
Exposure of Human Gastric Cells to Oxidized Lipids Stimulates Pathways of Amino Acid Biosynthesis on a Genomic and Metabolomic Level

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Quantitative Recovery of Linoleic Acid and Its Peroxidation Products in the Basolateral Compartment of HGT-1 Cells after Apical Exposure
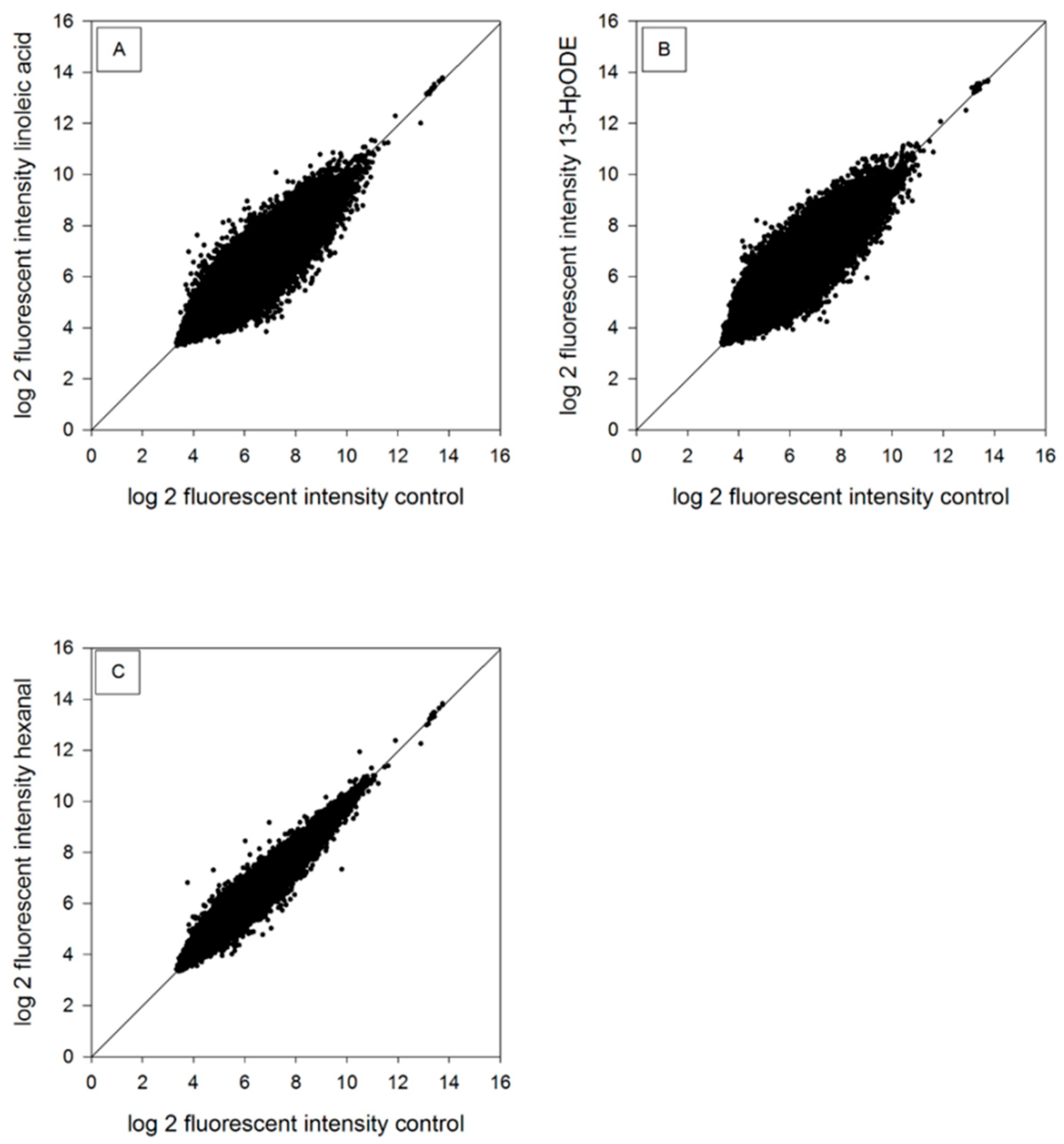
2.2. Genomic Analysis of RNA
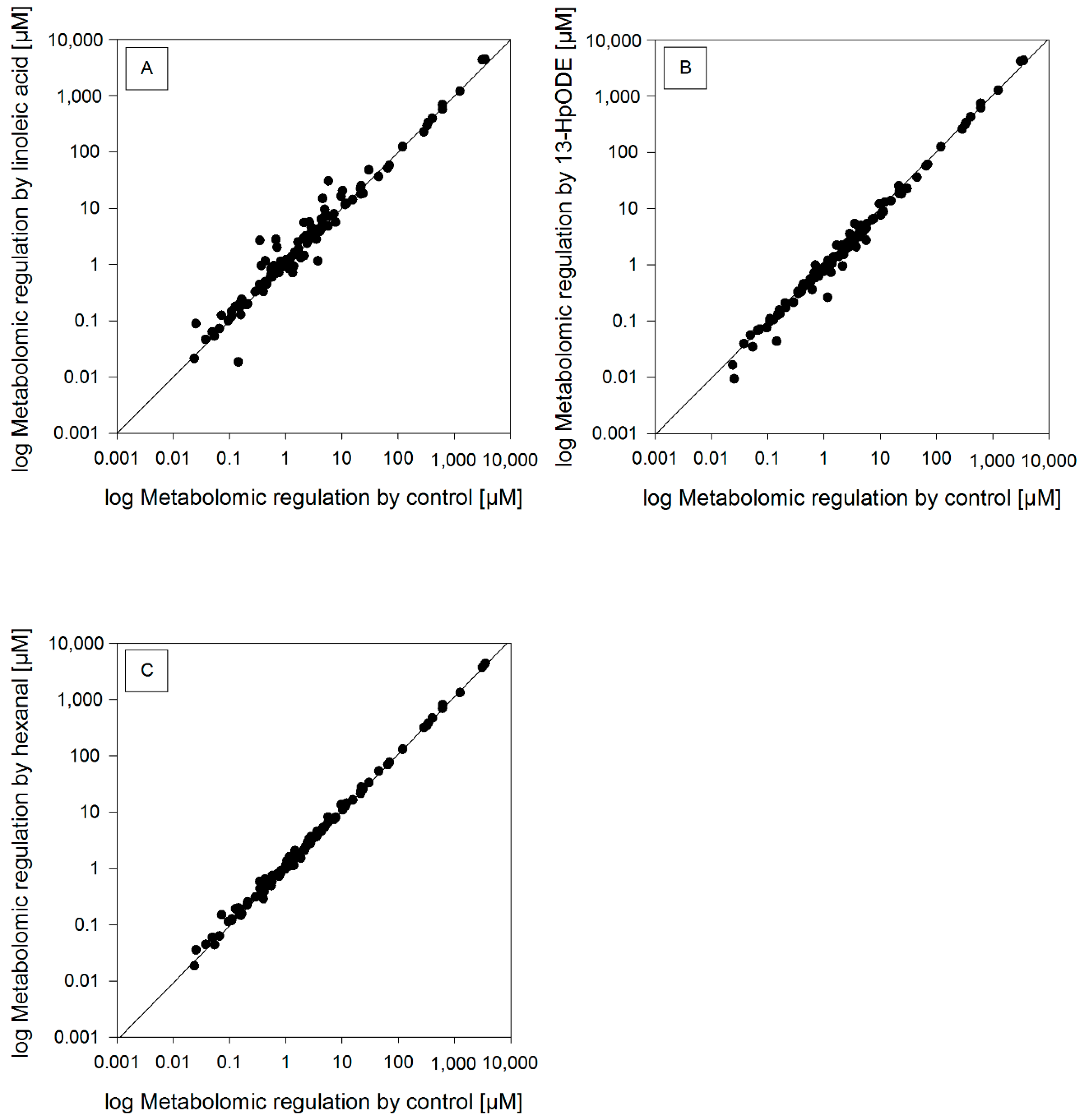
2.3. Metabolomic Analysis
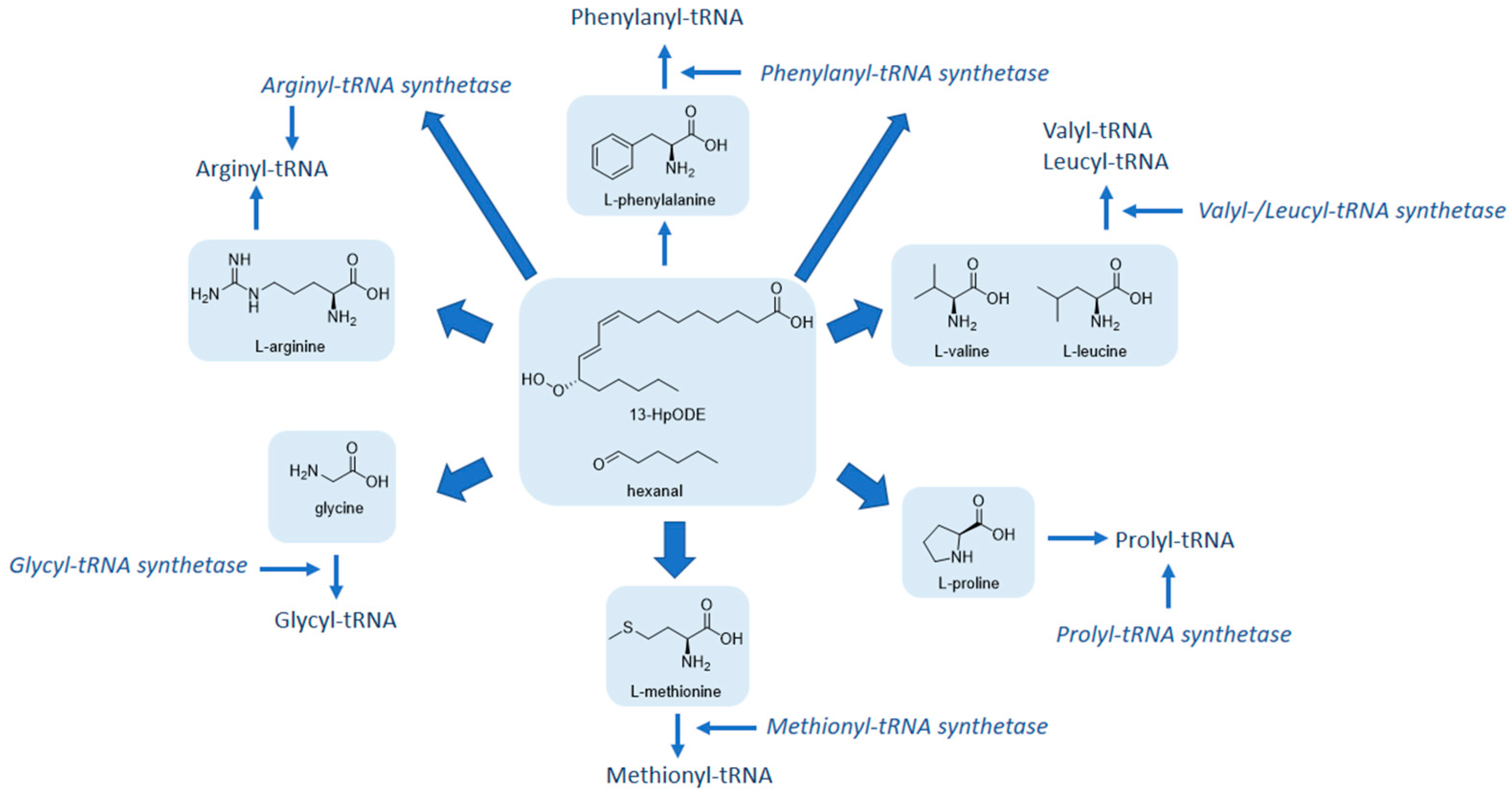
2.4. Combined Genomic and Metabolomic Effects of Linoleic Acid and Its Lipid Peroxidation Products on HGT-1 Cells: An Integrated Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Exposure Study
4.4. Quantitation of Linoleic Acid Peroxidation Products
4.5. Quantitation of Hexanal
4.6. Genome Analysis of RNA Regulation
4.7. Metabolomic Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Qi, Q.; Chu, A.Y.; Kang, J.H.; Huang, J.; Rose, L.M.; Jensen, M.K.; Liang, L.; Curhan, G.C.; Pasquale, L.R.; Wiggs, J.L.; et al. Fried food consumption, genetic risk, and body mass index: Gene-diet interaction analysis in three US cohort studies. BMJ 2014, 348, g1610. [Google Scholar] [CrossRef] [PubMed]
- Guallar-Castillon, P.; Rodriguez-Artalejo, F.; Fornes, N.S.; Banegas, J.R.; Etxezarreta, P.A.; Ardanaz, E.; Barricarte, A.; Chirlaque, M.D.; Iraeta, M.D.; Larranaga, N.L.; et al. Intake of fried foods is associated with obesity in the cohort of Spanish adults from the European Prospective Investigation into Cancer and Nutrition. Am. J. Clin. Nutr. 2007, 86, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Saleh, A.S.M.; Chen, J.; Shen, Q. Chemical alterations taken place during deep-fat frying based on certain reaction products: A review. Chem. Phys. Lipids 2012, 165, 662–681. [Google Scholar] [CrossRef] [PubMed]
- Choe, E.; Min, D.B. Mechanisms and factors for edible oil oxidation. Compr. Rev. Food Sci. Food Safe. 2006, 5, 169–186. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Lipid oxidation and improving the oxidative stability. Chem. Soc. Rev. 2010, 39, 4067–4079. [Google Scholar] [CrossRef]
