame rate | 400 Hz | 7 Hz |
| movement speed between x- and y-axis | - | 0.141 mm/s |
| measuring software | Flame 1.6.7 | Hyperspec III |
| image sensor name | - | Intevac Microvista Kamera |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brugger, A.; Behmann, J.; Paulus, S.; Luigs, H.-G.; Kuska, M.T.; Schramowski, P.; Kersting, K.; Steiner, U.; Mahlein, A.-K. Extending Hyperspectral Imaging for Plant Phenotyping to the UV-Range. Remote Sens. 2019, 11, 1401. https://doi.org/10.3390/rs11121401
Brugger A, Behmann J, Paulus S, Luigs H-G, Kuska MT, Schramowski P, Kersting K, Steiner U, Mahlein A-K. Extending Hyperspectral Imaging for Plant Phenotyping to the UV-Range. Remote Sensing. 2019; 11(12):1401. https://doi.org/10.3390/rs11121401
Chicago/Turabian StyleBrugger, Anna, Jan Behmann, Stefan Paulus, Hans-Georg Luigs, Matheus Thomas Kuska, Patrick Schramowski, Kristian Kersting, Ulrike Steiner, and Anne-Katrin Mahlein. 2019. "Extending Hyperspectral Imaging for Plant Phenotyping to the UV-Range" Remote Sensing 11, no. 12: 1401. https://doi.org/10.3390/rs11121401