
. Methodological context
The values of 14C dates used for the construction of prehistoric narratives are dictated by the precision and accuracy of radiocarbon determinations, both assessed in the context of the target events. Their usability is ultimately not determined on the basis of the quality of the 14C measurements themselves, but rather on the precision of the calibrated age range determination derived from them. Even if the causal and temporal relationships between the relevant archaeological event and the dated material are determined correctly, and the dates obtained do indeed correspond to the relevant moment in the past, it may still be the case that radiocarbon dates prove to be of little weight for the needs of a specific (pre)historic research question.
Of particular importance are the plateaus and wiggles on radiocarbon calibration curves. These may result in extending even the most precise and accurate 14C determination, spreading it over potentially very long stretches of time and often pointing at few or even several calendar periods. In this case, modelling of the 14C dates of temporarily sequenced samples (not necessarily solely 14C dates) using Bayesian statistics can increase the accuracy and may provide prehistorically conclusive and useful results.
Bayesian chronometric models can be based on externally pre-ordered (extrinsic) noosphere-related sequences of 14C dates. An example is a burial within a series of graves chronologically prearranged using archaeological methods. Other examples are a series of 14C dates temporally ordered by bio-spheric-related (intrinsic) natural sequences (e.g. tree-rings).
Drawing from forensic studies (cf. e.g. Alkass et al., 2011; Brock and Cook 2016; Calcagnile et al., 2013; Cook et al., 2015; Cook and MacKenzie 2014; and Ubelaker et al., 2006, 2015), we propose the Bayesian modelling of multiple 14C dates from single human skeletons, assuming that individual bone fragments can be defined as heterochronous 14C reservoirs. The dates derived from such ‘stratified’ samples create intrinsic ontogenetic sequences with gaps, the span of which may be estimated by anthropological methods.
. Archaeological context
The starting point for this brief chronometric study was the difficulty in establishing a satisfactorily precise chronology of inhumation discovered at the site of Mikulin, in the Lublin Province of eastern Poland. For comparative purposes, a single human burial from a large cemetery excavated at the Romanian site of Urziceni-Vada Ret (the formerly used toponym was Urziceni-Vamă), in the Satu Mare County (Fig. 1), was included in the analyses. These cases will be the subject of comparisons only on a methodological level. In no way are we going to search for the existence of a possible prehistoric ‘common denominator’. Nevertheless, before proceeding to the focal chronometric questions, we will briefly present both of the studied cases as prehistoric issues with a particular emphasis put on their relevant chronological problems.
Fig 1
Localisation of archaeological sites discussed in the text: the Funnel Beaker culture cemetery at site 9, Mikulin (with an adjacent settlement at site 1, Dobużek Kolonia ) and the Bodrogkeresztúr culture cemetery at the site of Urziceni-Vada Ret (based on a map of M. Juran).
The burial from Mikulin
The inhumation cemetery discovered at site 9 in Mikulin can be attributed to the Funnel Beaker culture exclusively, based on the analysis of funeral rites and the scarce material finds there. Certain topographical conditions, based on comparative materials, indicate that there was an extensive necropolis located around monumental earthen long barrows. To date, only a small part of the cemetery has been explored, within which a dozen graves have been unveiled. Of these, one of the two skeletons deposited in the pit marked as feature 9 (Fig. 2) was designated for dating. Using the standards proposed by Buikstra and Ubelaker (1994), the individual was identified as an adult female aged 30–35, based on the diagnostic pelvic bone and skull morphology.
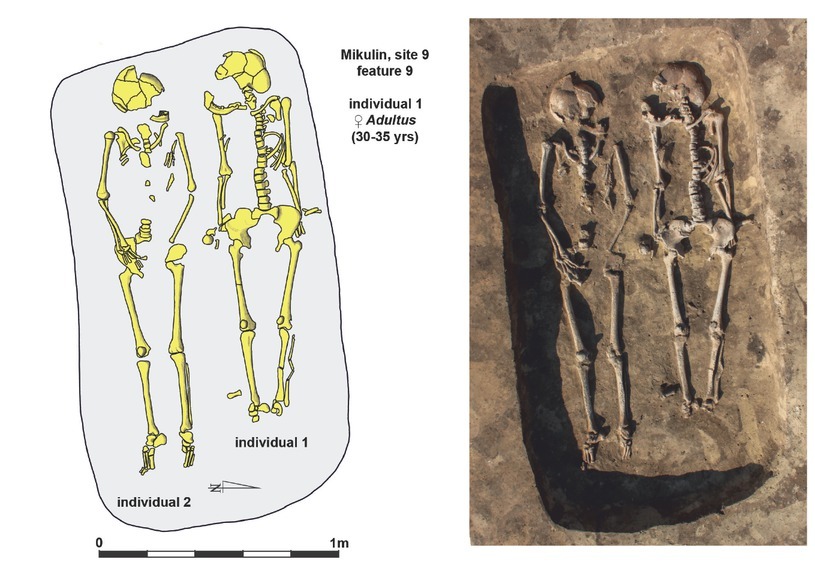
The most serious difficulty stems from the lack of grave goods and a lack of stratification between funerary features discovered within the burial ground. These precluded the use of classical archaeological methods for sequencing episodes within the cemetery. Considering the specificities of burying customs of the Funnel Beaker culture in this region, the unfavourable dating situation is not expected to improve due to possible continuation of excavations at the site. In view of the absence of any archaeologically defined elements, which would allow for a more precise temporal attribution of particular graves and the whole cemetery, we were forced to rely completely on the radiocarbon chronology of individual burials.