- Lim, P.K.; Jinap, S.; Sanny, M.; Tan, C.P.; Khatib, A. The Influence of Deep Frying Using Various Vegetable Oils on Acrylamide Formation in Sweet Potato (Ipomoea batatas L. Lam) Chips. J. Food Sci. 2014, 79, T115–T121. [Google Scholar] [CrossRef]
- Barison, A.; da Silva, C.W.P.; Campos, F.R.; Simonelli, F.; Lenz, C.A.; Ferreira, A.G. A simple methodology for the determination of fatty acid composition in edible oils through H-1 NMR spectroscopy. Magn. Reson. Chem. 2010, 48, 642–650. [Google Scholar] [CrossRef]
- Li, D.; Hu, X. Fatty Acid Content of Commonly Available Nuts and Seeds. In Nuts and Seeds in Health and Disease Prevention, 1st ed.; Preedy, V.R., Watson, R.R., Patel, V.B., Eds.; Academic Press: Cambridge, MA, USA, 2011; pp. 35–42. [Google Scholar]
- Shiozawa, S.; Tanaka, M.; Ohno, K.; Nagao, Y.; Yamada, T. Re-evaluation of peroxide value as an indicator of the quality of edible oils. J. Food Hyg. Soc. Jpn. 2007, 48, 51–57. [Google Scholar] [CrossRef]
- Kanazawa, K.; Ashida, H. Dietary hydroperoxides of linoleic acid decompose to aldehydes in stomach before being absorbed into the body. Biochim. Biophys. Acta 1998, 1393, 349–361. [Google Scholar] [CrossRef]
- Billek, G. Health aspects of thermoxidized oils and fats. Eur. J. Lipid Sci. Technol. 2000, 102, 587–593. [Google Scholar] [CrossRef]
- Ghidurus, M.; Turtoi, M.; Boskou, G.; Niculita, P.; Stan, V. Nutritional and health aspects related to frying (II). Rom. Biotechnol. Lett. 2011, 16, 6467–6472. [Google Scholar]
- Nourooz-Zadeh, J.; Tajaddini-Sarmadi, J.; Ling, K.L.E.; Wolff, S.P. Low-density lipoprotein is the major carrier of lipid hydroperoxides in plasma - Relevance to determination of total plasma lipid hydroperoxide concentrations. Biochem. J. 1996, 313, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Ringseis, R.; Eder, K. Regulation of genes involved in lipid metabolism by dietary oxidized fat. Mol. Nutr. Food Res. 2011, 55, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, K.; Ashida, H. Catabolic fate of dietary trilinoleoylglycerol hydroperoxides in rat gastrointestines. Biochim. Biophys. Acta 1998, 1393, 336–348. [Google Scholar] [CrossRef]
