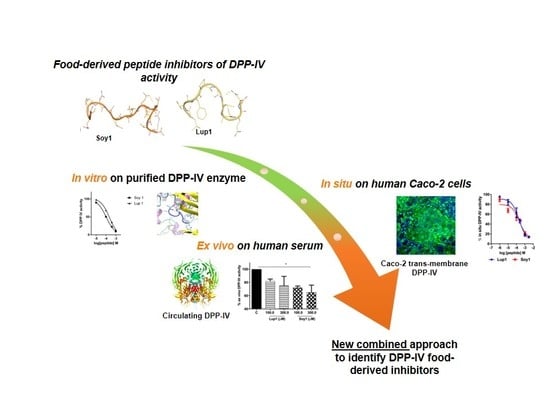
Soybean- and Lupin-Derived Peptides Inhibit DPP-IV Activity on In Situ Human Intestinal Caco-2 Cells and Ex Vivo Human Serum


 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
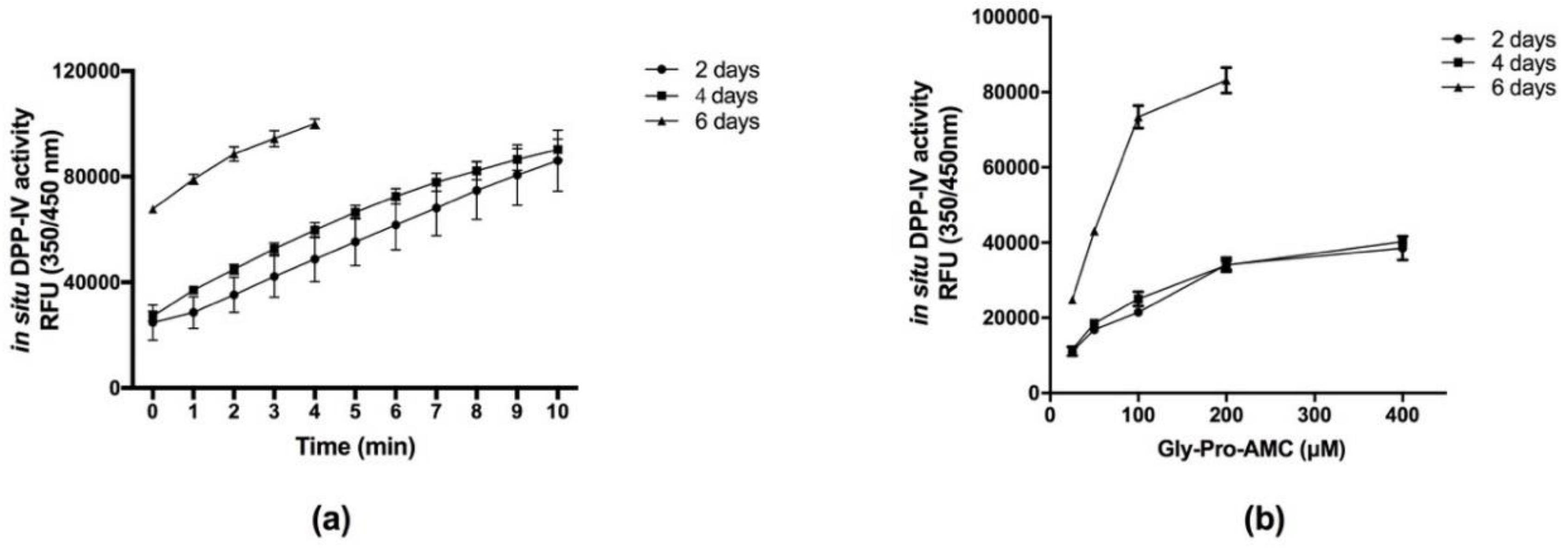
2.3. In Situ Cell-Based DPP-IV Activity Assay Optimization
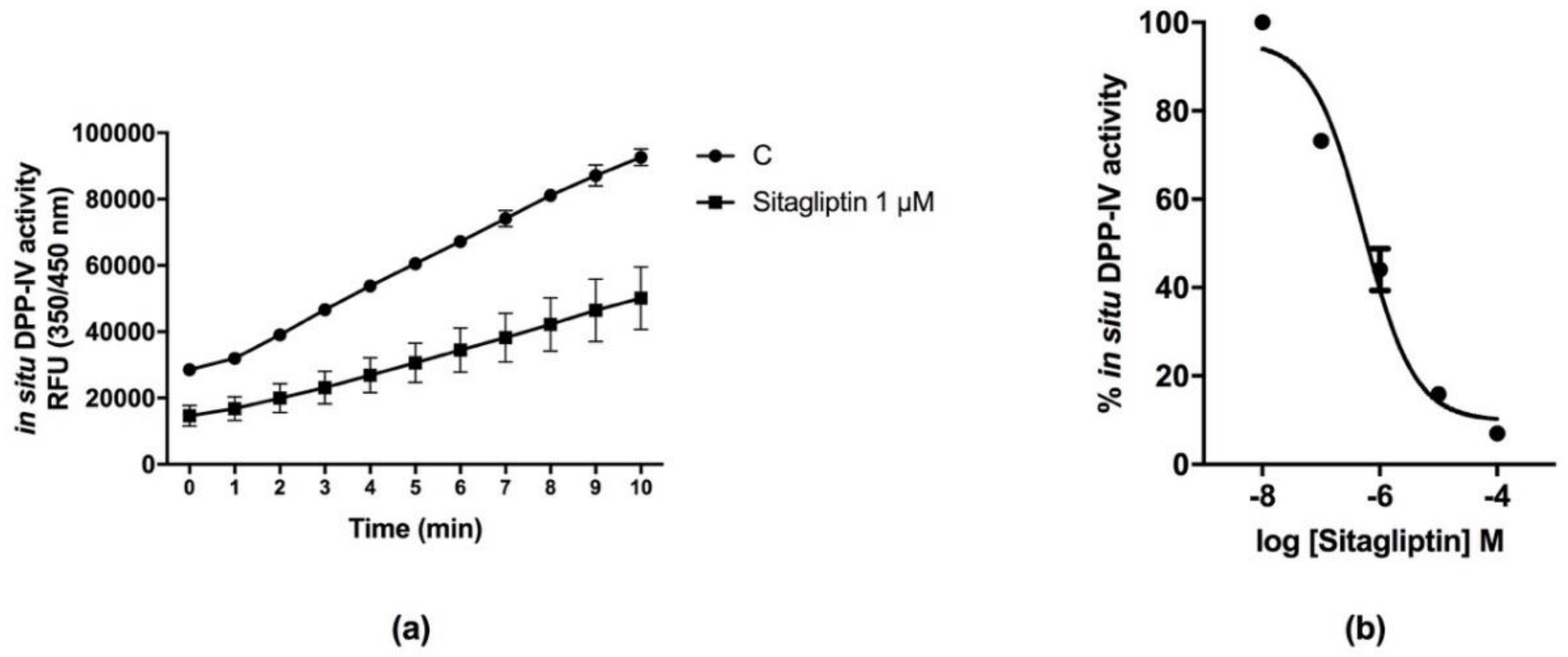
2.4. In Situ Evaluation of the Inhibitory Effect of Sitagliptin, Lup1, and Soy1 on In Situ DPP-IV Activity
2.5. Ex Vivo DPP-IV Activity Assay
2.6. Statistical Analysis
3. Results
3.1. Development and Optimization of a Caco-2 Cell-Based DPP-IV Assay
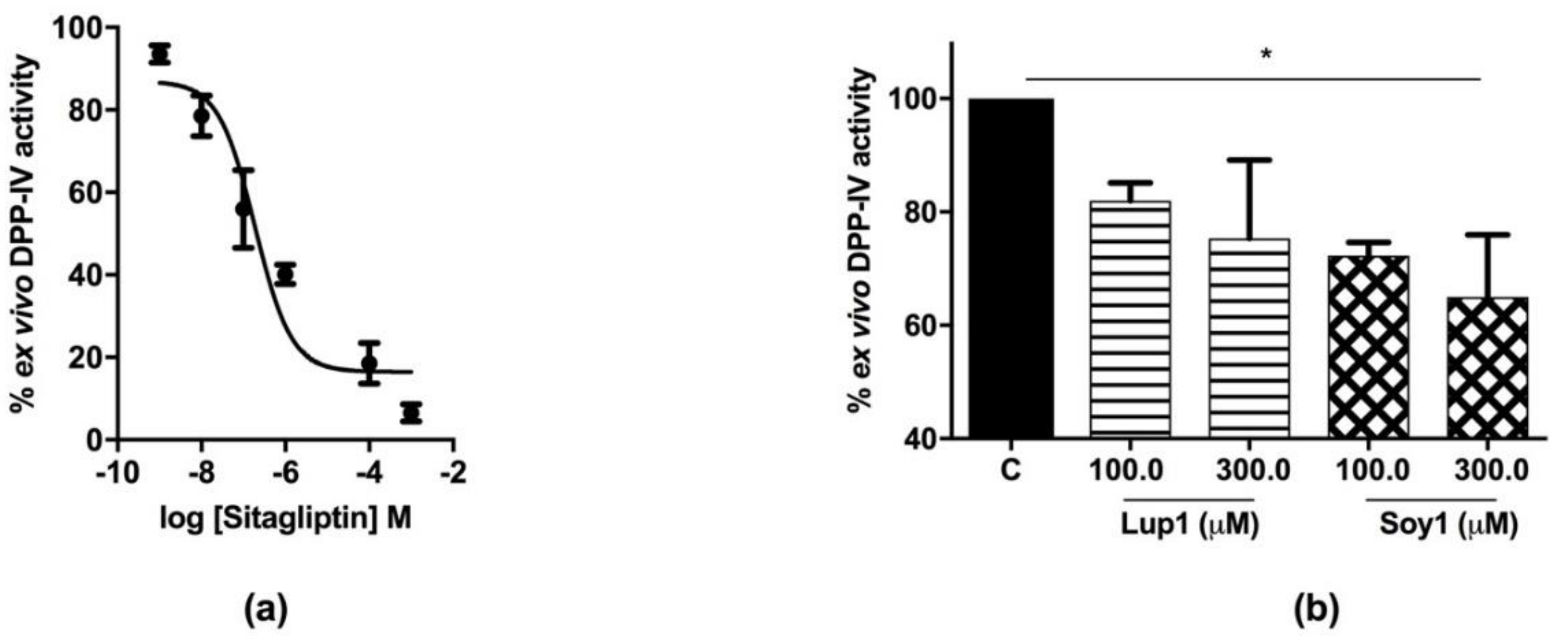
3.2. Sitagliptin Inhibits DPP-IV Activity in Caco-2 Cells
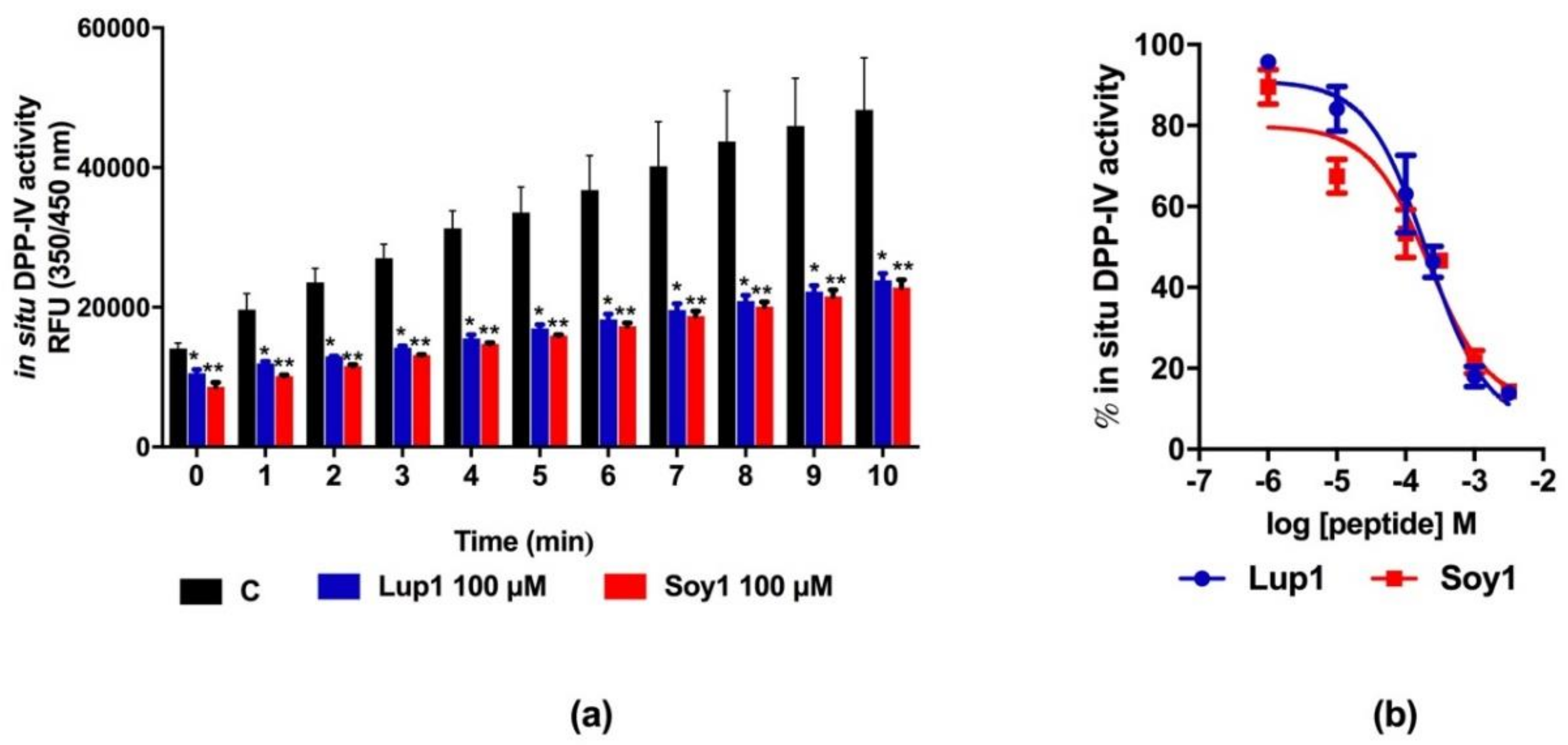
3.3. Peptides Lup1 and Soy1 Inhibit DPP-IV Activity Expressed in Caco-2 Cells
3.4. Circulating DPP-IV Inhibition by Peptides Lup1 and Soy1
4. Discussion
4.1. Development and Validation of a Cell-Based DPP-IV Activity Assay Using Human Intestinal Caco-2 Cells and an Ex Vivo Assay on Circulating DPP-IV Activity in Human Serum
4.2. Lup1 and Soy1 Inhibit DPP-IV Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMC | amido-4-methylcoumarin hydrobromide |
| BB | brush border |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DPP-IV | dipeptidyl peptidase IV |
| FBS | fetal bovine serum |
| GLP-1 | glucagon-like peptide-1 |
| GIP | gastrointestinal insulinotropic peptide |
| IC50 | 50% inhibitory concentration |
| PBS | phosphate-buffered saline |
| RFU | relative fluorescence unit |
| T2DM | type 2 diabetes |
References