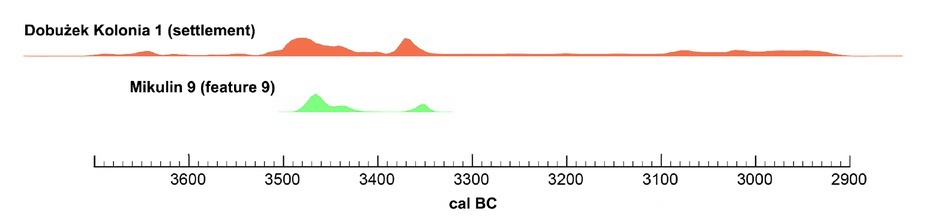
The site 9 cemetery at Mikulin is located in the immediate vicinity of the settlement from the same period (Dobużek Kolonia, site 1). The chronometric conclusions for the findings from the settlement allow its occupation to be dated between the 37th and 31st centuries BC (unpublished chronometric analyses). Within this period, a few significant episodes had already been defined, for which a precise radiocarbon chronology was established; in some cases, this was narrowed down to a period as short as half a century. Therefore, our aim was to most accurately determine the chronological relationships between the cemetery and the settlement.
The burial from Urziceni-Vada Ret
The cemetery discovered at the Vada Ret site in Urziceni constitutes the remnant of a vast burial field associated with the eneolithic Bodrogkeresztúr culture. So far, more than one hundred burials have been excavated and studied, representing the vast majority of the temporal span of this cultural formation (cf. Virag, 2018). In accordance with the assumptions presented here, the skeletal remains of a single burial (grave 39; Fig. 3) were subjected to 14C dating. The skeleton was identified as a mature female aged 35–45, based on the diagnostic pelvic bone and skull morphology.

In contrast to the funerary rites of the Funnel Beaker culture communities, bearers of the Bodrogkeresztúr culture laid their deceased in richly furnished graves. The existence of such large cemeteries as the one found in Urziceni allows for the effective application of basic archaeological dating techniques, such as cross-dating and seriation. Nonetheless, until the site is fully excavated, seriation analyses remain incomplete; the basis for the present estimation of the chronology of individual burials is provided by the typological premises and particular radiocarbon dating. However, the assessment of the chronology of the cemetery in Urziceni by means of 14C dates obtained so far is hindered by the presence of a large plateau on the calibration curve extending between 4220–4060 BC, a range in which all these measurements fall in. Additionally, the presence of certain typologically dated elements seemed to contradict the pre-determined radiocarbon age of the burials. At this stage of the research, we wanted to make an ad hoc attempt at getting a more precise chronometric decision for the grave considered as one of these problematic burials, supposing that the typo-chronological discrepancies could be a result of the adoption of erroneous presumptions about the age of some finds.
. Aims, Methods, Materials
Selection of samples
The immediate goal of the reported chronometric exercises was to model a sequence of 14C dates obtained for defined heterochronous ontogenetic deposits of collagen within single human skeletons. Due to budgetary constraints, we decided to stop at radiocarbon paired sequences, consisting of radiocarbon age determination for a short period of an individual’s development involving foetal life and the first half of early childhood (henceforth globally referred to as ‘very early childhood’) and the other one related to the shortest possible period prior to death (referred to as ‘terminal period’).