- Kanner, J.; Lapidot, T. The stomach as a bioreactor: Dietary lipid peroxidation in the gastric fluid and the effects of plant-derived antioxidants. Free Radic. Biol. Med. 2001, 31, 1388–1395. [Google Scholar] [CrossRef]
- Sack, M.; Hölz, K.; Holik, A.K.; Kretschy, N.; Somoza, V.; Stengele, K.P.; Somoza, M.M. Express photolithographic DNA microarray synthesis with optimized chemistry and high-efficiency photolabile groups. J. Nanobiotechnol. 2016, 14, 14. [Google Scholar] [CrossRef]
- Grigoryev, D.N.; Ma, S.-F.; Irizarry, R.A.; Ye, S.Q.; Quackenbush, J.; Garcia, J.G.N. Orthologous gene-expression profiling in multi-species models: Search for candidate genes. Genome Biol. 2004, 5, R34. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef]
- Pignitter, M.; Stolze, K.; Gartner, S.; Dumhart, B.; Stoll, C.; Steiger, G.; Kraemer, K.; Somoza, V. Cold fluorescent light as major inducer of lipid oxidation in soybean oil stored at household conditions for eight weeks. J. Agric. Food Chem. 2014, 62, 2297–2305. [Google Scholar] [CrossRef]
- Benini, L.; Brighenti, F.; Castellani, G.; Brentegani, M.T.; Casiraghi, M.C.; Ruzzenente, O.; Sembenini, C.; Pellegrini, N.; Caliari, S.; Porrini, M.; et al. Gastric-Emptying of Solids Is Markedly Delayed When Meals Are Fried. Dig. Dis. Sci. 1994, 39, 2288–2294. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Singh, R.P. Disintegration of solid foods in human stomach. J. Food Sci. 2008, 73, R67–R80. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.H.; Mayer, E.A.; Jehn, D.; Gu, Y.; Fink, A.S.; Fried, M. Gastric Processing and Emptying of Fat. Gastroenterology 1986, 90, 1176–1187. [Google Scholar] [CrossRef]