- Abbott, C.A.; Baker, E.; Sutherland, G.R.; McCaughan, G.W. Genomic organization, exact localization, and tissue expression of the human CD26 (dipeptidyl peptidase IV) gene. Immunogenetics 1994, 40, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Röhrborn, D.; Wronkowitz, N.; Eckel, J. DPP4 in Diabetes. Front. Immunol. 2015, 6, 386. [Google Scholar] [CrossRef] [PubMed]
- Nargis, T.; Chakrabarti, P. Significance of circulatory DPP4 activity in metabolic diseases. IUBMB Life 2018, 70, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Dipeptidyl peptidase-IV inhibitory activity of dairy protein hydrolysates. Int. Dairy J. 2012, 25, 97–102. [Google Scholar] [CrossRef]
- Nauck, M.A.; Baller, B.; Meier, J.J. Gastric inhibitory polypeptide and glucagon-like peptide-1 in the pathogenesis of type 2 diabetes. Diabetes 2004, 53 (Suppl. 3), S190–S196. [Google Scholar] [CrossRef] [PubMed]
- Doupis, J.; Veves, A. DPP4 inhibitors: A new approach in diabetes treatment. Adv. Ther. 2008, 25, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Dicker, D. DPP-4 inhibitors: impact on glycemic control and cardiovascular risk factors. Diabetes Care 2011, 34 (Suppl. 2), S276–S278. [Google Scholar] [CrossRef] [PubMed]
- Scheen, A.J. Cardiovascular effects of new oral glucose-lowering agents: DPP-4 and SGLT-2 inhibitors. Circ. Res. 2018, 122, 1439–1459. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Chan, C.B.; Wu, J. Egg white ovotransferrin-derived ACE inhibitory peptide ameliorates angiotensin II-stimulated insulin resistance in skeletal muscle cells. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Lima-Cabello, E.; Alche, V.; Foley, R.C.; Andrikopoulos, S.; Morahan, G.; Singh, K.B.; Alche, J.D.; Jimenez-Lopez, J.C. Narrow-leafed lupin (Lupinus angustifolius L.) β-conglutin proteins modulate the insulin signaling pathway as potential type 2 diabetes treatment and inflammatory-related disease amelioration. Mol. Nutr. Food Res. 2017, 61, 1600819. [Google Scholar] [CrossRef] [PubMed]
