Biometry, Distribution and Genetic Characterization of Blue Jack Mackerel Trachurus picturatus (Bowdich, 1825), a Rare Pelagic Fish Species in the Adriatic Sea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Processing and Biometric Analysis
2.2. Genetic Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethical Statement
References
- Lynch, R. Globalisation and Official Statistics, ONS UK. In Proceedings of the Conference of European Statisticians, Paris, France, 13–15 June 2006. [Google Scholar]
- Smith-Vaniz, W.F. Carangidae. In Fishes of the North-Eastern Atlantic and the Mediterranean; Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; pp. 649–674, 1009–1031. [Google Scholar]
- Menezes, G.M.; Sigler, M.F.; Silva, H.M.; Pinho, M.R. Structure and zonation of demersal fish assemblages off the Azores Archipelago (mid-Atlantic). Mar. Ecol. Prog. Ser. 2006, 324, 241–260. [Google Scholar] [CrossRef] [Green Version]
- FAO. World Review of Highly Migratory Species and Straddling Stocks; FAO: Rome, Italy, 1994. [Google Scholar]
- Binni, G. Atlante dei Pesci delle Coste Italiane; Mondo Sommerso: Milano, Italy, 1968. [Google Scholar]
- Jardas, I. The Adriatic Ichthyofauna; Školska Knjiga d.d.: Zagreb, Croatia, 1996. [Google Scholar]
- Šoljan, T. I Pesci dell’Adriatico; A. Mondadori: Verona, Italy, 1975. [Google Scholar]
- Tortonese, E. Osteichtyes (Pesci ossei). Fauna d’Italia; XI Calderini: Bologna, Italy, 1975. [Google Scholar]
- Relini, G.; Lanteri, L. Osteichthyes. Biol. Mar. Mediterr. 2010, 17, 649–674. [Google Scholar]
- Županović, Š.; Jardas, I. Fauna i Flora Jadrana, Jabučka Kotlina, II; Logos: Split, Croatia, 1987. [Google Scholar]
- Bolognini, L.; Domenichetti, F.; Grati, F.; Polidori, P.; Scarcella, G.; Fabi, G. New record of the blue jack mackerel, Trachurus picturatus, T.E. Bowdich, 1825 (Osteichthyes: Carangidae) from the Northern Adriatic Sea. Acta Adriat. 2015, 56, 305–308. [Google Scholar]
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef]
- Žužul, I.; Šegvić-Bubić, T.; Talijančić, I.; Džoić, T.; Lepen Pleić, I.; Beg Paklar, G.; Ivatek-Šahdan, S.; Katavić, I.; Grubišić, L. Spatial connectivity pattern of expanding gilthead seabream populations and its interactions with aquaculture sites: A combined population genetic and physical modelling approach. Sci. Rep. 2019, 9, 14718. [Google Scholar] [CrossRef] [Green Version]
- Šegvić-Bubić, T.; Talijančić, I.; Grubišić, L.; Izquierdo-Gomez, D.; Katavić, I. Morphological and molecular differentiation of wild and farmed gilthead sea bream Sparus aurata: Implications for management. Aquac. Environ. Interact. 2014, 6, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Avignon, S.; Tastard, E.; Weston, S.; Duhamel, G.; Denis, F. Morphological identification and DNA barcoding used for diet analysis of gilthead seabream (Sparus aurata) in its expanding northerly range. Aquat. Living Resour. 2017, 30, 1. [Google Scholar] [CrossRef] [Green Version]
- Taggart, J.B.; Hynes, R.A.; Prodöuhl, P.A.; Terguson, A. A simplified protocol for routine total DNA isolation from salmonid fishes. J. Fish Biol. 1992, 40, 963–965. [Google Scholar] [CrossRef]
- Kocher, T.D.; Thomas, W.K.; Meyer, A.; Edwards, S.V.; Pääbo, S.; Villablanca, F.X.; Wilson, A.C. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA 1989, 86, 6196–6200. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Larget, B.; Simon, D.L. Markov chain Monte Carlo algorithms for the Bayesian analysis of phylogenetic trees. Mol. Biol. Evol. 1999, 16, 750–759. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. Figtree Version 1.4.4. 2018. Available online: https://github.com/rambaut/figtree/releases (accessed on 3 December 2020).
