
Leptin Is Associated with Poor Clinical Outcomes and Promotes Clear Cell Renal Cell Carcinoma Progression

 , , and
, , and
Abstract
:1. Introduction
2. Results
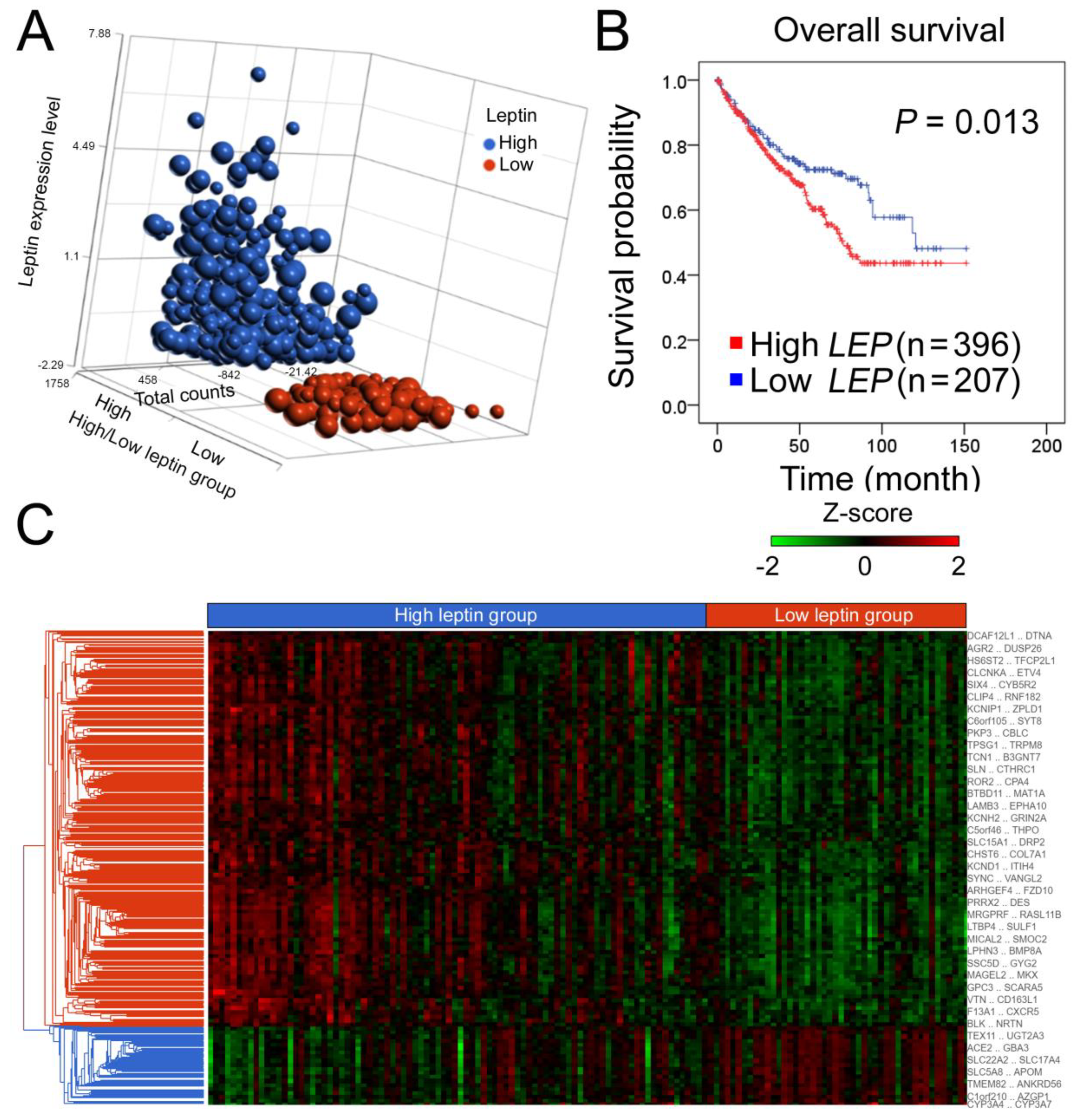
2.1. Leptin Is an Independent Predictor of a High Hazard Ratio for ccRCC, and the Hierarchical Clustering of Transcriptomic Data Is Shown Based on High and Low Leptin Groups in the ccRCC Cohort
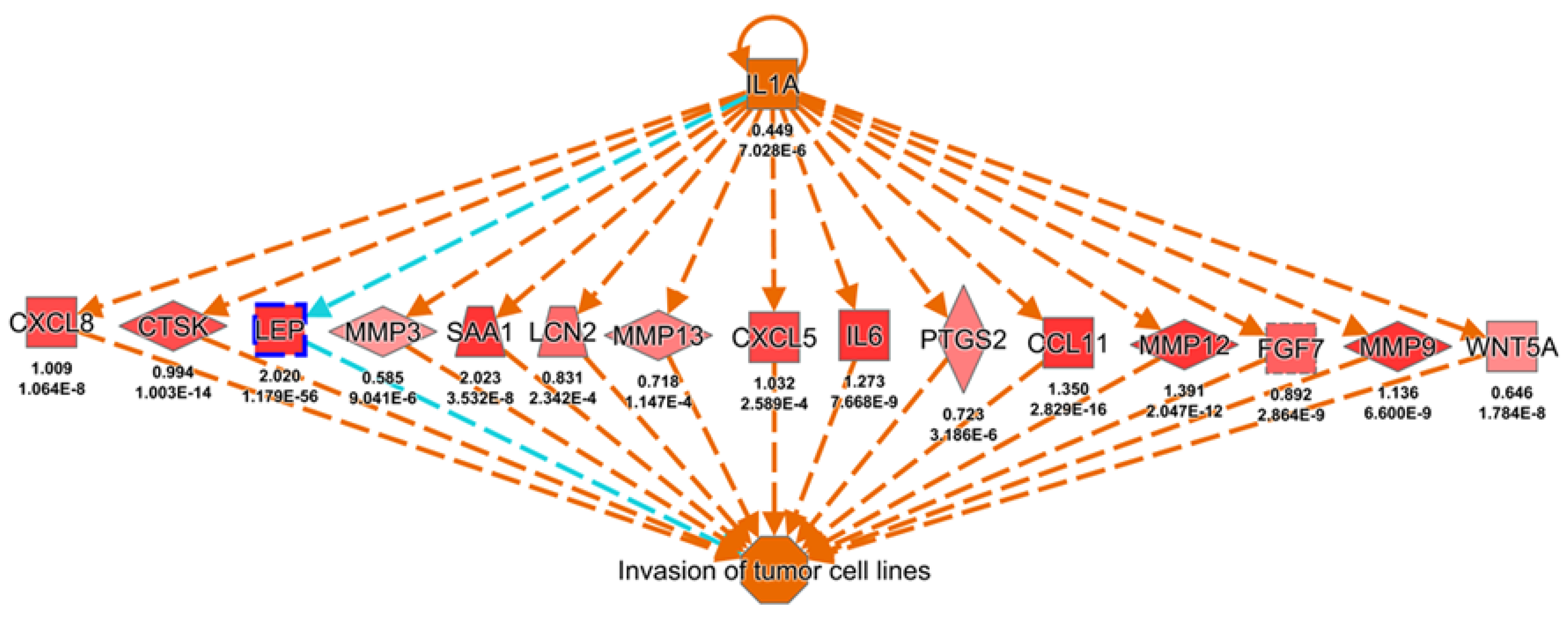
2.2. Knowledge-Based Transcriptomic Analysis Reveals Leptin’s Potential Effect in Promoting Cancer Metastasis
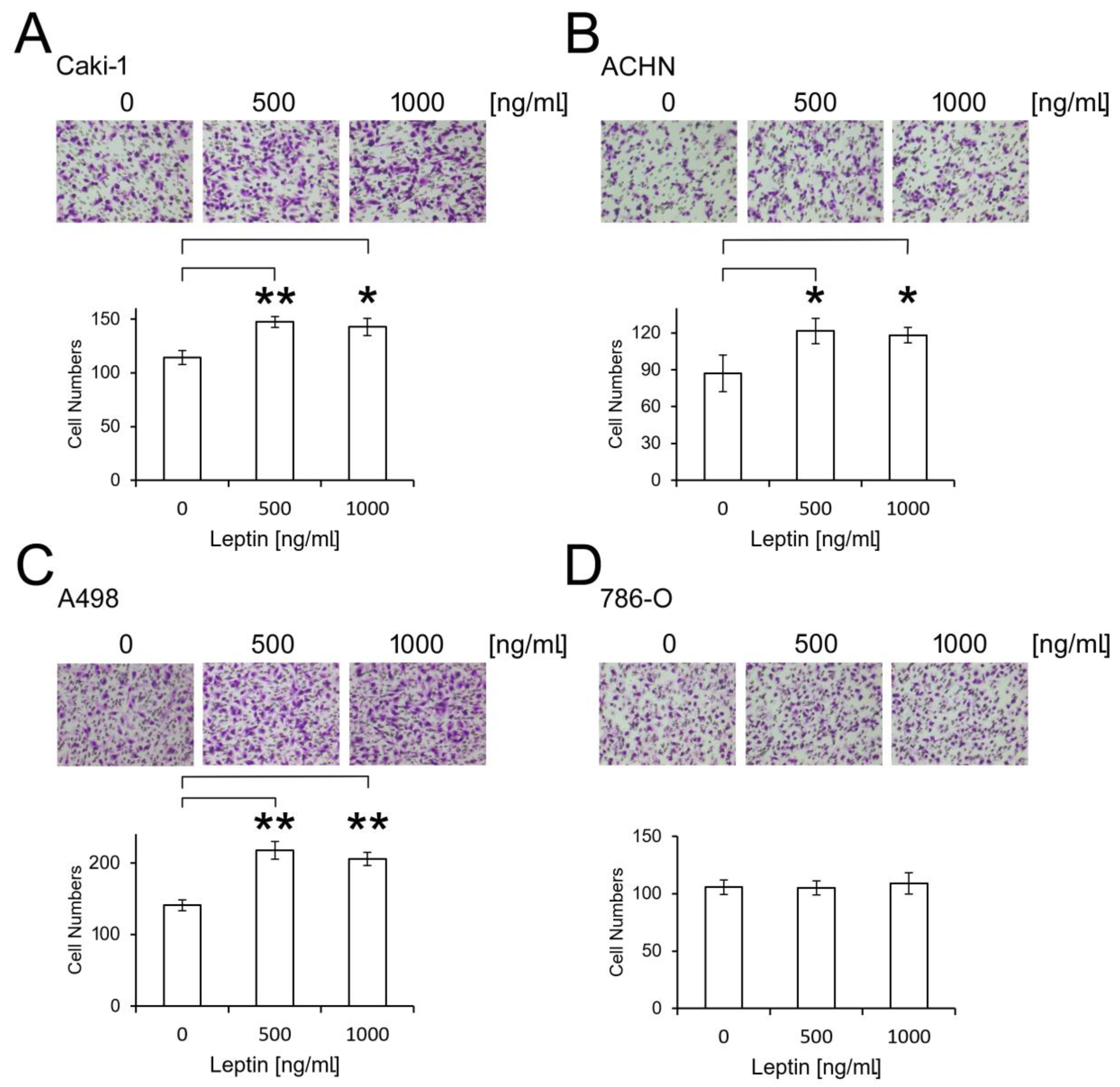
2.3. Leptin Promotes ccRCC Migration
2.4. The Planar Cell Polarity (PCP) Signaling Pathway Is Predictively Activated by Leptin
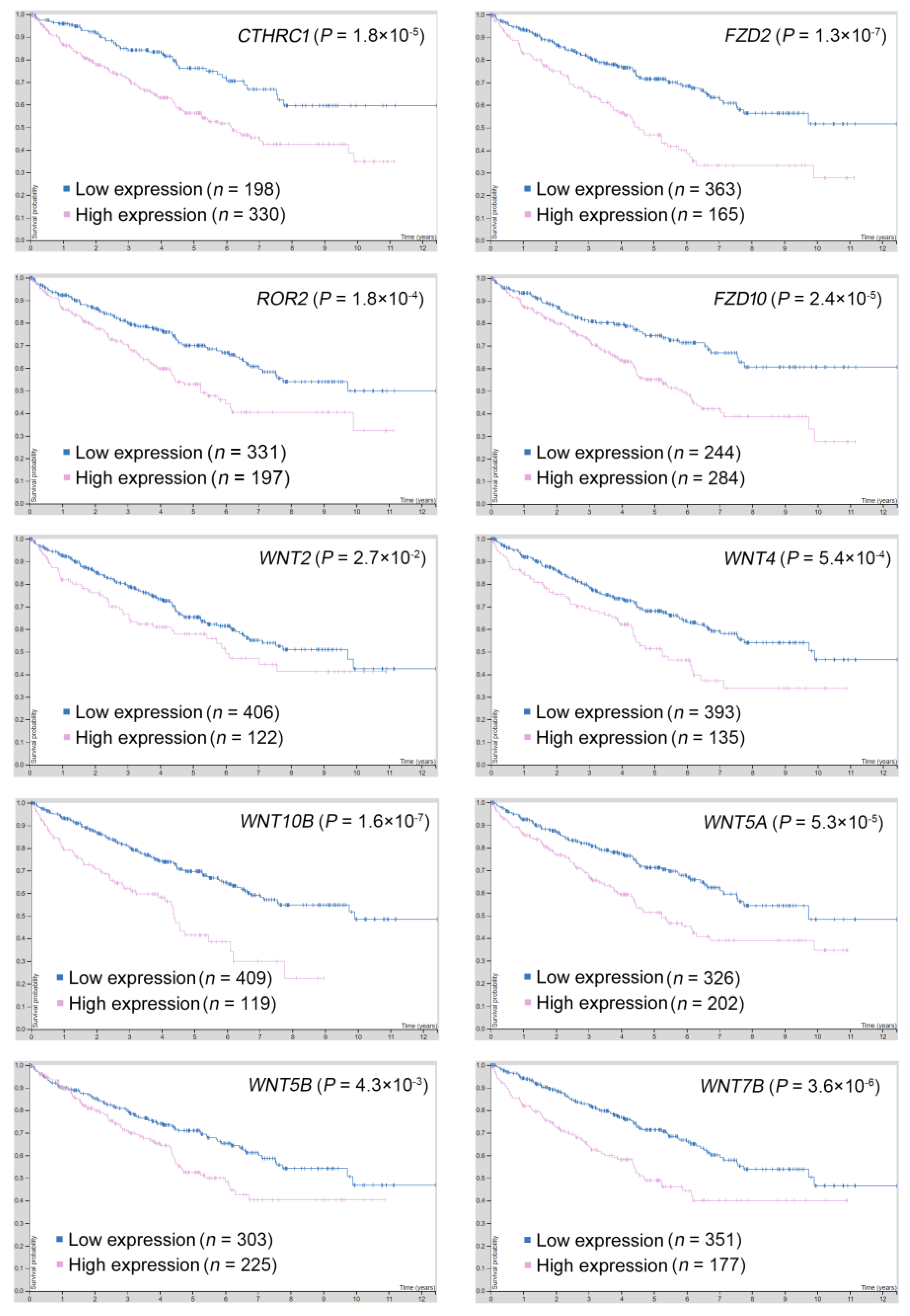
2.5. Gene Targets in the PCP Pathway Reveal Their Prognostic Significance in ccRCC
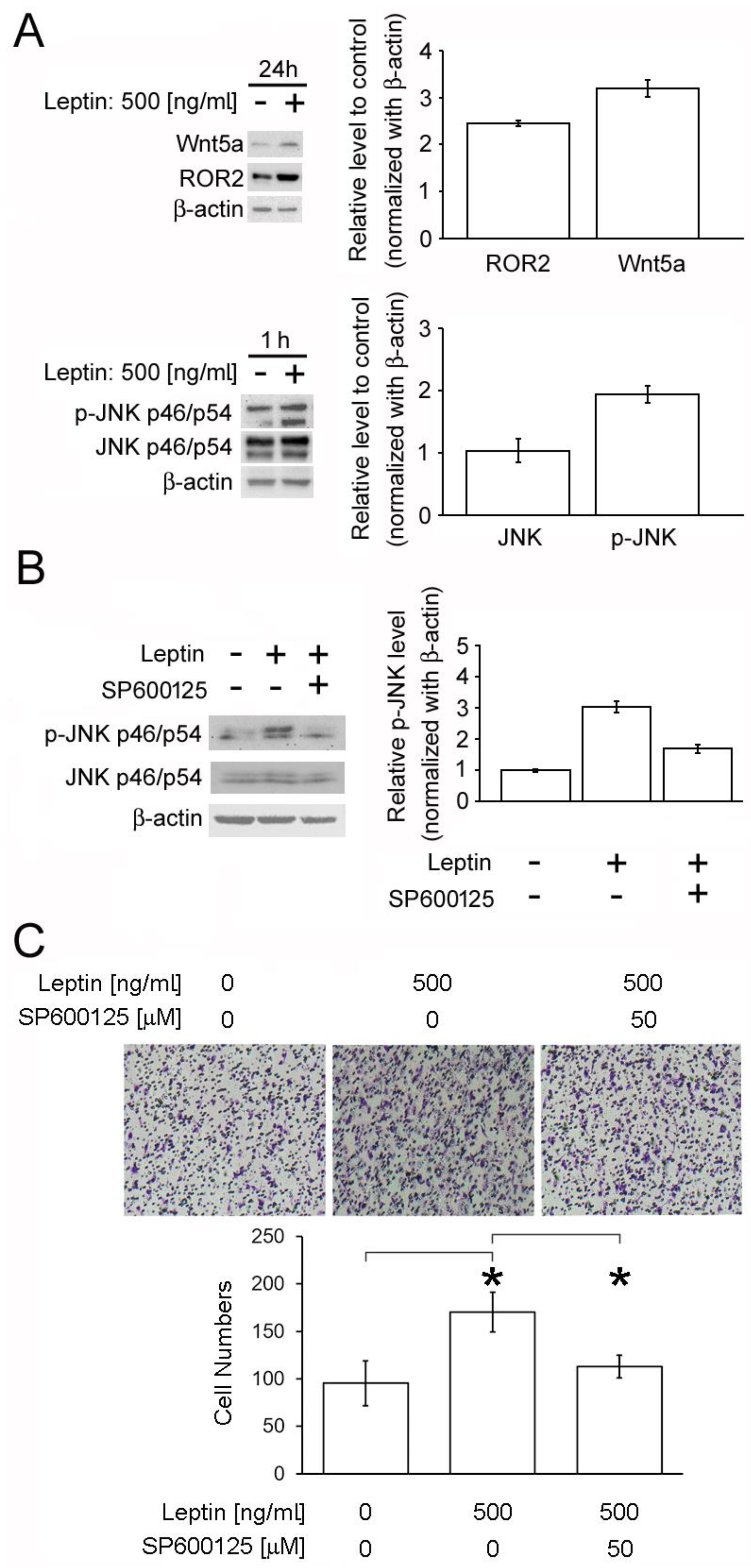
2.6. Leptin Triggers Cancer Cell Migration via the PCP/JNK Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. TCGA Dataset
4.2. Ingenuity Pathway Analysis (IPA)
4.3. Cell Culture
4.4. Cell Migration Assay
4.5. Western Blot Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, T.C.; Huang, K.W.; Liu, C.W.; Chang, Y.C.; Lin, W.M.; Yang, T.Y.; Hsiao, M. Leptin signaling axis specifically associates with clinical prognosis and is multifunctional in regulating cancer progression. Oncotarget 2018, 9, 17210–17219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nat. Cell Biol. 