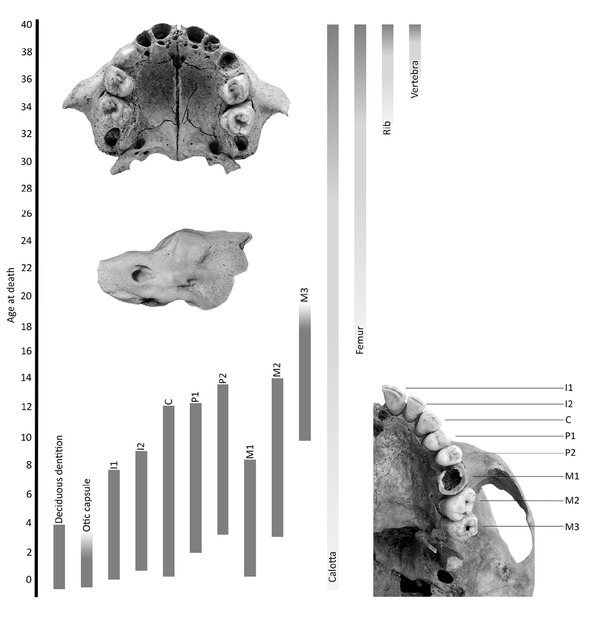
Taking into account the ontogenetic age of the particular human skeletal elements, we considered the petrous part of the temporal bone (and more specifically the otic capsule) as the best representative of radiocarbon age for the period covering foetal life and the very early childhood of a given individual. Its development is completed in the second year of life, and from this stage, this extremely hard bone tissue does not undergo any transformation. Consequently, the collagen deposited in the osseous labyrinth becomes an isotopic ‘time capsule’ of this very early period of ontogenesis (cf. e.g. Harvig et al., 2014 and Scheuer and Black, 2004). On the contrary, as a consequence of their intense remodelling almost throughout the lifetime of an individual, collagen turnover rate for ribs is very high (see Fahy et al., 2017), and therefore, they should yield a radiocarbon age closely corresponding to the terminal period of the individual’s life (Fig. 4).
Fig 4
Ontogenetic time of development and remodelling of specific human bones and teeth (based on Tütken, 2010; modified).
Importantly, the available skeletal remains under study had to provide reliable reservoirs of both target ‘ontogenetic deposits’ of collagen. It was precisely on such grounds that the human remains from Mikulin and Urziceni described above were selected. However, while the skeleton from the first cemetery mentioned was sampled in accordance with the above-described anatomical preferences, in the case of Urziceni, a previously existing radiocarbon date obtained from a fragment of tibia was used to determine the radiocarbon age of the terminal period of the individual’s life.
Preliminary simulations showed that our interest should concentrate primarily on skeletons of individuals who died at as advanced ages as possible. Only then could the calibrated 14C dates of samples from very early childhood (together with the anthropologically defined gap constituting our prior probabilities) largely influence the posterior probabilities of 14C dates related to terminal periods of their lives and, consequently, improve the precision of absolute dating of the bodies’ inhumation. Thus, the age of individuals at the time of death was an important criterion for the selection of these case studies.
Assessment of samples
One of the basic problems was to ensure the accuracy of 14C determinations to be made and thus their convergence with the target past events.
The prerequisite for obtaining such accurate 14C dates was the exclusion of samples loaded with diagenetic impurities, or the removal of such possible contaminations from samples qualified for dating. The collagen content (%coll) in bones, the nitrogen content (%N) and the atomic carbon-to-
nitrogen ratio (C/N) in the collagens were held as essential indicators of samples not suitable for dating (cf. Brock et al., 2010 and van Klinken, 1999). In all the laboratories in which the dating was performed, an appropriate ‘radiometric hygiene’ of the samples qualified was ensured by following relevant pre-treatment protocols of collagen, generally compatible with those described by Fiona Brock et al., (2010).
No less problematic would be a possible sampling of bones burdened with older extrinsic carbon as a result of consumption of aquatic organisms by given individuals (see e.g. Philippsen, 2013). To exclude this possibility, the diet of the deceased was assessed against the possible consumption of water-originating foodstuffs by determining the composition of stable isotopes of carbon (13C/12C) and nitrogen (15N/14N) in each of the collagen samples to be dated (expressed by δ13C and δ15N).
Additionally, to provide a firm isotopic background for such inferences (cf. Goslar et al., 2018), in the case of the burial from Mikulin, we decided to determine δ13C and δ15N for bones of herbivores (Bos taurus, Ovis/Capra and Equus sp.) consumed by the human group the individual belonged to (faunal remains from the aforementioned settlement of Dobużek Kolonia, adjacent to the Mikulin cemetery). Regrettably, no such reference samples were analysed for Urziceni.
Dating
In general, modelling with the use of Bayesian statistics allows for the efficient use of even relatively low-precision 14C dates (see e.g. Hamilton and Krus, 2018). However, the presumed ‘binary’ design of the proposed sequential models does not incorporate any radiocarbon dates that would chronologically be ahead of the hereby understood very early childhood or posterior to the terminal period. It was therefore to be expected that the precision of the particular conventional 14C ages would also have a significant impact on the posterior probabilities of the modelled dates.