- Wingler, K.; Muller, C.; Schmehl, K.; Florian, S.; Brigelius-Flohe, R. Gastrointestinal glutathione peroxidase prevents transport of lipid hydroperoxides in CaCo-2 cells. Gastroenterology 2000, 119, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Schieberle, P.; Grosch, W. Decomposition of linoleic acid hydroperoxides. II. Breakdown of Methyl 13-hydroperoxy-cis-9-trans-11-octadecadienoate by Radicals or Copper-II Ions. Z. Lebensm. Unters. 1981, 173, 192–198. [Google Scholar] [CrossRef]
- Ramsden, C.E.; Ringel, A.; Feldstein, A.E.; Taha, A.Y.; MacIntosh, B.A.; Hibbeln, J.R.; Majchrzak-Hong, S.F.; Faurot, K.R.; Rapoport, S.I.; Cheon, Y.; et al. Lowering dietary linoleic acid reduces bioactive oxidized linoleic acid metabolites in humans. Prostaglandins Leukot. Essent. Fat. Acids 2012, 87, 135–141. [Google Scholar] [CrossRef]
- Kanazawa, K.; Kanazawa, E.; Natake, M. Uptake of Secondary Autoxidation Products of Linoleic-Acid by the Rat. Lipids 1985, 20, 412–419. [Google Scholar] [CrossRef]
- Feldstein, A.E.; Lopez, R.; Tamimi, T.A.R.; Yerian, L.; Chung, Y.M.; Berk, M.; Zhang, R.L.; McIntyre, T.M.; Hazen, S.L. Mass spectrometric profiling of oxidized lipid products in human nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. J. Lipid Res. 2010, 51, 3046–3054. [Google Scholar] [CrossRef]
- Yoshida, Y.; Umeno, A.; Akazawa, Y.; Shichiri, M.; Murotomi, K.; Hone, M. Chemistry of Lipid Peroxidation Products and Their Use as Biomarkers in Early Detection of Diseases. J. Oleo Sci. 2015, 64, 347–356. [Google Scholar] [CrossRef]
- Guadagni, R.; Miraglia, N.; Simonelli, A.; Silvestre, A.; Lamberti, M.; Feola, D.; Acampora, A.; Sannolo, N. Solid-phase microextraction-gas chromatography-mass spectrometry method validation for the determination of endogenous substances: Urinary hexanal and heptanal as lung tumor biomarkers. Anal. Chim. Acta 2011, 701, 29–36. [Google Scholar] [CrossRef]