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Tartaglia, L.A.; Dembski, M.; Weng, X.; Deng, N.; Culpepper, J.; Devos, R.; Richards, G.J.; Campfield, L.; Clark, F.T.; Deeds, J.; et al. Identification and expression cloning of a leptin receptor, OB-R. Cell 1995, 83, 1263–1271. [Google Scholar] [CrossRef] [Green Version]
- Halaas, J.L.; Gajiwala, K.S.; Maffei, M.; Cohen, S.L.; Chait, B.T.; Rabinowitz, D.; Lallone, R.L.; Burley, S.K.; Friedman, J.M. Weight-reducing effects of the plasma protein encoded by the obese gene. Science 1995, 269, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Señarís, R.; Garcia-Caballero, T.; Casabiell, X.; Gallego, R.; Castro, R.; Considine, R.V.; Diéguez, C.; Casanueva, F.F. Synthesis of Leptin in Human Placenta. Endocrinololgy 1997, 138, 4501–4504. [Google Scholar] [CrossRef]
- Bado, A.; Levasseur, S.; Attoub, S.; Kermorgant, S.; Laigneau, J.P.; Bortoluzzi, M.N.; Moizo, L.; Lehy, T.; Guerre-Millo, M.; Le Marchand-Brustel, Y.; et al. The stomach is a source of leptin. Nat. Cell Biol. 1998, 394, 790–793. [Google Scholar] [CrossRef]
- Lin, T.C.; Lee, T.C.; Hsu, S.L.; Yang, C.S. The Molecular Mechanism of Leptin Secretion and Expression Induced by Aristolochic Acid in Kidney Fibroblast. PLoS ONE 2011, 6, e16654. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, R.; Hawkins, M.; Barzilai, N.; Rossetti, L. A nutrient-sensing pathway regulates leptin gene expression in muscle and fat. Nat. Cell Biol. 1998, 393, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Krabbe, L.M.; Bagrodia, A.; Margulis, V.; Wood, C.G. Surgical Management of Renal Cell Carcinoma. Semin. Interv. Radiol. 