Genetic Distinctiveness Highlights the Conservation Value of a Sicilian Manna Ash Germplasm Collection Assigned to Fraxinus angustifolia (Oleaceae)

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Morphological Trait Variation
2.3. Genome Size and Ploidy Level
2.4. DNA Extraction and Genotyping
2.5. Data Analysis
3. Results
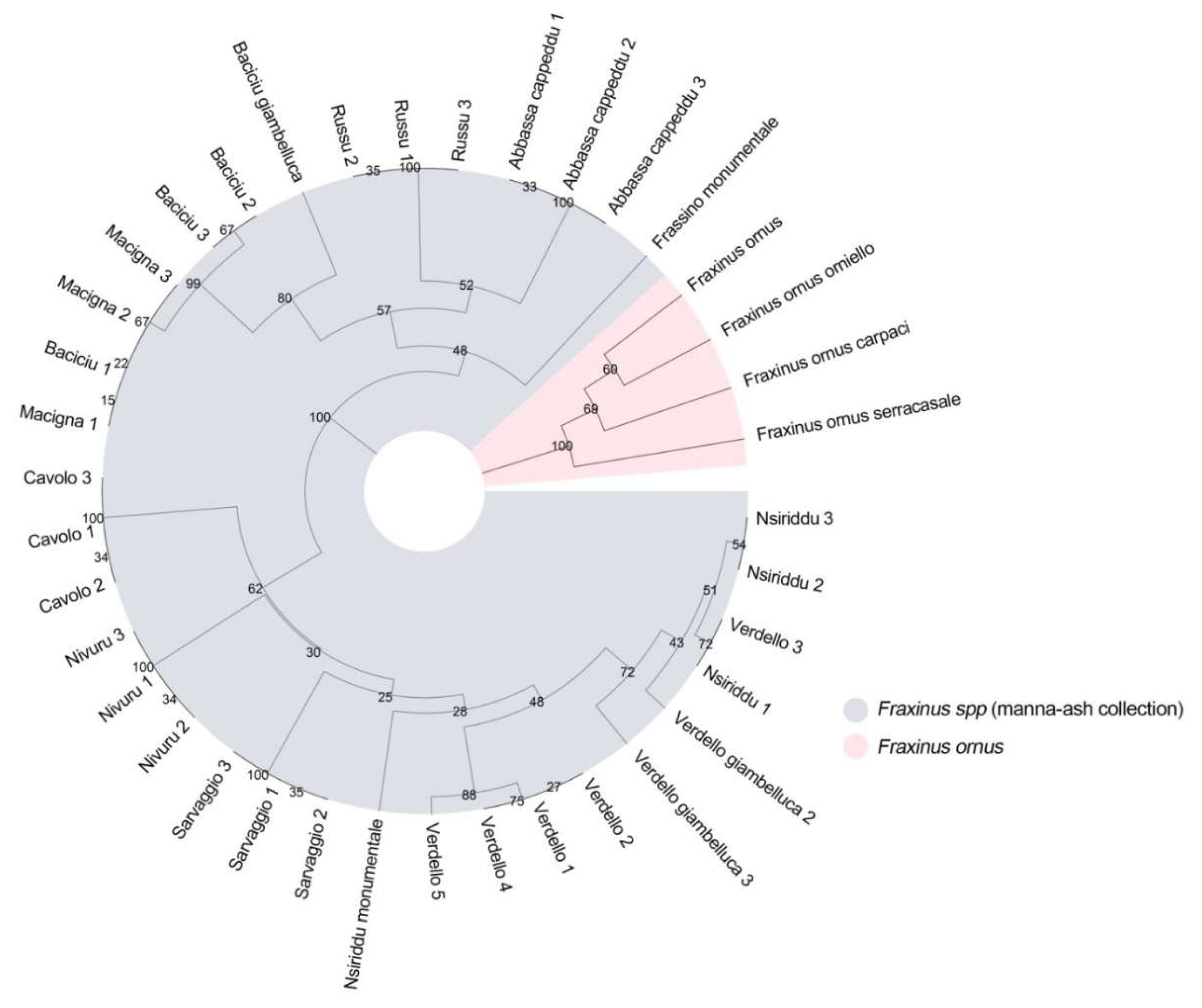
3.1. Genetic Variation of Plastid DNA Microsatellites (cpSSRs) and Amplified Regions in the Manna Ash Collection
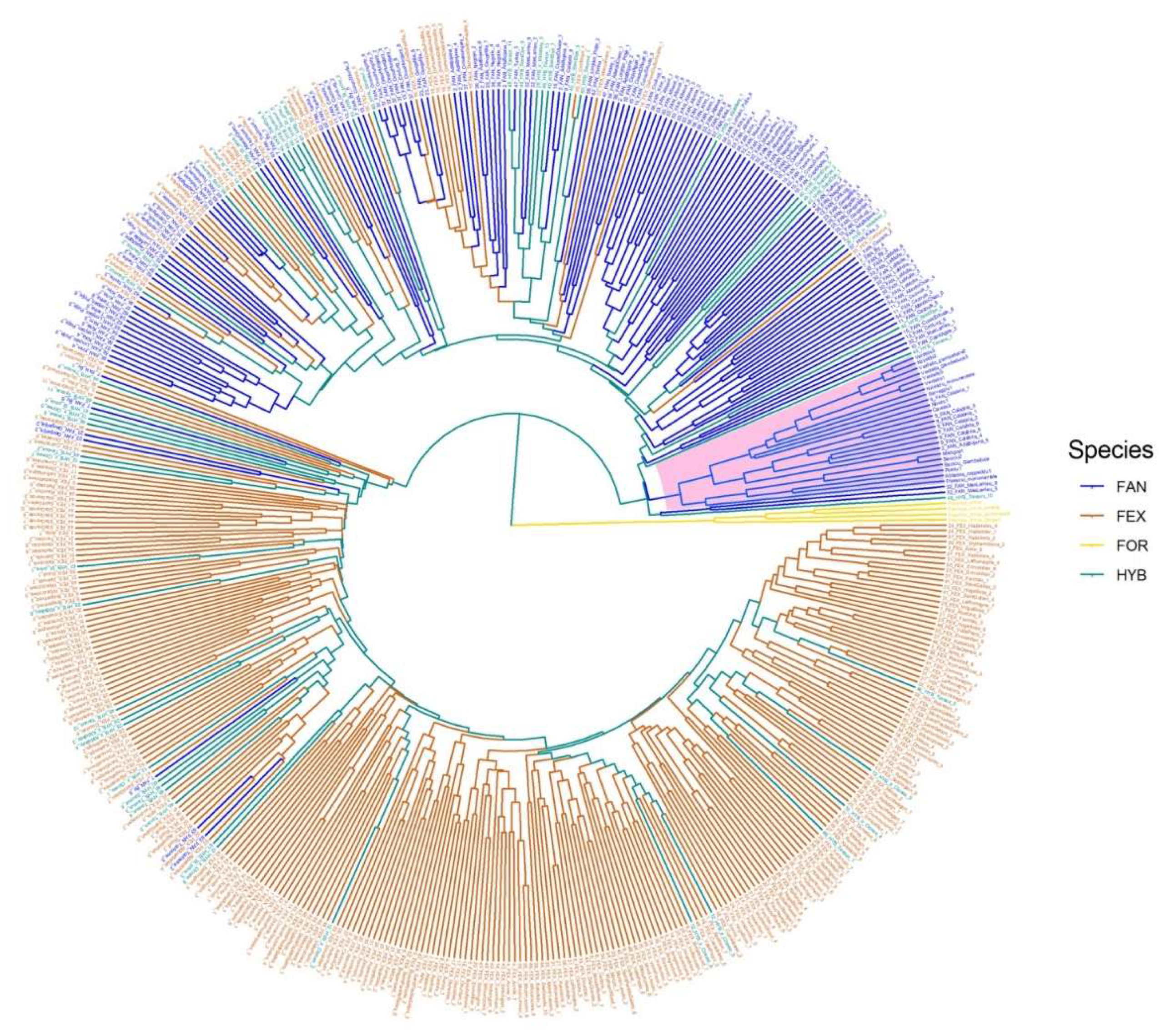
3.2. Genetic Diversity of Manna Ash Germplasm
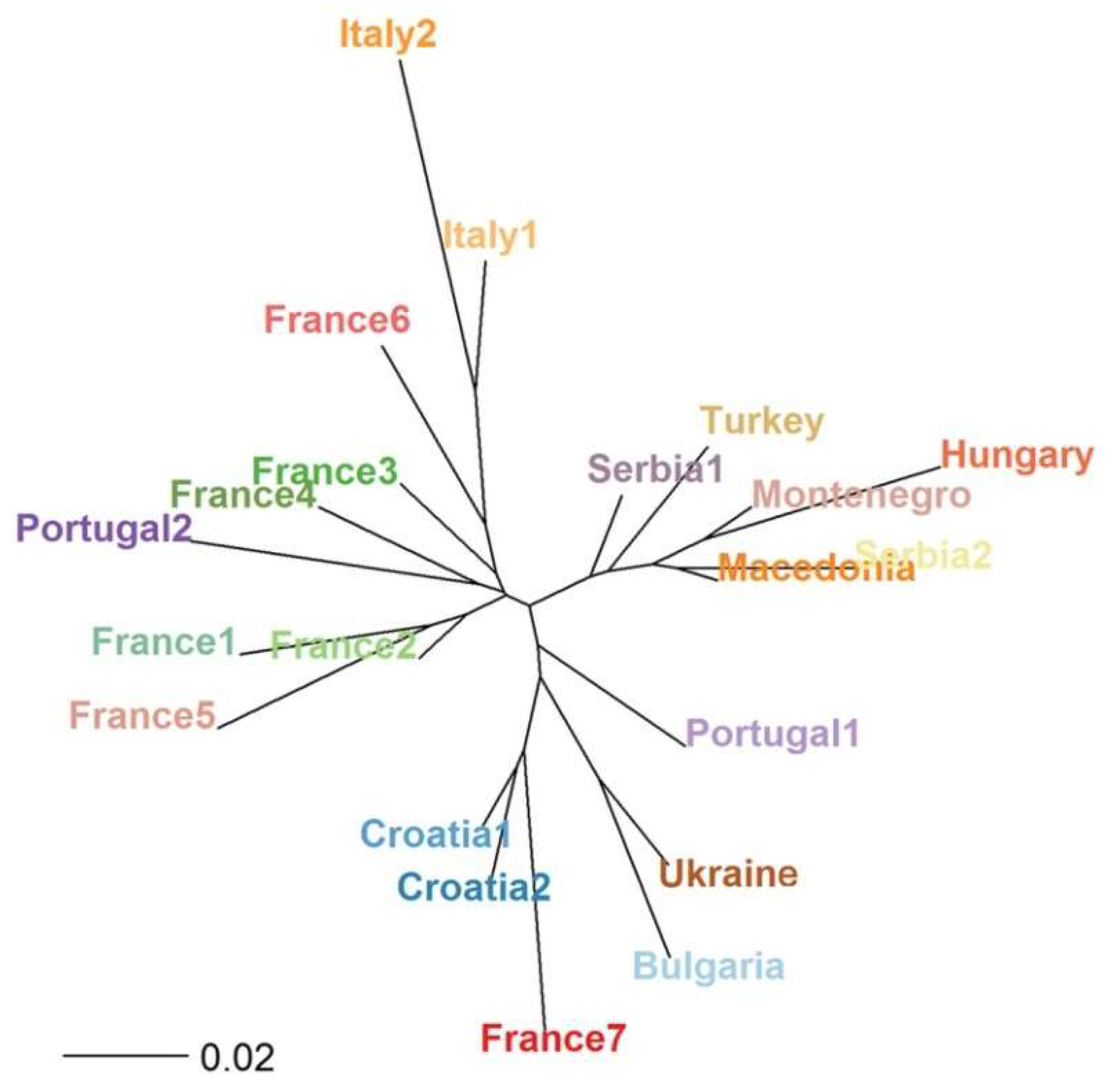
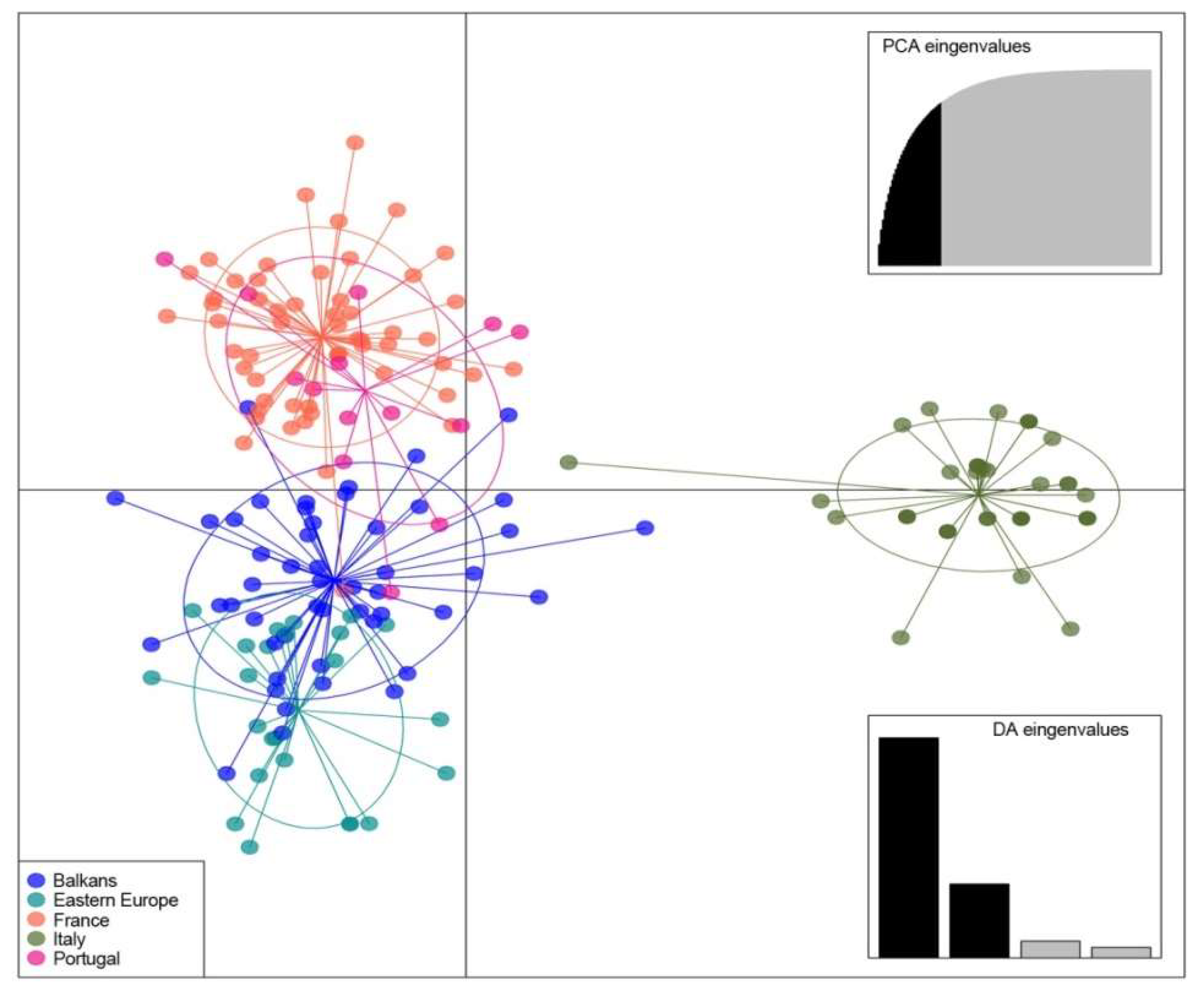
3.3. Genetic Diversity of Fraxinus spp. in Europe
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wallander, E.; Albert, V.A. Phylogeny and classification of Oleaceae based on rps16 and trnL-F sequence data. Am. J. Bot. 2000, 87, 1827–1841. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, D.D.; Gaudeul, M.; Couloux, A.; Bousquet, J.; Frascaria-Lacoste, N. The phylogeography of Eurasian Fraxinus species reveals ancient transcontinental reticulation. Mol. Phylogenet. Evol. 2014, 77, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Wallander, E. Systematics of Fraxinus (Oleaceae) and evolution of dioecy. Plant Syst. Evol. 2008, 273, 25–49. [Google Scholar] [CrossRef]
- Heuertz, M.; Carnevale, S.; Fineschi, S.; Sebastiani, F.; Hausman, J.F.; Paule, L.; Vendramin, G.G. Chloroplast DNA phylogeography of European ashes, Fraxinus sp. (Oleaceae): Roles of hybridization and life history traits. Mol. Ecol. 2006, 15, 2131–2140. [Google Scholar] [CrossRef] [PubMed]
- Picard, J.-F. Contribution à l’étude de la biologie florale et de la fructification du Frêne commun (Fraxinus excelsior L.). Rev. For. Fr. 1982, 97. [Google Scholar] [CrossRef] [Green Version]
- Wallander, E. Evolution of Wind-Pollination in Fraxinus (Oleaceae): An Ecophylogenetic Approach. Ph.D. Thesis, Göteborg University, Göteborg, Sweden, 2001. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.E.; Chater, A.O.; Defilipps, R.A.; Richardson, I.B.K.; et al. Flora Europaea: Plantaginaceae to Compositae (and Rubiaceae); Cambridge University Press: Cambridge, UK, 1972. [Google Scholar]
- Huntley, B.; Birks, H.J.B.; Harry, J.B. Atlas of Past and Present Pollen Maps for Europe, 0–13,000 Years Ago; Cambridge University Press: Cambridge, UK, 1983; ISBN 978-0-521-23735-2. [Google Scholar]
- Temunović, M.; Franjić, J.; Satovic, Z.; Grgurev, M.; Frascaria-Lacoste, N.; Fernández-Manjarrés, J.F. Environmental Heterogeneity Explains the Genetic Structure of Continental and Mediterranean Populations of Fraxinus angustifolia Vahl. PLoS ONE 2012, 7, e42764. [Google Scholar] [CrossRef] [Green Version]
- FRAXIGEN, 2005 Ash Species in Europe: Biological Characteristics and Practical Guidelines for Sustainable Use—ProQuest. Available online: https://search.proquest.com/openview/e333985a9ac22793988179029545b7da/1?pq-origsite=gscholarandcbl=29663 (accessed on 17 June 2020).