A common solution to improve measurement precision is to replicate them. This was adopted in the case of dating the skeleton of Mikulin. Each of the two bone samples was AMS dated twice – by the Oxford Radiocarbon Accelerator Unit (ORAU) and Poznań Radiocarbon Laboratory (Poz). Additionally, in the former case, dating of one sample was performed with the use of two carbon cathodes, what yielded double 14C age determinations for the otic capsule. However, for financial reasons, it was not possible to obtain multiple measurements for the skeleton from Urziceni. Thus, for the latter burial, we only had two radiocarbon determinations – one for the petrous pyramid, obtained for the purposes of palaeogenetic studies (cf. Mathieson et al., 2018) in the laboratory of the Pennsylvania State University (PSUAMS), and the other for tibia by the Hertelendi Laboratory of Environmental Studies in Debrecen (DeA).
The specifications of the samples, including the basic isotopic parameters of their radiocarbon homogeneity or lack thereof, as well as the results of the radiocarbon measurements obtained for each sample, were compiled in accordance with the modern 14C dates’ reporting conventions (cf. Bayliss, 2015 and Millard, 2014).
Modelling
OxCal version 4.3 (Bronk Ramsey, 2009a) and IntCal13 calibration curve (Reimer et al., 2013) were used to calibrate the results of radiocarbon measurements and to create a model that complied with the assumptions made. Our goal was to build simple two-element sequences, based on the radiocarbon dates of the birth and death of individuals, with a defined gap between these events [V_Sequence; cf. Bronk Ramsey, 2008].
The age determinations for both ontogenetically defined collagen deposits within the skeleton of the grave from Urziceni were based on the single radiocarbon measurements and as such were incorporated into the model directly. In the case of Mikulin, however, the age determinations destined for modelling consist of combinations [Combine] of multiple dates obtained for particular bone fragments.
As both the skeletons were of individuals who died at quite an advanced age, when bone remodelling rates become lower, the 14C determinations obtained for the terminal deposit of collagen from the dated skeletons corresponded to moments somewhat preceding the death and inhumation of the individuals. Due to the lack of estimations of the radiocarbon lag based on specific anatomical fragments, for the dates obtained by us from ribs and tibia, we applied a general correction to the date of death as studied by Mebus Geyh (2001). In reference to his study, Barta and Štolc, (2007) introduced a concept of human bone collagen offset (HBCO), calculated its values for several classes of skeletal age (the age at which an individual died) and proposed implementing HBCOs by means of the Delta_R command available in many 14C calibration tools. However, the Delta_R, widely applied when correcting 14C dates from marine environments (where 14C concentration is lower with respect to that in the atmosphere, making radiocarbon ages older by the so-called reservoir age), concerns the domain of radiocarbon timescale. Conversely, the fact that the collagen in a bone contains carbon derived from the environment over a considerable period of time preceding the moment of organism’s death should be corrected for in the domain of calendar time. In the language of OxCal [cf. Bronk Ramsey, 2009b], such a kind of correction is rather approximated by the command Reservoir, with parameter ‘tau’ representing the length of the aforementioned time period. Calibrating 14C ages of ribs and tibia of individuals from Mikulin and Urziceni studied in the present work, we applied this particular OxCal command, adopting for values of tau the HBCOs of 14.01±1.94 and 21.21±1.67 as calculated by Barta and Štolc (2007, see Table 2 therein) for the adult and the mature skeletal ages, respectively.
Table 1
Urziceni-Vada Ret, Satu Mare County, Mikulin, site 9, and Dobużek Kolonia, site 1, Lublin Province. Archaeological context and specification of samples used for 14C dating and dietary studies.