- Jao, C.L.; Hung, C.C.; Tung, Y.S.; Lin, P.Y.; Chen, M.C.; Hsu, K.C. The development of bioactive peptides from dietary proteins as a dipeptidyl peptidase iv inhibitor for the management of type 2 diabetes. BioMedicine 2015, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Peptides derived from soy and lupin protein as dipeptidyl-peptidase IV inhibitors: in vitro biochemical screening and in silico molecular modeling study. J. Agric. Food Chem. 2016, 64, 9601–9606. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Paolella, S.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Identification of novel dipeptidyl peptidase IV (DPP-IV) inhibitory peptides in camel milk protein hydrolysates. Food Chem. 2018, 244, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Mazzocchi, C.; Paolella, S.; FitzGerald, R.J. Release of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from milk protein isolate (MPI) during enzymatic hydrolysis. Food Res. Int. 2017, 94, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Darmoul, D.; Lacasa, M.; Baricault, L.; Marguet, D.; Sapin, C.; Trotot, P.; Barbat, A.; Trugnan, G. Dipeptidyl peptidase IV (CD 26) gene expression in enterocyte-like colon cancer cell lines HT-29 and Caco-2. Cloning of the complete human coding sequence and changes of dipeptidyl peptidase IV mRNA levels during cell differentiation. J. Biol. Chem. 1992, 267, 4824–4833. [Google Scholar] [PubMed]