- Librado, P.; Rozas, J. DNASP V5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Res. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar]
- Ciampalini, A.; Forli, M.; Guerrini, A.; Sammartino, F. The marine fossils malacofauna in a Plio-Pleistocene section from Vallin Buio (Livorno, Italy). Biodivers. J. 2014, 5, 9–18. [Google Scholar]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- HIJRM. (Adriatic Sea. General Map of the Bottom Dediments) 1: 1 000 000; Jadransko More. Generalna Karta Sedimenta dna; Hidrografski Institut Jugoslavenske Ratne Mornarice: Split, Croatia, 1985. [Google Scholar]
- ICES. Report of the Working Group on Southern Horse Mackerel, Anchovy and Sardine (WGHANSA); ICES: Copenhagen, Denmark, 2018. [Google Scholar]
- Menezes, G.M.; Rosa, A.; Melo, O.; Pinho, M.R. Demersal fish assemblages off the Seine and Sedlo seamounts (Northeast Atlantic). Deep Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 2683–2704. [Google Scholar] [CrossRef]
- Jurado-Ruzafa, A.; Santamaria, M.T.G. Reproductive biology of the blue jack mackerel, Trachurus picturatus (Bowdich, 1825), off the Canary Islands. J. Appl. Ichthyol. 2013, 29, 526–531. [Google Scholar] [CrossRef]