| Site | feature/sampled individual | dated bone(s) | Laboratory code | Age (14C BP) | coll% | δ13C (‰) | δ15N (‰) | %C | %N | (C/N)at |
|---|---|---|---|---|---|---|---|---|---|---|
| Urziceni-Vada ret | grave 39/female (Maturus, ca. 40-45 yrs) | petrous part | PSUAMS 4229 | 5300±25 | 10.3 | -20.4b) | 12.0 | 28.1d) | 10.2d) | 3.2d) |
| tibia | DeA-17598 | 5334±33 | 1.0 | -19.7b) | 10.9 | 39.7d) | 13.6d) | 3.4d) | ||
| Mikulin, site 9 | feature 9 (grave)/individual 1: female (Adultus, ca. 30-35 yrs) | petrous part | Poz-105744a) | 4596±37 | 1.4 | -17.4b) | n.m. | 8.5c) | 1.6c) | 3.2d) |
| (burial ground) | Poz-105745a) | 4661±37 | 48.2d) | 17.6d) | ||||||
| OxA-38015 | 4699±31 | 5.6 | -19.4b) | 12.0 | 45.6d) | 16.1d) | 3.3d) | |||
| ribs | Poz-104804 | 4635±30 | 13.1 | -19.6b) | 10.2 | 8.0c) | 3.2c) | 3.2d) | ||
| 49.5d) | 18.1d) | |||||||||
| OxA-38219 | 4614±30 | 10.9 | -19.9b) | 10.5 | 46.6d) | 16.4d) | 3.3d) | |||
| Dobużek Kolonia, site 1(settlement) | feature 24/Bos taurus sp. | humerus | Poz-105439 | 4635±35 | 3.5 | -21.1b) | 7.9 | 6.4c) | 1.4c) | 3.2d) |
| 50.5d) | 18.5d) | |||||||||
| feature 38/ Ovis/ | ribs | Poz-105447 | 4590±35 | 3.8 | -20.3b) | 8.8 | 5.1c) | 1.1c) | 3.2d) | |
| Capra sp. | 50.4d) | 18.5d) | ||||||||
| feature 42/ | pelvic bone | Poz-105529 | 4680±40 | 3.1 | -21.3b) | 4.8 | 8.4c) | 0.8c) | 3.2d) | |
| Equus sp. | 49.4d) | 18.1d) |
a) the two radiocarbon dates were originally reported as a single one as their weighted mean (Poz-105744: 4628±26 BP)
Table 2
Results of calibration and modelling of 14C dates (rounded by 5 years).
As mentioned above, the gap between the elements of the sequence was defined each time on the basis of the anthropological estimates of the age of the individuals at the time of death. However, in terms of precision, the values obtained by applying anthropological methods had to be adapted to the existing possibilities of modelling variable sequences. Because of the imprecision of the calibration curve currently in use, OxCal imposes a minimum error range of ±5 years. The individual buried in Urziceni did not present any difficulties in this regard for the Maturus age class can be expressed as a gap of 40±5 years. In the case of Mikulin, though, we were forced to use a less precise definition of the life time span (gap=30±5 yrs) than the anthropological analyses would allow to (32.5±2.5 yrs).
. Results
Dating of the burial from Mikulin
Despite slight inter-laboratory differences, the basic parameters of collagen (%coll, C/N ratio) extracted from the bones of the individual buried in feature 9 in Mikulin allowed to exclude the possible bias of resulting 14C dates due to their diagenetic contamination (Table 1).
The results of the stable isotopic analyses demonstrate general inter-laboratory coherence (Table 1). Values of δ15N for ribs of the buried woman appear to be just 1.5–2.5‰ higher than those obtained for bone remains (from Dobużek Kolonia) of herbivore animals herded or hunted by the human group the individual apparently belonged to. As δ15N values increase by some 3.5‰ with every link in the food chain, the values of δ15N for the human ribs seem to prove that the diet of the woman in the terminal period of life was based mainly on the terrestrial plants and the meat of herbivores, with no detectable share of aquatic foodstuff. A somewhat higher value of δ15N (12.0‰) was recorded for the petrous bone of the same individual, pointing to the existence of one more intermediate link in the food chain. It is easy to explain, as the individual at the stage of very early childhood must have been breastfed. In conclusion, these results allow the exclusion of any freshwater reservoir effect on the 14C dates obtained for the bone tissues and indirectly confirm the ontogenetic heterochronicity of the collagen samples.
Individual conventional dates (Table 1) are generally in line with the expectations based on the current knowledge about the chronology of analogous prehistoric finds in this region (cf. e.g. Chmielewski, 2015). They also fit the interval of summed radiocarbon dates for the settlement sequence of the Funnel Beaker culture from Dobużek Kolonia, what confirms the correctness of assumption about contemporaneity and relatedness of the latter with the burial ground.
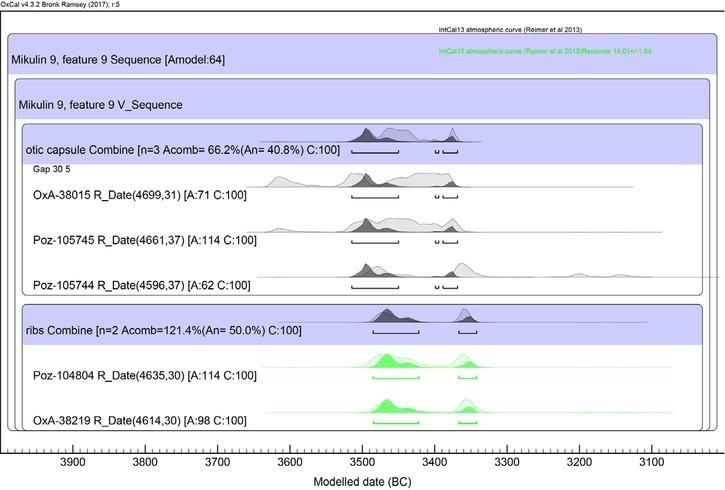
The agreement indexes for the chronological model (Amodel≈64; Aoverall≈77) as well as for the individual measurements are above thresholds of acceptance (Figs. 5 and 6). Following the modelling, the radiocarbon chronology of the terminal period of the individual’s life was almost completely narrowed down to the confidence interval of approximately 3480–3440 BC (59.2 of 68.2%; Table 2). This allows accurate and precise synchronisation of the cemetery with the early stage of the development of the village (Fig. 7).