- Thomas, P.A. Biological Flora of the British Isles: Fraxinus excelsior. J. Ecol. 2016, 104, 1158–1209. [Google Scholar] [CrossRef]
- Caudullo, G.; Houston Durrant, T. Fraxinus angustifolia in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species 2016; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; p. 97. [Google Scholar]
- Bogdan, S.; Kajba, D.; Franjic, J.; Idzojtic, M.; Skvorc, Z.; Trupčević, I.K. Genetic variation in quantitative traits within and among Croatian narrow-leaved ash (Fraxinus angustifolia Vahl) populations assessed in an open-pollinated progeny test. Period. Boil. 2007, 109, 1–11. [Google Scholar]
- Gérard, P.R.; Temunović, M.; Sannier, J.; Bertolino, P.; Dufour, J.; Frascaria-Lacoste, N.; Fernández-Manjarrés, J.F. Chilled but not frosty: Understanding the role of climate in the hybridization between the Mediterranean Fraxinus angustifolia Vahl and the temperate Fraxinus excelsior L. (Oleaceae) ash trees. J. Biogeogr. 2013, 40, 835–846. [Google Scholar] [CrossRef]
- Noti, R.; van Leeuwen, J.F.N.; Colombaroli, D.; Vescovi, E.; Pasta, S.; La Mantia, T.; Tinner, W. Mid- and late-Holocene vegetation and fire history at Biviere di Gela, a coastal lake in southern Sicily, Italy. Veg. Hist. Archaeobot. 2009, 18, 371–387. [Google Scholar] [CrossRef] [Green Version]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Mazzola, P.; Schicchi, R.; Spadaro, V. Ricerche finalizzate al recupero e conservazione del germoplasma dei frassini da manna sulle Madonie (Sicilia settentrionale). In Proceedings of the 101 Congresso Società Botanica Italiana, Caserta, Italy, 27–29 September 2006; p. 137. [Google Scholar]
- Schicchi, R.; Camarda, L.; Spadaro, V.; Pitonzo, R. Caratterizzazione chimica della manna estratta nelle Madonie (Sicilia) da cultivar di Fraxinus angustifolia e di Fraxinus ornus (Oleaceae). Quad. Bot. Amb. Appl. 2006, 17, 151–174. [Google Scholar]
- Giardinieri, A.; Schicchi, R.; Geraci, A.; Rosselli, S.; Maggi, F.; Fiorini, D.; Ricciutelli, M.; Loizzo, M.R.; Bruno, M.; Pacetti, D. Fixed oil from seeds of narrow-leaved ash (F. angustifolia subsp. angustifolia): Chemical profile, antioxidant and antiproliferative activities. Food Res. Int. 2019, 119, 369–377. [Google Scholar] [CrossRef]
- Guarcello, R.; Gaglio, R.; Todaro, A.; Alfonzo, A.; Schicchi, R.; Cirlincione, F.; Moschetti, G.; Francesca, N. Insights Into the Cultivable Microbial Ecology of “Manna” Ash Products Extracted From Fraxinus angustifolia (Oleaceae) Trees in Sicily, Italy. Front. Microbiol. 2019, 10, 984. [Google Scholar] [CrossRef]
- Minà Palumbo, F. Monografia sulla coltivazione del frassino. Agric. Lal. 1876, 1875, 19–25. [Google Scholar]
- Falci, R. Il frassino da manna in Sicilia. Boll. R. Orto Bot. Giard. Col. Palermo 1910, 9, 145–168. [Google Scholar]
- Weber, J.L. Informativeness of human (dC-dA) n (dG-dT) n polymorphisms. Genomics 1990, 7, 524–530. [Google Scholar] [CrossRef]
- Morgante, M.; Olivieri, A.M. PCR-amplified microsatellites as markers in plant genetics. Plant J. 1993, 3, 175–182. [Google Scholar] [CrossRef]
- Bacchi, M.; Leone, M.; Mercati, F.; Preiti, G.; Sunseri, F.; Monti, M. Agronomic Evaluation and Genetic Characterization of Different Accessions in Lentil (Lens culinaris Medik.). Ital. J. Agron. 2010, 5, 303. [Google Scholar] [CrossRef]
- Jiao, Y.; Jia, H.; Li, X.; Chai, M.; Jia, H.; Chen, Z.; Wang, G.; Chai, C.; van de Weg, E.; Gao, Z. Development of simple sequence repeat (SSR) markers from a genome survey of Chinese bayberry (Myrica rubra). BMC Genom. 2012, 13, 201. [Google Scholar] [CrossRef] [Green Version]
- Mercati, F.; Longo, C.; Poma, D.; Araniti, F.; Lupini, A.; Mammano, M.M.; Fiore, M.C.; Abenavoli, M.R.; Sunseri, F. Genetic variation of an Italian long shelf-life tomato (Solanum lycopersicon L.) collection by using SSR and morphological fruit traits. Genet. Resour. Crop Evol. 2015, 62, 721–732. [Google Scholar] [CrossRef]
- Mercati, F.; Catarcione, G.; Paolacci, A.R.; Abenavoli, M.R.; Sunseri, F.; Ciaffi, M. Genetic diversity and population structure of an Italian landrace of runner bean (Phaseolus coccineus L.): Inferences for its safeguard and on-farm conservation. Genetica 2015, 143, 473–485. [Google Scholar] [CrossRef] [PubMed]
- King, R.A.; Ferris, C. Chloroplast DNA phylogeography of Alnus glutinosa (L.) Gaertn. Mol. Ecol. 1998, 7, 1151–1161. [Google Scholar] [CrossRef]