- Yoshioka, M.; Erickson, R.H.; Matsumoto, H.; Gum, E.; Kim, Y.S. Expression of dipeptidyl aminopeptidase IV during enterocytic differentiation of human colon cancer (Caco-2) cells. Int. J. Cancer 1991, 47, 916–921. [Google Scholar] [CrossRef] [PubMed]
- Lambeir, A.M.; Durinx, C.; Scharpé, S.; De Meester, I. Dipeptidyl-peptidase IV from bench to bedside: an update on structural properties, functions, and clinical aspects of the enzyme DPP IV. Crit. Rev. Clin. Lab. Sci. 2003, 40, 209–294. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.; Salles, T.A.; Arruda-Junior, D.F.; Campos, L.C.G.; Pereira, A.C.; Barreto, A.L.T.; Antonio, E.L.; Mansur, A.J.; Tucci, P.J.F.; Krieger, J.E.; et al. Circulating dipeptidyl peptidase iv activity correlates with cardiac dysfunction in human and experimental heart failure. Circ. Hear. Fail. 2013, 6, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Aiello, G.; Vistoli, G.; Zanoni, C.; Arnoldi, A.; Sambuy, Y.; Ferruzza, S.; Ranaldi, G. A multidisciplinary investigation on the bioavailability and activity of peptides from lupin protein. J. Funct. Foods 2016, 24, 297–306. [Google Scholar] [CrossRef]
- Zanoni, C.; Aiello, G.; Arnoldi, A.; Lammi, C. Investigations on the hypocholesterolaemic activity of LILPKHSDAD and LTFPGSAED, two peptides from lupin beta-conglutin: Focus on LDLR and PCSK9 pathways. J. Funct. Foods 2017, 32, 1–8. [Google Scholar] [CrossRef]
- Natoli, M.; Leoni, B.D.; D’Agnano, I.; D’Onofrio, M.; Brandi, R.; Arisi, I.; Zucco, F.; Felsani, A. Cell growing density affects the structural and functional properties of caco-2 differentiated monolayer. J. Cell Physiol. 2011, 226, 1531–1543. [Google Scholar] [CrossRef] [PubMed]