Fig 5
Mikulin, Lublin Province, site 9 – feature 9, individual 1. Specification of model based on the series of 14C dates obtained for ribs and otic capsule (cf. Table 1).

Dating of the burial from Urziceni-Vada Ret
Similar to the burial from Mikulin, the basic parameters of collagen samples (%coll, C/N ratio) taken from the petrous bone and tibia of the examined individual from Urziceni did not reveal any diagenetic contamination of the dated material (Table 1).
Although no bones of herbivore animals used by the Bodrogkeresztúr culture population from Urziceni were provided, both the δ15N value obtained for tibia of the buried woman (10.9‰) and the predictably slightly higher δ15N for otic capsule (12.0‰) are very close to the corresponding numbers for Mikulin. This strongly suggests that aquatic environments played no important role as a source of food of the individual and that her diet did not affect 14C ages of the sampled bones.
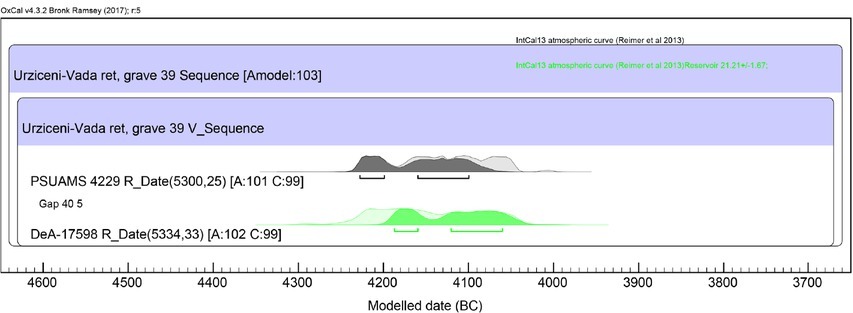
The agreement indexes of the chronological model (Figs. 8 and 9) are relatively high (Amodel≈103; Aoverall≈102). Nonetheless, due to the course of the calibration curve (the presence of a large plateau), the outcome of the modelling is less satisfying than for the grave from Mikulin. The primary confidence interval of the 14C date obtained for tibia of about 4220–4060 BC is narrowed down to about 4190–4060 BC (68.2%), which corresponds to some 20% reduction in the length of the interval corresponding with the burying of the individual.
Fig 8
Urziceni-Vada Ret, Satu Mare County – grave 39. Specification of model based on two 14C dates obtained for tibia and otic capsule (cf. Table 1).

Fig 9
Urziceni-Vada Ret, Satu Mare County – grave 39. Multiplot based on the model specified in the previous figure.
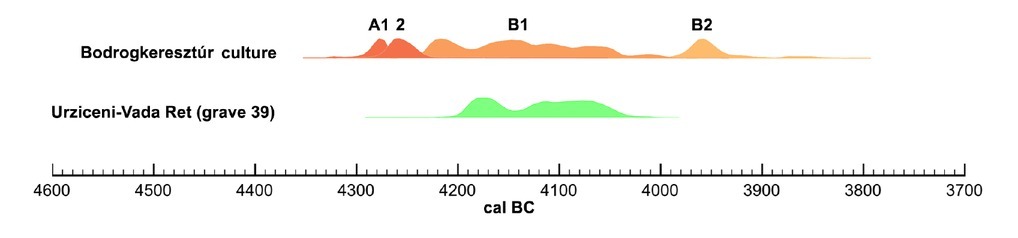
The dating results, both if reported conventionally and when modelled (see Table 2), provide us with the absolute age of the grave 39 from Urziceni that is in accordance with the current chronology of similar burials, placing moment of deposition of the individual dated at the stage B1 of the Bodrogkeresztúr culture (Fig. 10).
Fig 10
Synchronisation of the radiocarbon dated burial from Urziceni with the continuous sequence of phases of the Bodrogkeresztúr culture (the latter based on Chmielewski 2020).