- Palmé, A.E. Chloroplast DNA variation, postglacial recolonization and hybridization in hazel, Corylus avellana. Mol. Ecol. 2002, 11, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Rajora, O.P.; Dancik, B.P. Chloroplast DNA inheritance in Populus. Theor. Appl. Genet. 1992, 84, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Dumolin, S.; Demesure, B.; Petit, R.J. Inheritance of chloroplast and mitochondrial genomes in pedunculate oak investigated with an efficient PCR method. Theor. Appl. Genet. 1995, 91, 1253–1256. [Google Scholar] [CrossRef]
- Harbourne, M.E.; Douglas, G.C.; Waldren, S.; Hodkinson, T.R. Characterization and primer development for amplification of chloroplast microsatellite regions of Fraxinus excelsior. J. Plant Res. 2005, 118, 339–341. [Google Scholar] [CrossRef]
- Erichsen, E.O.; Budde, K.B.; Sagheb-Talebi, K.; Bagnoli, F.; Vendramin, G.G.; Hansen, O.K. Hyrcanian forests—Stable rear-edge populations harbouring high genetic diversity of Fraxinus excelsior, a common European tree species. Divers. Distrib. 2018, 24, 1521–1533. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, E.A. Relationships of cereal crops and other grasses. PNAS 1998, 95, 2005–2010. [Google Scholar] [CrossRef] [Green Version]
- Leitch, I.J.; Chase, M.W.; Bennett, M.D. Phylogenetic Analysis of DNA C-values Provides Evidence for a Small Ancestral Genome Size in Flowering Plants. Ann. Bot. 1998, 82, 85–94. [Google Scholar] [CrossRef]
- Doležel, J.; Bartoš, J. Plant DNA Flow Cytometry and Estimation of Nuclear Genome Size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
- Petrov, D.A.; Sangster, T.A.; Johnston, J.S.; Hartl, D.L.; Shaw, K.L. Evidence for DNA Loss as a Determinant of Genome Size. Science 2000, 287, 1060–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennetzen, J.L.; Ma, J.; Devos, K.M. Mechanisms of Recent Genome Size Variation in Flowering Plants. Ann. Bot. 2005, 95, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harkess, A.; Mercati, F.; Abbate, L.; McKain, M.; Pires, J.C.; Sala, T.; Sunseri, F.; Falavigna, A.; Leebens-Mack, J. Retrotransposon Proliferation Coincident with the Evolution of Dioecy in Asparagus. G3 Genes Genomes Genet. 2016, 6, 2679–2685. [Google Scholar] [CrossRef]
- Murray, B.G. When does Intraspecific C-value Variation become Taxonomically Significant? Ann. Bot. 2005, 95, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Garnatje, T.; Garcia, S.; Vilatersana, R.; Vallès, J. Genome Size Variation in the Genus Carthamus (Asteraceae, Cardueae): Systematic Implications and Additive Changes During Allopolyploidization. Ann. Bot. 2006, 97, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Raimondo, F.M.; Bonomo, R.; Lentini, F. Il frassino da manna in Sicilia, pianta officinale da rivalutare. Giorn. Bot. Ital. 1980, 114, 141–142. [Google Scholar]
- Otto, F.J. Preparation and Staining of Cells for High-Resolution DNA Analysis. In Flow Cytometry and Cell Sorting; Radbruch, A., Ed.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 65–68. ISBN 978-3-662-02785-1. [Google Scholar]
- Weising, K.; Gardner, R.C. A set of conserved PCR primers for the analysis of simple sequence repeat polymorphisms in chloroplast genomes of dicotyledonous angiosperms. Genome 1999, 42, 9–19. [Google Scholar] [CrossRef]
- Lefort, F.; Brachet, S.; Frascaria-Lacoste, N.; Edwards, K.J.; Douglas, G.C. Identification and characterization of microsatellite loci in ash (Fraxinus excelsior L.) and their conservation in the olive family (Oleaceae). Mol. Ecol. 1999, 8, 1088–1089. [Google Scholar] [CrossRef]
- Brachet, S.; Jubier, M.F.; Richard, M.; Jung-Muller, B.; Frascaria-Lacoste, N. Rapid identification of microsatellite loci using 5′ anchored PRC in the common ash Fraxinus excelsior. Mol. Ecol. 1999, 8, 160–163. [Google Scholar]
- Aggarwal, R.K.; Allainguillaume, J.; Bajay, M.M.; Barthwal, S.; Bertolino, P.; Chauhan, P.; Consuegra, S.; Croxford, A.; Dalton, D.L.; Belder, E.D.; et al. Permanent Genetic Resources added to Molecular Ecology Resources Database 1 August 2010–30 September 2010. Mol. Ecol. Resour. 2011, 11, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Garfì, G.; Mercati, F.; Fontana, I.; Collesano, G.; Pasta, S.; Vendramin, G.G.; De Michele, R.; Carimi, F. Habitat features and genetic integrity of wild grapevine Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi populations: A case study from Sicily. Flora Morphol. Distrib. Funct. Ecol. Plants 2013, 208, 538–548. [Google Scholar] [CrossRef]