- Caron, J.; Domenger, D.; Dhulster, P.; Ravallec, R.; Cudennec, B. Using Caco-2 cells as novel identification tool for food-derived DPP-IV inhibitors. Food Res. Int. 2017, 92, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Ferruzza, S.; Rossi, C.; Scarino, M.L.; Sambuy, Y. A protocol for differentiation of human intestinal Caco-2 cells in asymmetric serum-containing medium. Toxicol. Vitro 2012, 26, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- Herman, G.; Stevens, C.; Vandyck, K.; Bergman, A.; Yi, B.; Desmet, M.; Snyder, K.; Hilliard, D.; Tanen, M.; Tanaka, W. Pharmacokinetics and pharmacodynamics of sitagliptin, an inhibitor of dipeptidyl peptidase iv, in healthy subjects: results from two randomized, double-blind, placebo-controlled studies with single oral doses. Clin. Pharmacol. Ther. 2005, 78, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Varin, E.M.; Gladanac, B.; Campbell, J.E.; Ussher, G.R.; Baggio, L.L.; Yusta, B.; Ayala, J.; Burmeister, M.A.; Matthews, D.; et al. Cellular sites and mechanisms linking reduction of dipeptidyl peptidase-4 activity to control of incretin hormone action and glucose homeostasis. Cell Metab. 2017, 25, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Carr, R.D. Drug development from the bench to the pharmacy: with special reference to dipeptidyl peptidase-4 inhibitor development. Diabet. Med. 2016, 33, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J.; Tahrani, A.A.; Barnett, A.H. Future glucose-lowering drugs for type 2 diabetes. Lancet Diabetes Endocrinol. 2016, 4, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Ahrén, B.; Foley, J.E. Improved glucose regulation in type 2 diabetic patients with dpp-4 inhibitors: ocus on alpha and beta cell function and lipid metabolism. Diabetologia 2016, 59, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.; Li-Chan, E.C. Overview