- Bilge, G.; Yapıcı, S.; Filiz, H.; Cerim, H. Weight–length relations for 103 fish species from the southern Aegean Sea, Turkey. Acta Ichthyol. Piscat. 2014, 44, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.; Pereira, J.G.; Canha, Â.; Reis, D.; Diogo, H. Life history parameters of blue jack mackerel Trachurus picturatus (Teleostei: Carangidae) from north-east Atlantic. J. Mar. Biol. Assoc. UK 2015, 95, 401–410. [Google Scholar] [CrossRef]
- Jurado-Ruzafa, A.; Santamaria, M.T.G. Age, growth and natural mortality of blue jack mackerel Trachurus picturatus (Carangidae) from the Canary Islands, Spain (NW Africa). Afr. J. Mar. Sci. 2018, 40, 451–460. [Google Scholar] [CrossRef]
- Bektas, Y.; Belduz, A.O. Molecular phylogeny of Turkish Trachurus species (Perciformes: Carangidae) inferred from mitochondrial DNA analyses. J. Fish Biol. 2008, 73, 1228–1248. [Google Scholar] [CrossRef]
- Karaiskou, N.; Triantafyllidis, A.; Triantaphyllidis, C. Shallow genetic structure of three species of the genus Trachurus in European waters. Mar. Ecol. Prog. Ser. 2004, 281, 193–205. [Google Scholar] [CrossRef]
- Karaiskou, N.; Apostolidis, A.P.; Triantafyllidis, A.; Kouvatsi, A.; Triantaphyllidis, C. Genetic identification and phylogeny of three species of the genus Trachurus based on mitochondrial DNA analysis. Mar. Biotechnol. 2003, 5, 493–504. [Google Scholar]