- Bruvo, R.; Michiels, N.K.; D’souza, T.G.; Schulenburg, H. A simple method for the calculation of microsatellite genotype distances irrespective of ploidy level. Mol. Ecol. 2004, 13, 2101–2106. [Google Scholar] [CrossRef] [PubMed]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Ahmed, I. Adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. The Interpretation of Population Structure by F-Statistics with Special Regard to Systems of Mating. Evolution 1965, 19, 395–420. [Google Scholar] [CrossRef]
- Nei, M. Analysis of Gene Diversity in Subdivided Populations. PNAS 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J. Hierfstat, a package for r to compute and test hierarchical F-statistics. Mol. Ecol. Notes 2005, 5, 184–186. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Raymond, M.; Rousset, F. An Exact Test for Population Differentiation. Evolution 1995, 49, 1280–1283. [Google Scholar] [CrossRef] [PubMed]
- Hardy, O.J.; Vekemans, X. Spagedi: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol. Ecol. Notes 2002, 2, 618–620. [Google Scholar] [CrossRef] [Green Version]
- Siljak-Yakovlev, S.; Temunović, M.; Robin, O.; Raquin, C.; Frascaria-Lacoste, N. Molecular–cytogenetic studies of ribosomal RNA genes and heterochromatin in three European Fraxinus species. Tree Genet. Genomes 2014, 10, 231–239. [Google Scholar] [CrossRef]
- Ivanova, D.; Dimitrova, D.; Vladimirov, V. Chromosome numbers of selected woody species from the Bulgarian. Phytol. Balc. 2006, 12, 79–84. [Google Scholar]
- Jarni, K.; Westergren, M.; Kraigher, H.; Brus, R. Morphological variability of Fraxinus angustifolia Vahl in the north-western Balkans. Acta Soc. Bot. Pol. 2011, 80, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Ziegenhagen, B.; Fladung, M. DNA Markers for Identification and Evaluation of Genetic Resources in Forest Trees: Case Studies in Abies, Picea and Populus. In Molecular Marker Systems in Plant Breeding and Crop Improvement; Lörz, H., Wenzel, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 413–429. ISBN 978-3-540-26538-2. [Google Scholar]
- Schmuths, H.; Meister, A.; Horres, R.; Bachmann, K. Genome Size Variation among Accessions of Arabidopsis thaliana. Ann. Bot. 2004, 93, 317–321. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bigelow, C.A.; Jiang, Y. Ploidy Level and DNA Content of Perennial Ryegrass Germplasm as Determined by Flow Cytometry. HortScience 2009, 44, 2049–2052. [Google Scholar] [CrossRef] [Green Version]
- Díez, C.M.; Gaut, B.S.; Meca, E.; Scheinvar, E.; Montes-Hernandez, S.; Eguiarte, L.E.; Tenaillon, M.I. Genome size variation in wild and cultivated maize along altitudinal gradients. New Phytol. 2013, 199, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Eilam, T.; Anikster, Y.; Millet, E.; Manisterski, J.; Sagi-Assif, O.; Feldman, M. Genome size and genome evolution in diploid Triticeae species. Genome 2007, 50, 1029–1037. [Google Scholar] [CrossRef]
- Greilhuber, J. Intraspecific Variation in Genome Size in Angiosperms: Identifying its Existence. Ann. Bot. 2005, 95, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, J.; Rodriguez, E.; Doležel, J.; Santos, C. Flow Cytometric and Microscopic Analysis of the Effect of Tannic Acid on Plant Nuclei and Estimation of DNA Content. Ann. Bot. 2006, 98, 515–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besnard, G.; Khadari, B.; Baradat, P.; Bervillé, A. Olea europaea (Oleaceae) phylogeography based on chloroplast DNA polymorphism. Theor. Appl. Genet. 2002, 104, 1353–1361. [Google Scholar] [CrossRef] [PubMed]
- Jeandroz, S.; Roy, A.; Bousquet, J. Phylogeny and Phylogeography of the Circumpolar GenusFraxinus(OLEACEAE) Based on Internal Transcribed Spacer Sequences of Nuclear Ribosomal DNA. Mol. Phylogenet. Evol. 1997, 7, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Morand-Prieur, M.E.; Vedel, F.; Raquin, C.; Brachet, S.; Sihachakr, D.; Frascaria-Lacoste, N. Maternal inheritance of a chloroplast microsatellite marker in controlled hybrids between Fraxinus excelsior and Fraxinus angustifolia. Mol. Ecol. 2002, 11, 613–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, S. Recolonisation Postglaciaire de Quelques Taxons Tempérés en Europe: Une Approche Spatiale et Temporelle. Ph.D. Thesis, Cornell University, Marseille, France, 2002. [Google Scholar]