- Li, J.; Peng, Y.L.; Liu, Y.; Li, W.W.; Jin, Y.; Tang, Z.T.; Duan, Y.X. Investigation of potential breath biomarkers for the early diagnosis of breast cancer using gas chromatography-mass spectrometry. Clin. Chim. Acta 2014, 436, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Paushkin, S.V.; Patel, M.; Furia, B.S.; Peltz, S.W.; Trotta, C.R. Identification of a Human Endonuclease Complex Reveals a Link between tRNA Splicing and Pre-mRNA 3′ End Formation. Cell 2004, 117, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Monirujjaman, M.; Ferdouse, A. Metabolic and physiological roles of branched-chain amino acids. Adv. Mol. Biol. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Yoshizawa, F. Regulation of protein synthesis by branched-chain amino acids in vivo. Biochem. Biophys. Res. Commun. 2004, 313, 417–422. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Xu, W.T.; Tang, R.; Li, L.; Refaey, M.M.; Li, D.P. Thermally processed diet greatly affects profiles of amino acids rather than fatty acids in the muscle of carnivorous Silurus meridionalis. Food Chem. 2018, 256, 244–251. [Google Scholar] [CrossRef]
- Jiao, J.; Han, S.F.; Zhang, W.; Xu, J.Y.; Tong, X.; Yin, X.B.; Yuan, L.X.; Qin, L.Q. Chronic leucine supplementation improves lipid metabolism in C57BL/6J mice fed with a high-fat/cholesterol diet. Food Nut. Res. 2016, 60, 31304. [Google Scholar] [CrossRef] [Green Version]
- Yao, K.; Duan, Y.; Li, F.; Tan, B.; Hou, Y.; Wu, G.; Yin, Y. Leucine in Obesity: Therapeutic Prospects. Trends Pharmacol. Sci. 2016, 37, 714–727. [Google Scholar] [CrossRef]
- Pignitter, M.; Zaunschirm, M.; Lach, J.; Unterberger, L.; Kopic, A.; Kessler, C.; Kienesberger, J.; Pischetsrieder, M.; Eggersdorfer, M.; Riegger, C.; et al. Regioisomeric distribution of 9- and 13-hydroperoxy linoleic acid in vegetable oils during storage and heating. J. Sci. Food Agric. 2017, 98, 1240–1247. [Google Scholar] [CrossRef] [Green Version]
- Laboisse, C.L.; Augeron, C.; Couturier-Turpin, M.H.; Gespach, C.; Cheret, A.M.; Potet, F. Characterization of a newly established human gastric cancer cell line HGT-1 bearing histamine H2-receptors. Cancer Res. 1982, 42, 1541–1548. [Google Scholar]