- Moreira, C.; Correia, A.T.; Vaz-Pires, P.; Froufe, E. Genetic diversity and population structure of the blue jack mackerel Trachurus picturatus across its western distribution. J. Fish Biol. 2019, 94, 725–731. [Google Scholar] [CrossRef]
- Tuset, V.M.; Jurado-Ruzafa, A.; Otero-Ferrer, J.L.; Santamaría, M.T.G. Otolith phenotypic variability of the blue jack mackerel, Trachurus picturatus, from the Canary Islands (NE Atlantic): Implications in its population dynamic. Fish. Res. 2019, 218, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, J.; Vieira, A.R.; Sequeira, V.; González, J.A.; Kaufmann, M.; Gordo, L.S. Identifying populations of the blue jack mackerel (Trachurus picturatus) in the Northeast Atlantic by using geometric morphometrics and otolith shape analysis. Fish. Bull. 2018, 116, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Moreira, C.; Presa, P.; Correia, A.T.; Vaz-Pires, P.; Froufe, E. Spatio-temporal microsatellite data suggest a multidirectional connectivity pattern in the Trachurus picturatus metapopulation from the Northeast Atlantic. Fish. Res. 2020, 225, 105499. [Google Scholar] [CrossRef]
- Tanner, S.E.; Pérez, M.; Presa, P.; Thorrold, S.R.; Cabral, H.N. Integrating microsatellite DNA markers and otolith geochemistry to assess population structure of European hake (Merluccius merluccius). Estuar. Coast. Shelf Sci. 2014, 142, 68–75. [Google Scholar] [CrossRef]