2014, 31, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Tosco, L.; Van Poppel, H.; Frea, B.; Gregoraci, G.; Joniau, S. Survival and Impact of Clinical Prognostic Factors in Surgically Treated Metastatic Renal Cell Carcinoma. Eur. Urol. 2013, 63, 646–652. [Google Scholar] [CrossRef]
- Teng, J.; Gao, Y.; Chen, M.; Wang, K.; Cui, X.; Liu, Y.; Xu, D. Prognostic value of clinical and pathological factors for surgically treated localized clear cell renal cell carcinoma. Chin. Med. J. 2014, 127, 1640–1644. [Google Scholar]
- Williamson, S.R.; Gupta, N.S.; Eble, J.N.; Rogers, C.G.; Michalowski, S.; Zhang, S.; Wang, M.; Grignon, D.J.; Cheng, L. Clear Cell Renal Cell Carcinoma With Borderline Features of Clear Cell Papillary Renal Cell Carcinoma. Am. J. Surg. Pathol. 2015, 39, 1502–1510. [Google Scholar] [CrossRef]
- Zhu, J.; Sanborn, J.Z.; Benz, S.; Szeto, C.; Hsu, F.; Kuhn, R.M.; Karolchik, D.; Archie, J.; Lenburg, M.E.; Esserman, L.J.; et al. The UCSC Cancer Genomics Browser. Nat. Methods 2009, 6, 239–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandervorst, K.; Dreyer, C.A.; Konopelski, S.E.; Lee, H.; Ho, H.Y.H.; Carraway, K.L. Wnt/PCP Signaling Contribution to Carcinoma Collective Cell Migration and Metastasis. Cancer Res. 2019, 79, 1719–1729. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Zhou, Q.; Liu, X.; Wang, C.; Liu, G. CTHRC1 overexpression promotes cervical carcinoma progression by activating the Wnt/PCP signaling pathway. Oncol. Rep. 2019, 41, 1531–1538. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Lu, H.; Lyu, Y.Y.; Yang, X.M.; Zhu, L.Y.; Yang, G.D.; Jiang, P.C.; Re, Y.; Song, W.W.; Wang, J.H.; et al. E6/E7-P53-POU2F1-CTHRC1 axis promotes cervical cancer metastasis and activates Wnt/PCP pathway. Sci. Rep. 2017, 7, 44744. [Google Scholar] [CrossRef] [Green Version]
- Luga, V.; Zhang, L.; Viloria-Petit, A.M.; Ogunjimi, A.A.; Inanlou, M.R.; Chiu, E.; Buchanan, M.; Hosein, A.N.; Basik, M.; Wrana, J.L. Exosomes Mediate Stromal Mobilization of Autocrine Wnt-PCP Signaling in Breast Cancer Cell Migration. Cell 2012, 151, 1542–1556. [Google Scholar] [CrossRef] [Green Version]
- Saling, M.; Duckett, J.K.; Ackers, I.; Coschigano, K.; Jenkinson, S.; Malgor, R. Wnt5a / planar cell polarity signaling pathway in urothelial carcinoma, a potential prognostic biomarker. Oncotarget 2017, 8, 31655–31665. [Google Scholar] [CrossRef]
- Martinez, S.; Scerbo, P.; Giordano, M.; Daulat, A.M.; Lhoumeau, A.-C.; Thomé, V.; Kodjabachian, L.; Borg, J.-P. The PTK7 and ROR2 Protein Receptors Interact in the Vertebrate WNT/Planar Cell Polarity (PCP) Pathway. J. Biol. Chem. 2015, 290, 30562–30572. [Google Scholar] [CrossRef] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Blal, H.A.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Uhlén, M.; Björling, E.; Agaton, C.; Szigyarto, C.A.K.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A Human Protein Atlas for Normal and Cancer Tissues Based on Antibody Proteomics. Mol. Cell. Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef] [Green Version]