of food products and dietary constituents with antidiabetic properties and their putative mechanisms of action: a natural approach to complement pharmacotherapy in the management of diabetes. Mol. Nutr. Food Res. 2014, 58, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Investigation of the potential of hemp, pea, rice and soy protein hydrolysates as a source of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides. Food Dig. Res. Curr. Opin. 2015, 6, 19–29. [Google Scholar]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Aiello, G.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A.; Vistoli, G.; Lammi, C. Behavior of three hypocholesterolemic peptides from soy protein in an intestinal model based on differentiated Caco-2 cell. J. Funct. Foods 2018, 45, 363–370. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lammi, C.; Bollati, C.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A. Soybean- and Lupin-Derived Peptides Inhibit DPP-IV Activity on In Situ Human Intestinal Caco-2 Cells and Ex Vivo Human Serum. Nutrients 2018, 10, 1082. https://doi.org/10.3390/nu10081082
Lammi C, Bollati C, Ferruzza S, Ranaldi G, Sambuy Y, Arnoldi A. Soybean- and Lupin-Derived Peptides Inhibit DPP-IV Activity on In Situ Human Intestinal Caco-2 Cells and Ex Vivo Human Serum. Nutrients. 2018; 10(8):1082. https://doi.org/10.3390/nu10081082
Chicago/Turabian StyleLammi, Carmen, Carlotta Bollati, Simonetta Ferruzza, Giulia Ranaldi, Yula Sambuy, and Anna Arnoldi. 2018. "Soybean- and Lupin-Derived Peptides Inhibit DPP-IV Activity on In Situ Human Intestinal Caco-2 Cells and Ex Vivo Human Serum" Nutrients 10, no. 8: 1082. https://doi.org/10.3390/nu10081082