- Grbec, B.; Morović, M.; Matić, F.; Ninčević Gladan, Ž.; Marasović, I.; Vidjak, O.; Bojanić, N.; Čikeš Keč, V.; Zorica, B.; Kušpilić, G.; et al. Climate regime shifts and multi-decadal variability of the Adriatic Sea pelagic ecosystem. Acta Adriat. 2014, 55, 117–126. [Google Scholar]





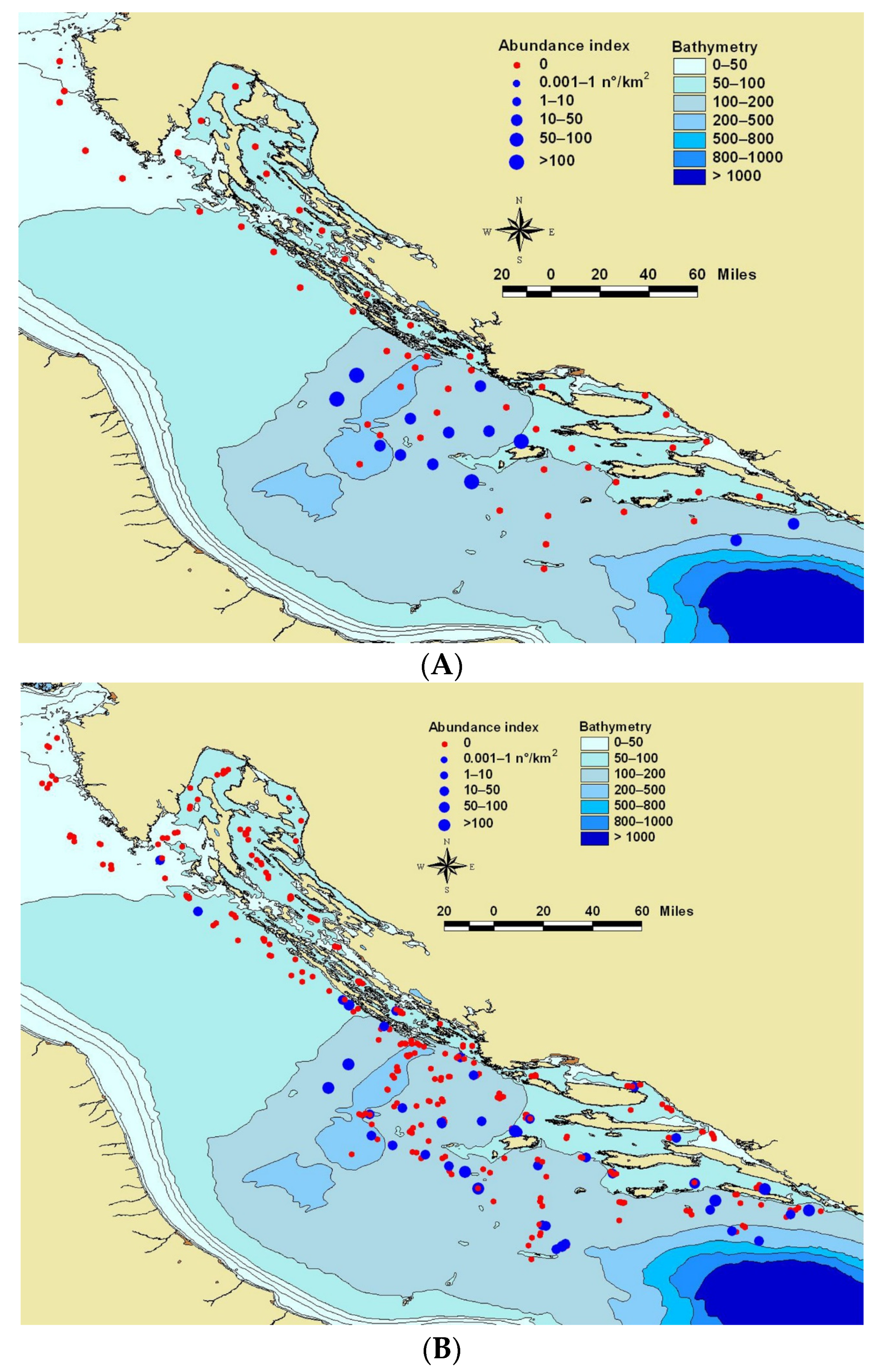{kind=link}
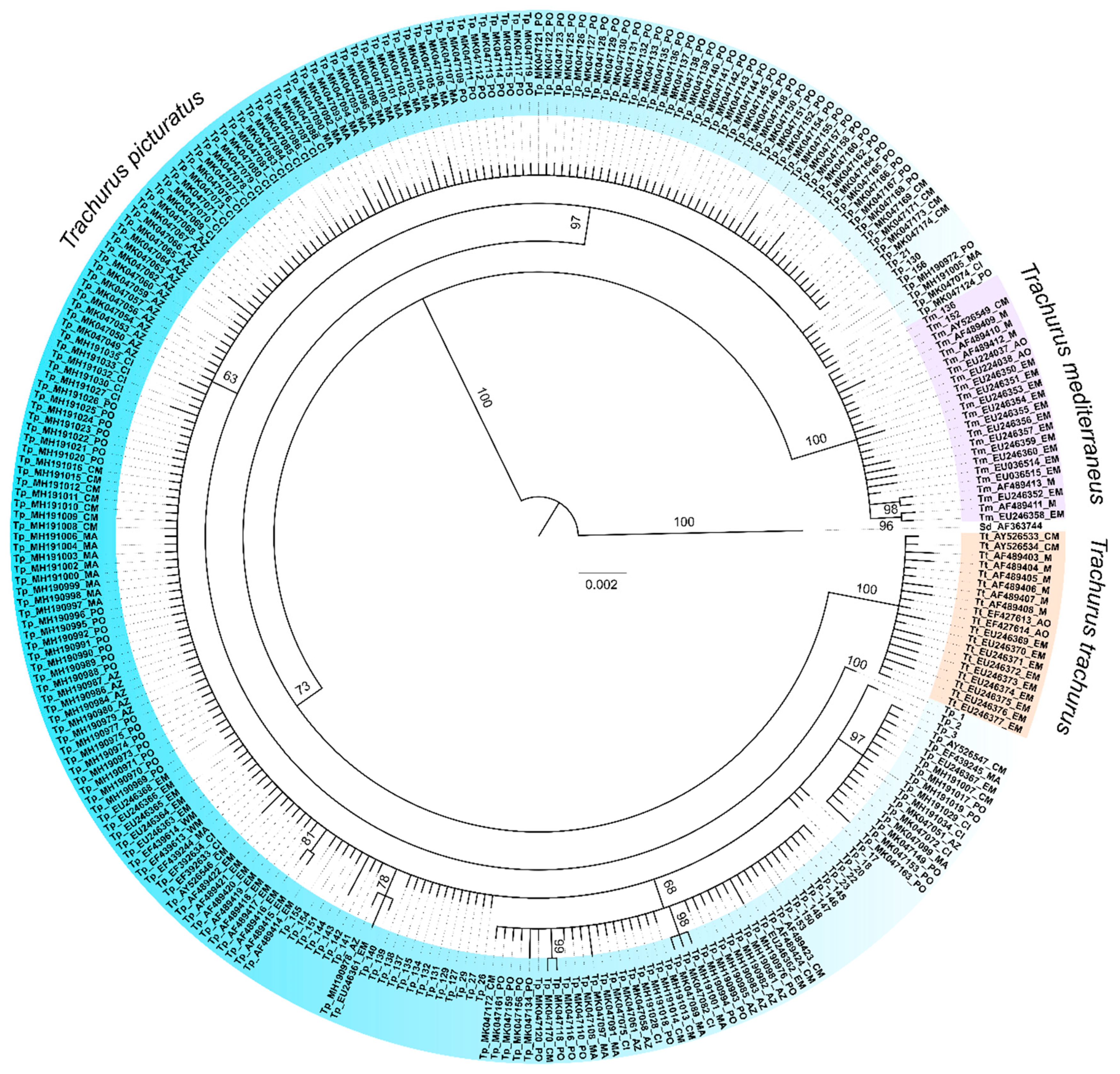{kind=link}
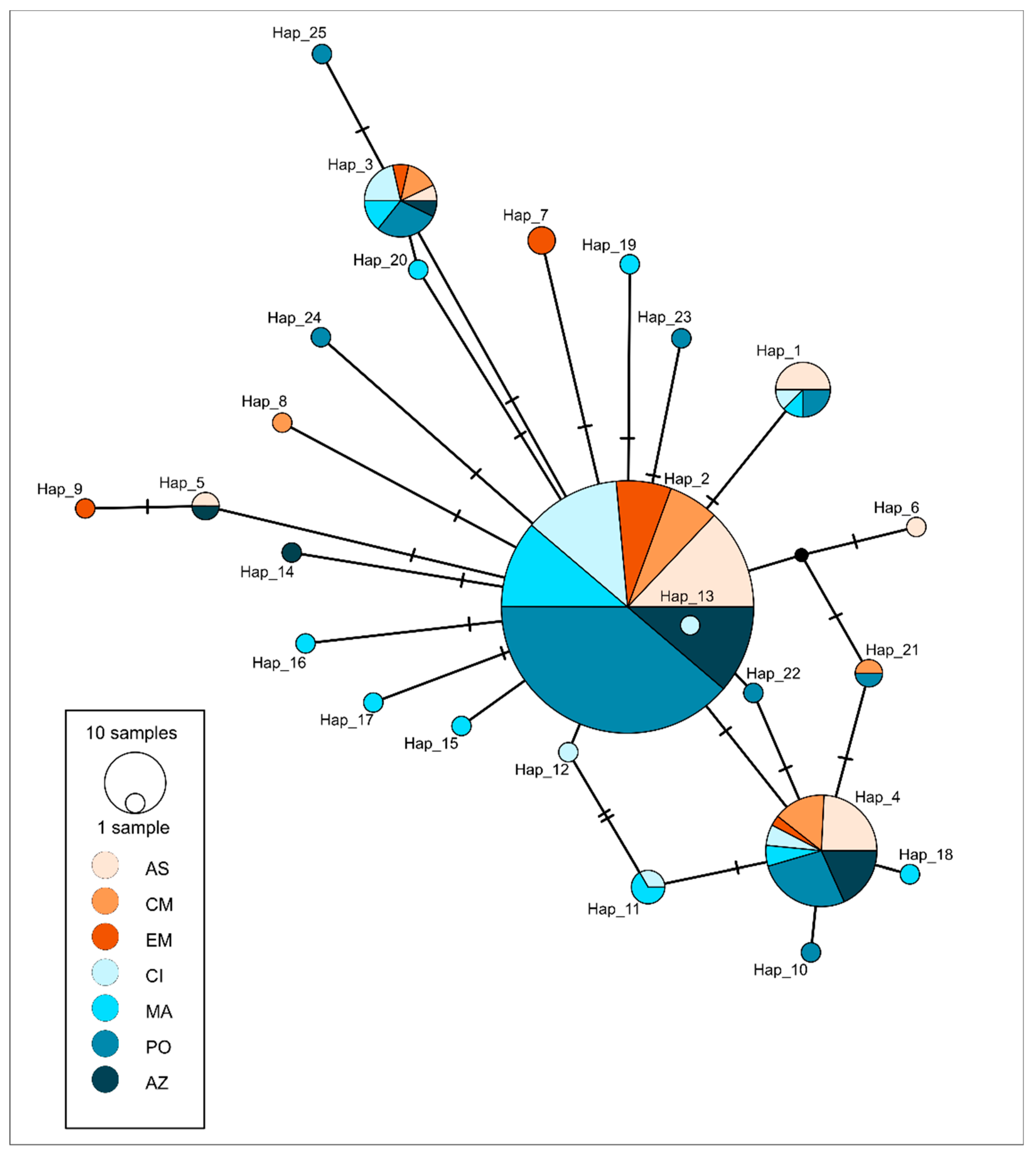{kind=link}
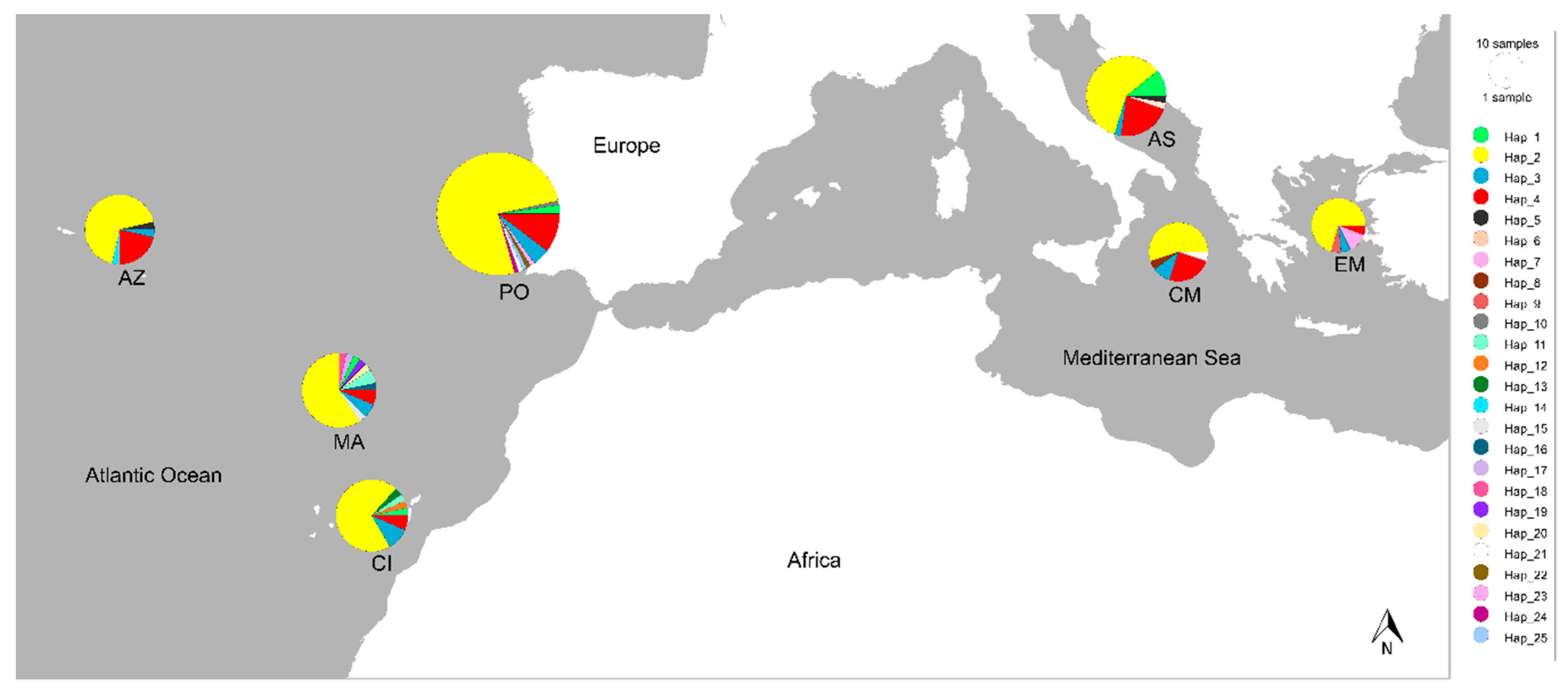{kind=link}
| Morphometric | |||||
| Relation | N | Range (%) | (%) | (%) | CV (%) |
| FL/TL | 155 | 79.9–98.3 | 91.3 ± 2.0 | 91.3 ± 0.1 | 1.42 |
| SL/TL | 155 | 75.0–95.7 | 86.8 ± 2.5 | 86.8 ± 0.1 | 1.59 |
| PAL/TL | 155 | 40.3–63.7 | 55.3 ± 17.7 | 55.3 ± 0.3 | 4.21 |
| LH/TL | 155 | 15.3–32.8 | 24.1 ± 2.8 | 24.1 ± 0.1 | 1.67 |
| O/LH | 155 | 17.9–38.9 | 24.8 ± 5.9 | 24.8 ± 0.2 | 2.43 |
| H/TL | 155 | 13.9–23.5 | 17.8 ± 1.4 | 17.8 ± 0.1 | 1.16 |
| LD1/TL | 155 | 11.8–20.0 | 14.3 ± 1.3 | 14.3 ± 0.1 | 1.14 |
| LD2/TL | 155 | 29.2–41.5 | 35.5 ± 2.4 | 35.5 ± 0.1 | 1.56 |
| LP/TL | 155 | 16.5–26.4 | 19.6 ± 2.0 | 19.6 ± 0.1 | 1.43 |
| Meristic | |||||
| Characteristic | N | Range | CV | ||
| 1st dorsal fin rays (D1) | 59 | 6–9 | 7.8 ± 0.58 | 7.8 ± 0.07 | 7.39 |