- Tong, W.; Zeng, Y.; Chow, D.H.K.; Yeung, W.; Xu, J.; Deng, Y.; Chen, S.; Zhao, H.; Zhang, X.; Ho, K.K.; et al. Wnt16 attenuates osteoarthritis progression through a PCP/JNK-mTORC1-PTHrP cascade. Ann. Rheum. Dis. 2019, 78, 551–561. [Google Scholar] [CrossRef]
- Katoh, M. WNT/PCP signaling pathway and human cancer (Review). Oncol. Rep. 2005, 14, 1583–1588. [Google Scholar] [CrossRef]
- Crentsil, V.C.; Liu, H.; Sellitti, D.F. Comparison of exosomal microRNAs secreted by 786-O clear cell renal carcinoma cells and HK-2 proximal tubule-derived cells in culture identifies microRNA-205 as a potential biomarker of clear cell renal carcinoma. Oncol. Lett. 2018, 16, 1285–1290. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, A.; Hashemy, S.I.; Aghaei, M.; Panjehpour, M. Leptin induces matrix metalloproteinase 7 expression to promote ovarian cancer cell invasion by activating ERK and JNK pathways. J. Cell. Biochem. 2018, 119, 2333–2344. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Jin, S.W.; Kim, Y.A.; Khanal, T.; Lee, G.H.; Kim, S.J.; Rhee, S.D.; Chung, Y.C.; Hwang, Y.J.; Jeong, T.C.; et al. Leptin induces CREB-dependent aromatase activation through COX-2 expression in breast cancer cells. Food Chem. Toxicol. 2017, 106, 232–241. [Google Scholar] [CrossRef]
- Cao, D.; Luo, Y.; Qin, S.; Yu, M.; Mu, Y.; Ye, G.; Yang, N.; Cong, Z.; Chen, J.; Qin, J.; et al. Metallopanstimulin-1 (MPS-1) mediates the promotion effect of leptin on colorectal cancer through activation of JNK/c-Jun signaling pathway. Cell Death Dis. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunwobi, O.; Mutungi, G.; Beales, I.L.P. Leptin Stimulates Proliferation and Inhibits Apoptosis in Barrett’s Esophageal Adenocarcinoma Cells by Cyclooxygenase-2-Dependent, Prostaglandin-E2-Mediated Transactivation of the Epidermal Growth Factor Receptor and c-Jun NH2-Terminal Kinase Activation. Endocrinology 2006, 147, 4505–4516. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.L.; Guo, P.; Wang, J.; Liu, Q.Y.; Xu, J.F.; Yang, H.C.; Zhang, J.M. Association of the expression of leptin and leptin receptor with bone metastasis in pulmonary adenocarcinoma. Chin. J. Oncol. 2016, 38, 840–844. [Google Scholar]
- Oba, J.; Wei, W.; Gershenwald, J.E.; Johnson, M.M.; Wyatt, C.M.; Ellerhorst, J.A.; Grimm, E.A. Elevated Serum Leptin Levels are Associated with an Increased Risk of Sentinel Lymph Node Metastasis in Cutaneous Melanoma. Medicine 2016, 95, e3073. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Li, C.; Ai, H. Correlation analysis between the expressions of leptin and its receptor (ObR) and clinicopathology in endometrial cancer. Cancer Biomark. 2014, 14, 353–359. [Google Scholar] [CrossRef]
- Fan, Y.; Gan, Y.; Shen, Y.; Cai, X.; Song, Y.; Zhao, F.; Yao, M.; Gu, J.; Tu, H. Leptin signaling enhances cell invasion and promotes the metastasis of human pancreatic cancer via increasing MMP-13 production. Oncotarget 2015, 6, 16120–16134. [Google Scholar] [CrossRef] [Green Version]