- Thomson, M.J.; Septiningsih, E.M.; Suwardjo, F.; Santoso, T.J.; Silitonga, T.S.; McCouch, S.R. Genetic diversity analysis of traditional and improved Indonesian rice (Oryza sativa L.) germplasm using microsatellite markers. Theor. Appl. Genet. 2007, 114, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Díaz, L.M.; Buendía, H.F.; Duque, M.C. Genetic diversity, seed size associations and population structure of a core collection of common beans (Phaseolus vulgaris L.). Theor. Appl. Genet. 2009, 119, 955–972. [Google Scholar] [CrossRef]
- Carimi, F.; Mercati, F.; De Michele, R.; Fiore, M.C.; Riccardi, P.; Sunseri, F. Intra-varietal genetic diversity of the grapevine (Vitis vinifera L.) cultivar ‘Nero d’Avola’ as revealed by microsatellite markers. Genet. Resour. Crop Evol. 2011, 58, 967. [Google Scholar] [CrossRef]
- Laidò, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; Marone, D.; Mastrangelo, A.M.; Papa, R.; De Vita, P. Genetic Diversity and Population Structure of Tetraploid Wheats (Triticum turgidum L.) Estimated by SSR, DArT and Pedigree Data. PLoS ONE 2013, 8, e67280. [Google Scholar] [CrossRef] [Green Version]
- Papi, R.M.; Spanos, K.A.; Kyriakidis, D.A. Genetic variation of Fraxinus angustifolia natural populations in Greece based on nuclear and chloroplast microsatellite markers. Eur. J. For. Res. 2012, 131, 1151–1161. [Google Scholar] [CrossRef]
- Fernandez-Manjarres, J.F.; Gerard, P.R.; Dufour, J.; Raquin, C.; Frascaria-Lacoste, N. Differential patterns of morphological and molecular hybridization between Fraxinus excelsior L. and Fraxinus angustifolia Vahl (Oleaceae) in eastern and western France. Mol. Ecol. 2006, 15, 3245–3257. [Google Scholar] [CrossRef]
- Gerard, P.R.; Fernandez-Manjarres, J.F.; Frascaria-Lacoste, N. Temporal cline in a hybrid zone population between Fraxinus excelsior L. and Fraxinus angustifolia Vahl. Mol. Ecol. 2006, 15, 3655–3667. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, D.D.; Basak, J.; Gaudeul, M.; Cruaud, C.; Bertolino, P.; Frascaria-Lacoste, N.; Bousquet, J. The Phylogeny and Biogeographic History of Ashes (Fraxinus, Oleaceae) Highlight the Roles of Migration and Vicariance in the Diversification of Temperate Trees. PLoS ONE 2013, 8, e80431. [Google Scholar] [CrossRef] [PubMed]
- Ilardi, V.; Raimondo, F.M. The genus Fraxinus, L. (Oleaceae) in Sicily. Flora Mediterr. 1999, 9, 305–318. [Google Scholar]
- Schicchi, R.; Marino, P. Taxa relitti della flora forestale siciliana e problemi di conservazione. Relict taxa of the Sicilian forest flora and conservation issues. Biogeographia 2011, 30, 141–150. [Google Scholar]
- Schicchi, R.; Camarda, L.; Di Stefano, V.; Spadaro, V.; Pitonzo, R. Analisi qualitativa dei costituenti cumarinici presenti in campioni di manna prodotti da esemplari di Fraxinus angustifolia (Oleaceae) delle Madonie. In Proceedings of the Atti Del 102 Congresso Della Societa Botanica Italiana, Palermo, Italy, 26–29 September 2007; p. 388. [Google Scholar]
- Minà Palumbo, F. Notizie sui frassini di Sicilia e sulla coltivazione dell’Amolleo in Castelbuono. Stamp. Leg. F. Ruffino Palermo 1847, 17. [Google Scholar]
- Donoghue, M.J.; Bell, C.D.; Li, J. Phylogenetic Patterns in Northern Hemisphere Plant Geography. Int. J. Plant Sci. 2001, 162, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Magri, D.; Fineschi, S.; Bellarosa, R.; Buonamici, A.; Sebastiani, F.; Schirone, B.; Simeone, M.C.; Vendramin, G.G. The distribution of Quercus suber chloroplast haplotypes matches the palaeogeographical history of the western Mediterranean. Mol. Ecol. 2007, 16, 5259–5266. [Google Scholar] [CrossRef]
- Hayes, A.; Kucera, M.; Kallel, N.; Sbaffi, L.; Rohling, E.J. Glacial Mediterranean sea surface temperatures based on planktonic foraminiferal assemblages. Quat. Sci. Rev. 2005, 24, 999–1016. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Local Varieties | Ploidy Level | 2C DNA Value * | Apical Leaf (Length/Width Ratio) * | Samara Stalk Length (cm) * | Number of Samaras/Raceme * | Flowering Time | Apical Buds Colour |
|---|---|---|---|---|---|---|---|---|
| Manna ash | Abbassa cappeddu (3) | 2n | 1.540 ± 0.01 | 0.21 ± 0.01 | 0.81 ± 0.07 | 5.51 ± 0.20 | November–January | Red |
| Baciciu (4) | 2n | 1.544 ± 0.03 | 0.42 ± 0.02 | 1.12 ± 0.07 | 3.50 ± 1.02 | November–January | Red | |
| Cavolo (3) | 2n | 1.542 ± 0.02 | 0.23 ± 0.09 | 2.11 ± 0.06 | 5.50 ± 0.07 | November–January | Red | |
| Macigna (3) | 2n | 1.541 ± 0.01 | 0.42 ± 0.03 | 0.79 ± 0.02 | 6.01 ± 0.70 | November–January | Red | |
| Nivuru (3) | 2n | 1.541 ± 0.04 | 0.35 ± 0.01 | 3.04 ± 0.01 | 7.10 ± 0.62 | November–January | Red | |
| Nsiriddu (3) | 2n | 1.544 ± 0.03 | 0.24 ± 0.08 | 1.15 ± 0.05 | 5.09 ± 0.07 | November–January | Red | |