| 2nd dorsal fin rays (D2) | 59 | 26–36 | 32.0 ± 2.33 | 32.0 ± 0.30 | 7.31 |
| Pectoral fin rays (P) | 59 | 17–23 | 19.9 ± 1.50 | 19.9 ± 0.19 | 7.53 |
| Anal fin rays (A) | 59 | 26–34 | 30.9 ± 1.78 | 30.9 ± 0.23 | 5.78 |
| Ventral fin rays (V) | 59 | 5 | 5 ± 0.00 | 5.0 ± 0.00 | 0 |
| Equation | a | b | r2 |
|---|---|---|---|
| FL = a + bTL | 0.29 | 0.89 | 0.995 |
| SL = a + bTL | −0.05 | 0.87 | 0.994 |
| PAL = a + bTL | 1.8 | 0.4 | 0.852 |
| LH = a + bTL | −0.11 | 0.25 | 0.934 |
| O = a + bLH | −0.02 | 0.25 | 0.883 |
| H = a + bTL | −0.22 | 0.2 | 0.943 |
| LD1 = a + bTL | 0.05 | 0.14 | 0.877 |
| LD2 = a + bTL | −0.10 | 0.36 | 0.967 |
| LP = a + bTL | −0.85 | 0.27 | 0.957 |
| Sampling Locality | N | h (±SD) | π (±SD) | S | Tajima’s D | Fu’s FS |
|---|---|---|---|---|---|---|
| Adriatic Sea | 37 | 0.602 ± 0.075 | 0.0038 ± 0.0007 | 6 | −1.274 (p = 0.10) | −2.217 (p = 0.06) |
| Eastern Mediterranean Sea | 17 | 0.945 ± 0.027 | 0.0085 ± 0.0016 | 5 | −1.719 (p = 0.02) | −2.308 (p = 0.01) |
| Central Mediterranean Sea | 20 | 0.652 ± 0.093 | 0.0042 ± 0.0009 | 4 | −0.760 (p = 0.26) | −1.592 (p = 0.08) |
| Portugal mainland | 87 | 0.415 ± 0.065 | 0.0027 ± 0.0005 | 9 | −1.788 (p = 0.01) | −8.135 (p < 0.001) |
| Canary Islands | 30 | 0.508 ± 0.108 | 0.0032 ± 0.0008 | 5 | −1.352 (p = 0.07) | −4.511 (p < 0.001) |