. Discussion
The archaeological literature is still being filled with radiocarbon dates and sequential models based thereon, many of which are of negligible heuristic values already at the time of publication. This is often not so much because of misinterpretation or incorrectness of the radiocarbon age determinations themselves, but rather due to their prehistoric irrelevance. Therefore, every application of the proposed approach should first be considered in terms of accuracy of age determinations for the sake of diachronicities and synchronicities that are to underpin prehistoric narrative(s) at stake. It is not worthwhile to dedicate large resources or to obtain a measurement’s resolution which is not useful due to the large inaccuracy of the chronological scale in which the narrative is to be pursued. Neither is it useful to obtain results still not precise enough for the intended prehistoric inferring.
However, the problem is not limited to the proper assessment of the possibility of obtaining correct 14C measurements and models meeting chronological expectations of archaeologists. Due to the specific characteristics of the material to be dated, it is also necessary to foresee the emergence of various operational dependencies between the course of the proposed procedure for obtaining radiocarbon dates and potential parallel bioarchaeological studies.
Fortunately, the need to integrate radiocarbon measurements with classical anthropological analyses and bioarchaeological isotopic and genetic research is not for the purpose of multiple independent works with occasional conflict of interest. Cooperation between different scholars involved in this research is positively stimulated by the mutual need for the data of key importance for the research. On the one hand, bioarchaeology, and in particular archaeogenetics, commonly uses radiocarbon age determinations to build their models and infer from them, and on the other hand, as proven also by the current study, the assessment of the quality parameters of radiocarbon dating results is strongly dependent on the results of bioarchaeological studies.
Of course, whether one can expect to get results whose precision is adequate for the temporal scale of the analysed (pre)historic phenomena, and whether or not we interfere with any parallel research avenues, can be assessed only by knowing the advantages and limitations of the implementation of the proposed approach to radiocarbon dating. It is just at the operational level (starting with the sampling of skeletons and ending with modelling derived from their age determinations), where technological and analytical interdependencies and possible interference between the chronometric and bioarchaeological research can be pinpointed.
Firstly, it is necessary to, at the very least, take into account the possible negative consequences of destructive sampling in view of the possible other physical anthropology studies potentially performed. This problem arises even if the ‘stratified’ sampling of skeletons is carried out to such a limited extent as has been done here.
The preference of ribs, as an optimal reservoir of an ontogenetic deposit of collagen closely corresponding with the terminal life of individuals, is not determined only by chronometric factors, as alternatives could include vertebrae (cf. Fig. 4). The use of ribs is, however, better justified due to their large number and a relatively low diagnostic value in classical anthropological studies. Nevertheless, even the fragments of ribs intended for 14C dating should be chosen carefully, as there is a number of widely used and still developing methods of anthropological analysis based on these bone elements (cf. e.g. Dudar et al., 1993; İşcan et al., 1984; and Yoder et al., 2001).
The unavoidable destruction of the bony labyrinth raises a more serious methodological dilemma. Although also this skeletal element, in certain circumstances, appears to be much more important for anthropological analyses (cf. e.g. Lynnerup et al., 2006; Ponce de León et al., 2018; and Schutkowski, 1983), the problem goes far beyond the narrowly understood field of interest of the specialists involved in anthropology. It is not as much about the destroying of the structure of the organ itself, as about its bone tissue. Constituting a unique ‘biochemical archive’, it can yield a priceless material also for analyses of diet (see Jørkov et al., 2009), migration (see Harvig et al., 2014) and genetic studies (see Pinhasi et al., 2015).
The need to develop and maintain appropriate procedures in research programmes based on destructive analyses of (pre)historic human remains has recently been highlighted in the context of their perceived fairly unfettered use for genetic and isotopic analyses (Fox and Hawks, 2019). Currently, the growing awareness of the value of the petrous pyramid of the temporal bone for numerous and varied scientific studies has led to the elaboration of procedures for assessing the suitability of bony labyrinths for possible genetic and isotopic analyses (see Kontopoulos et al., 2019), as well as the devising of sampling techniques to maximise the variety of biochemical derivatives obtained from single bones (see Korlević et al., 2018). Nevertheless, being aware that such optimisation strategies cannot be always applied, we suggest that researchers intending to perform similar chronometric work to that reported in this study should, if possible, select skeletons with both temporal bones preserved and only analyse one of them.
Considering the interdisciplinary dependencies from the chronometric perspective, it should be stressed that careless use of the strategic bone fragments just for the purposes of radiocarbon dating would have negative implications also for the assessment of the 14C dates obtained. Narrowing our focus to the bioarchaeological premises that supported our reasoning, we would like to point to the exclusion of samples subject to the reservoir effect possibly resulting from the consumption of animals or plants from an aquatic environment. The possible inclusion in our analyses of radiocarbon dates based on the material with such a flaw would have distorted the results of the target modelling, as they depend on the exact determination of a temporal gap between two ontogenetically defined events. Due to the commonly known difficulty in the precise quantification of the freshwater reservoir offset (cf. e.g. Ascough et al., 2005; Dury et al., 2018; Keaveney and Reimer, 2012; and Philippsen and Heinemeier, 2013), all samples affected by this phenomenon would have to be excluded from chronometric analyses as indeterminable outliers (of the R-type, as defined by Bronk Ramsey, 2009b).