- Liszt, K.; Ley, J.P.; Lieder, B.; Behrens, M.; Stöger, V.; Reiner, A.; Hochkogler, C.M.; Köck, E.; Marchiori, A.; Hans, J.; et al. Caffeine induces gastric acid secretion via bitter taste signaling in gastric parietal cells. Proc. Natl. Acad. Sci. USA 2017, 114, E6260–E6269. [Google Scholar] [CrossRef] [Green Version]
- Maznah, I., Jr. The use of Caco-2 cells as an in vitro method to study bioavailability of iron. Malays. J. Nutr. 1999, 5, 31–45. [Google Scholar] [PubMed]
- Procida, G.; Cichelli, A.; Lagazio, C.; Conte, L.S. Relationships between volatile compounds and sensory characteristics in virgin olive oil by analytical and chemometric approaches. J. Sci. Food Agric. 2015, 96, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Romero, I.; Garcia-Gonzalez, D.L.; Aparicio-Ruiz, R.; Morales, M.T. Validation of SPME-GCMS method for the analysis of virgin olive oil volatiles responsible for sensory defects. Talanta 2015, 134, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.J.; Li, X.Q.; Shoemaker, C.F.; Wang, S.C. Ultrahigh Performance Liquid Chromatography Analysis of Volatile Carbonyl Compounds in Virgin Olive Oils. J. Agric. Food Chem. 2013, 61, 12253–12259. [Google Scholar] [CrossRef] [PubMed]
- Dufour, C.; Loonis, M. Regio- and stereoselective oxidation of linoleic acid bound to serum albumin: Identification by ESI-mass spectrometry and NMR of the oxidation products. Chem. Phys. Lipids 2005, 138, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.; Haihong, W.; Zhiming, G.; Chong, S.; Shuang, R.; Daoying, W.; Muhan, Z.; Fang, L.; Weimin, X. Simultaneous Determination of 13-HODE, 9,10-DHODE, and 9,10,13-THODE in Cured Meat Products by LC-MS/MS. Food Anal. Methods 2016, 9, 2832–2841. [Google Scholar] [CrossRef]
- Holik, A.K.; Rohm, B.; Somoza, M.M.; Somoza, V. N(ε)-Carboxymethyllysine (CML), a Maillard reaction product, stimulates serotonin release and activates the receptor for advanced glycation end products (RAGE) in SH-SY5Y cells. Food Funct. 2013, 4, 1111–1120. [Google Scholar] [CrossRef]