| Russu (3) | 2n | 1.542 ± 0.04 | 0.32 ± 0.07 | 0.68 ± 0.07 | 5.52 ± 0.27 | November–January | Red | |
| Sarvaggio (3) | 2n | 1.539 ± 0.02 | 0.31 ± 0.05 | 0.77 ± 0.02 | 5.01 ± 0.19 | November–January | Red | |
| Verdello (7) | 2n | 1.542 ± 0.05 | 0.21 ± 0.01 | 0.76 ± 0.08 | 6.53 ± 0.35 | November–January | Red | |
| Frassino monumentale (1) | 2n | 1.541 ± 0.01 | 0.23 ± 0.03 | 0.92 ± 0.03 | 7.05 ± 0.10 | November–January | Red | |
| Nsiriddu monumentale (1) | 2n | 1.544 ± 0.01 | 0.22 ± 0.01 | 0.91 ± 0.05 | 7.03 ± 0.32 | November–January | Red | |
| mean | - | 1.542 ** | 0.28 ** | 1.18 ** | 5.80 ** | - | - | |
| F. ornus (4) | 2n | 1.98 ± 0.01 | 0.52 ± 0.02 | 0.41 ± 0.03 | 46.02 ± 2.05 | May–June | White |
| Samples/Local Varieties | Nh # | ccmp2 | ccmp3 | ccmp4 | ccmp6 | ccmp7 | ccmp10 | CPFRAX6 | atpB/rbcL | matK | Haplotypes * |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Abbassa cappeddu (3) | 1 | 194 | 97 | 140 | 98 | 117 | 104 | 365 | 157 | 253 | H10 |
| Baciciu (4) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 |
| Cavolo (3) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 |
| Macigna (3) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 |
| Nivuru (3) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 |
| Nsiriddu (3) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 366 | 157 | 253 | H5 a |
| Russu (3) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 |
| Sarvaggio (3) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 |
| Verdello (7) | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 | |
| 3 | 194 | 97 | 140 | 98 | 117 | 103 | 366 | 157 | 253 | H5 a | |
| 194 | 97 | 140 | 98 | 117 | 104 | 365 | 157 | 253 | H10 | ||
| Frassino monumentale (1) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 |
| Nsiriddu monumentale (1) | 1 | 194 | 97 | 140 | 98 | 117 | 103 | 365 | 157 | 253 | H5 |
| F. ornus (4) | 1 | 190 | 97 | 140 | 99 | 117 | 106 | 364 | 157 | 253 | H19 |
| Group. | FAN (Italy) | FAN | HYB | FEX |
|---|---|---|---|---|
| FAN (Italy) | 0.000 | 0.284 | 0.536 | 0.874 |
| FAN | 0.066 | 0.000 | 0.159 | 0.437 |
| HYB | 0.107 | 0.030 | 0.000 | 0.110 |
| FEX | 0.163 | 0.084 | 0.028 | 0.000 |
| Group ID. | n. | I | Ho | He | Fis | AR |
|---|---|---|---|---|---|---|
| Bulgaria | 8 | 0.964 | 0.615 | 0.525 | −0.208 | 2.73 |
| Croatia1 | 8 | 1.156 | 0.591 | 0.563 | −0.025 | 3.05 |
| Croatia2 | 8 | 1.000 | 0.614 | 0.523 | −0.135 | 2.71 |
| France1 | 7 | 1.259 | 0.645 | 0.632 | −0.030 | 3.32 |
| France2 | 7 | 1.320 | 0.803 | 0.680 | −0.216 * | 3.42 |
| France3 | 8 | 1.316 | 0.614 | 0.658 | 0.039 | 3.32 |
| France4 | 8 | 1.171 | 0.659 | 0.607 | −0.074 | 3.03 |
| France5 | 8 | 1.094 | 0.489 | 0.572 | 0.204 ** | 2.93 |
| France6 | 8 | 1.256 | 0.648 | 0.626 | −0.042 | 3.22 |
| France7 | 8 | 0.953 | 0.659 | 0.513 | −0.269 | 2.63 |
| Hungary | 7 | 0.801 | 0.805 | 0.506 | −0.607 *** | 2.28 |
| Italy1 | 16 | 1.170 | 0.591 | 0.553 | 0.040 *** | 2.96 |
| Italy2 | 8 | 1.032 | 0.623 | 0.541 | −0.112 *** | 2.85 |
| Macedonia | 8 | 1.325 | 0.750 | 0.651 | −0.152 | 3.39 |
| Montenegro | 8 | 1.215 | 0.659 | 0.607 | −0.087 | 3.13 |
| Portugal1 | 6 | 1.185 | 0.621 | 0.624 | 0.017 | 3.17 |
| Portugal2 | 8 | 1.029 | 0.534 | 0.529 | 0.038 | 2.86 |
| Serbia1 | 8 | 1.323 | 0.623 | 0.623 | −0.022 | 3.40 |
| Serbia2 | 6 | 1.193 | 0.667 | 0.580 | −0.159 | 3.26 |
| Turkey | 3 | 1.089 | 0.727 | 0.601 | −0.241 | 3.55 |
| Ukraine | 8 | 1.040 | 0.610 | 0.553 | −0.109 | 2.88 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbate, L.; Mercati, F.; Di Noto, G.; Heuertz, M.; Carimi, F.; Fatta del Bosco, S.; Schicchi, R. Genetic Distinctiveness Highlights the Conservation Value of a Sicilian Manna Ash Germplasm Collection Assigned to Fraxinus angustifolia (Oleaceae). Plants 2020, 9, 1035. https://doi.org/10.3390/plants9081035
Abbate L, Mercati F, Di Noto G, Heuertz M, Carimi F, Fatta del Bosco S, Schicchi R. Genetic Distinctiveness Highlights the Conservation Value of a Sicilian Manna Ash Germplasm Collection Assigned to Fraxinus angustifolia (Oleaceae). Plants. 2020; 9(8):1035. https://doi.org/10.3390/plants9081035
Chicago/Turabian StyleAbbate, Loredana, Francesco Mercati, Giuseppe Di Noto, Myriam Heuertz, Francesco Carimi, Sergio Fatta del Bosco, and Rosario Schicchi. 2020. "Genetic Distinctiveness Highlights the Conservation Value of a Sicilian Manna Ash Germplasm Collection Assigned to Fraxinus angustifolia (Oleaceae)" Plants 9, no. 8: 1035. https://doi.org/10.3390/plants9081035