- Perez-Hernandez, I.A.; Catalan, V.; Gomez-Ambrosi, J.; Rodriguez, A.; Fruhbeck, G. Mechanisms Linking Excess Adiposity and Carcinogenesis Promotion. Front. Endocrinol. 2014, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Pérez-Hernández, A.I.; Gurbindo, J.; Ramírez, B.; Méndez-Giménez, L.; Rotellar, F.; Valentí, V.; Moncada, R.; et al. Activation of Noncanonical Wnt Signaling Through WNT5A in Visceral Adipose Tissue of Obese Subjects Is Related to Inflammation. J. Clin. Endocrinol. Metab. 2014, 99, E1407–E1417. [Google Scholar] [CrossRef] [Green Version]
- Catalán, V.; Gómez-Ambrosi, J.; Rotellar, F.; Silva, C.; Rodríguez, A.; Salvador, J.; Gil, M.J.; Cienfuegos, J.A.; Frühbeck, G. Validation of Endogenous Control Genes in Human Adipose Tissue: Relevance to Obesity and Obesity-associated Type 2 Diabetes Mellitus. Horm. Metab. Res. 2007, 39, 495–500. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.-C. DDX3X is Epigenetically Repressed in Renal Cell Carcinoma and Serves as a Prognostic Indicator and Therapeutic Target in Cancer Progression. Int. J. Mol. Sci. 2020, 21, 2881. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Univariate | Multivariate | ||||
|---|---|---|---|---|---|
| Variable | Comparison | HR (95% CI) | p-Value | HR (95% CI) | p-Value |
| Sex | M:F | 0.939 (0.704–1.253) | 0.670 | 0.924 (0.607–1.404) | 0.924 |
| Stage | 3-4:1-2 | 3.817 (2.784–5.233) | <0.001 | 1.733 (0.685–4.385) | 0.246 |
| T | T3-4:T1-2 | 3.138 (2.320–4.245) | <0.001 | 1.370 (0.604–3.110) | 0.451 |
| N | N1:N0 | 3.380 (1.795–6.367) | <0.001 | 1.677 (0.860–3.268) | 0.129 |
| M | M1:M0 | 4.486 (3.372–5.967) | <0.001 | 2.458 (1.482–4.079) | <0.001 |