A harder issue to be faced with in the pursuit of radiocarbon dating with the accuracy of the destined chronological model is the relationship between human bones’ ontogenetic age and the age of the collagen they contain. It must therefore be stressed that the approach applied in the present work cannot be considered as an ideal answer to the problem. Following the study of Geyh (2001), we used the determination of the temporal lag between the radiocarbon age determinations obtained for the bones of deceased individuals in relation to the actual date of their death according to the developmental age of those individuals at the moment of death. However, at the time of that study, it was already suggested that ‘the specific 14C activity may not be uniformly distributed in all the bones in the body and may change along the growth direction’ (Geyh, 2001). It is those alleged differences in the isotopic composition of particular parts of the human skeleton (including the variation in 14C content) resulting from ontogenesis that was confirmed by the results of more recent studies (e.g. Fahy et al., 2017). Nevertheless, until now, anatomy-specific corrections have been provided only for femora (see Ubelaker et al., 2015). Future attempts at investigating radiocarbon lag in other bone tissues with different ontogenetic dynamics will surely improve the accuracy of 14C dates used to build sequences based on the ‘stratified’ sampling of human collagen.
The growing awareness of the ontogenetic differentiation of isotopic signatures of bone tissues within human skeleton not only makes the problem of their use for radiocarbon dating more sophisticated, but also reveals still unexplored potential of the advocated approach.
As could be easily noticed, the particular posterior probability distributions of the 14C dates modelled were to a large extent imposed by the shape of the corresponding calibration curve. However, this limited effect was mainly due to the simplicity of our binary sequences. It is well known that the final precision of age determinations obtained on the basis of similarly defined sequences strongly depends on the number of dates the basic chronometric models were built with (cf. e.g. Galimberti et al., 2004 and Goslar and Mądry, 1998). Therefore, although the Bayesian models based on the three results, i.e. the two 14C ages of the birth and one at death, complemented with the known lifespan of each individual, fulfilled our basic expectations, the creation of analogous sequences composed of 14C age determinations for definable intermediate ontological ‘strata’ of collagen is recommended.
In reference to the forensic studies that use radiocarbon dating to determine the date of birth and death on the basis of osteological remains, such extensive models shall be further developed by obtaining additional radiocarbon age determinations for teeth, especially molars (cf. Fig. 4). Of course, the use of successive bone tissues will complicate the sampling procedures and extend the range of bioarchaeological analyses associated with the dates. However, owing to the rapid development of genetic and isotopic research, most of these issues have already been explored (cf. e.g. Rose et al., 2018 and van der Sluis et al., 2015). Extending this to sequences of 14C dates based on the heterochronous collagen reservoirs in the human skeleton should therefore not pose major problems.
. Conclusions
This study explored the possibility of using a sequence of 14C dates from heterochronous ‘ontogenetic deposits’ of collagen within a single human skeleton in order to obtain more precise 14C dates for the moment of death and deposition of the body.
It revealed some potential as well as the most serious limitations for the implementation of the proposed radiocarbon dating strategy of (pre)historic human remains. Among the latter, a strong dependence of modelled radiocarbon dates’ precision on the shape of the corresponding calibration curve should be highlighted at first. For that reason, the precision of an individual’s radiocarbon age at the time of death in some cases can be raised only by several, whereas in others by several dozen per cent.
Also, the high costs of the multiple 14C dating and the necessity to combine these with multiproxy bioarchaeological analyses should be highlighted. Undoubtedly, this interdisciplinary entanglement additionally raises the expenses and difficulty of research. However, the agenda of modern bioarchaeological research on (pre)historic populations often comprises of isotopic and genetic investigations, which themselves commonly include 14C dates for the samples being analysed.
The implementation of the solution presented above should be relatively easy, especially since the petrous bone or teeth are biochemical archives commonly used for such bioarchaeological studies. Consequently, in order to obtain more precise dates in the manner presented here, it might be sufficient to add complementary radiocarbon age determinations from ribs or other ‘bone archives’ of ontogenetically close collagen to the research agenda.
Likewise, the broader application of the approach presented here promotes the continuous development of archaeological chronometric research significantly based on the burial (skeletal) finds. When attempting to refine chronologies, it is certainly worthwhile to consider the wider use of existing radiocarbon dates and the execution of complementary dates that could contribute to sequences similar to those created in this study. The inevitable consequence of such attempts is the coming together of chronometric studies with appropriate multifaceted bioarchaeological analyses.