| Madeira | 32 | 0.649 ± 0.096 | 0.0047 ± 0.0010 | 9 | −1.764 (p = 0.02) | −8.764 (p < 0.001) |
| Azores | 28 | 0.131 ± 0.082 | 0.0004 ± 0.0003 | 4 | −1.182 (p = 0.11) | −2.239 (p = 0.04) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zorica, B.; Bušelić, I.; Čikeš Keč, V.; Vuletin, V.; Pleić, I.L.; Isajlović, I.; Radonić, I.; Vrgoč, N. Biometry, Distribution and Genetic Characterization of Blue Jack Mackerel Trachurus picturatus (Bowdich, 1825), a Rare Pelagic Fish Species in the Adriatic Sea. Diversity 2020, 12, 463. https://doi.org/10.3390/d12120463
Zorica B, Bušelić I, Čikeš Keč V, Vuletin V, Pleić IL, Isajlović I, Radonić I, Vrgoč N. Biometry, Distribution and Genetic Characterization of Blue Jack Mackerel Trachurus picturatus (Bowdich, 1825), a Rare Pelagic Fish Species in the Adriatic Sea. Diversity. 2020; 12(12):463. https://doi.org/10.3390/d12120463
Chicago/Turabian StyleZorica, Barbara, Ivana Bušelić, Vanja Čikeš Keč, Vedran Vuletin, Ivana Lepen Pleić, Igor Isajlović, Ivana Radonić, and Nedo Vrgoč. 2020. "Biometry, Distribution and Genetic Characterization of Blue Jack Mackerel Trachurus picturatus (Bowdich, 1825), a Rare Pelagic Fish Species in the Adriatic Sea" Diversity 12, no. 12: 463. https://doi.org/10.3390/d12120463