| Leptin | High:low | 1.479 (1.084–2.018) | 0.013 | 2.044 (1.224–3.416) | 0.006 |
| ID | Consistency Score | Regulators | Diseases and Functions |
|---|---|---|---|
| 1 | 3.615 | IL1A | Invasion of tumor cell lines |
| 2 | 3.5 | IL17A | Invasion of cells |
| 3 | 3.491 | PI3K (complex) | Migration of cells |
| 4 | 3.464 | EGFR | Size of body |
| 5 | 3.464 | cyclic AMP | Activation of cells |
| 6 | 3.411 | PI3K (complex) | Cell movement |
| 7 | 3.357 | Cigarette smoke | Cell proliferation of tumor cell lines |
| 8 | 3.333 | CCL11 | Migration of cells |
| 9 | 3.317 | Alpha catenin | Invasion of tumor cell lines |
| 10 | 3.317 | IL22 | Activation of cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, W.-L.; Yeh, Y.-M.; Liu, T.-T.; Lin, W.-M.; Yang, T.-Y.; Lee, C.-W.; Lin, T.-C. Leptin Is Associated with Poor Clinical Outcomes and Promotes Clear Cell Renal Cell Carcinoma Progression. Biomolecules 2021, 11, 431. https://doi.org/10.3390/biom11030431
Fan W-L, Yeh Y-M, Liu T-T, Lin W-M, Yang T-Y, Lee C-W, Lin T-C. Leptin Is Associated with Poor Clinical Outcomes and Promotes Clear Cell Renal Cell Carcinoma Progression. Biomolecules. 2021; 11(3):431. https://doi.org/10.3390/biom11030431
Chicago/Turabian StyleFan, Wen-Lang, Yuan-Ming Yeh, Tsung-Ta Liu, Wei-Ming Lin, Tse-Yen Yang, Chao-Wei Lee, and Tsung-Chieh Lin. 2021. "Leptin Is Associated with Poor Clinical Outcomes and Promotes Clear Cell Renal Cell Carcinoma Progression" Biomolecules 11, no. 3: 431. https://doi.org/10.3390/biom11030431