- Ouellet, M.; Adams, P.D.; Keasling, J.D.; Mukhopadhyay, A. A rapid and inexpensive labeling method for microarray gene expression analysis. BMC Biotechnol. 2009, 9. [Google Scholar] [CrossRef] [Green Version]
- Hochkogler, C.M.; Lieder, B.; Rust, P.; Berry, D.; Meier, S.M.; Pignitter, M.; Riva, A.; Leitinger, A.; Bruk, A.; Wagner, S.; et al. 12-week intervention with nonivamide, a TRPV1 agonist, prevents a dietary-induced body fat gain and increases peripheral serotonin in moderately overweight subjects. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Schieberle, P. The carbon module labeling (CAMOLA) technique: A useful tool for identifying transient intermediates in the formation of maillard-type target molecules. Ann. Ny Acad. Sci. 2005, 1043, 236–248. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Incubation Substance | Linoleic Acid | 13-HpODE | Hexanal |
|---|---|---|---|
| Quantitation of | 13-HODE [µM] | 13-HODE [µM] | Hexanal [µM] |
| apical | 1.11 ± 0.05 a | 1.22 ± 0.05 a | 3.15 ± 0.62 b |
| lysate | n.d. | n.d. | n.d. |
| basolateral | 1.12 ± 0.05 a | 2.09 ± 0.53 b,* | 0.20 ± 0.13 c,* |
| Compound | Pathway | Hits (Official Gene Symbol, KEGG Compound Entry) | p Value | Topology |
|---|---|---|---|---|
| Linoleic acid | Pyrimidine metabolism | UMPS, POLR1A, CANT1, NTSC3, TXNRD2, TYMP, TYMS | 0.036 | 0.37 |
| Cyanoamino metabolism | 5HMT1, C00037 | 0.040 | 0.75 | |
| 13-HpODE | Aminoacyl-tRNA biosynthesis | C00123, C00037, RARS2, C00148, FARSB, C00073 | < 0.001 | 0.12 |
| Arginine and proline metabolism | GLS2, ASS1, C00148, GOT2, ALDH3A2 | 0.004 | 0.17 | |
| Linoleic acid metabolism | C00157, PLA2G6, CYP2C8 | 0.006 | 0.88 | |
| Hexanal | Valine, leucine and isoleucine biosynthesis | PDHA1, C00183, C00123 | < 0.001 | 0.45 |
| Aminoacyl-tRNA biosynthesis | C00183, C00123, C00037, C00062, C00073 | < 0.001 | 0.07 | |
| Pantothenate and CoA biosynthesis | VNN2, C00183 | 0.030 | 0.13 | |
| Arginine and proline metabolism | NOS2, C00062, C00077 | 0.044 | 0.27 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaunschirm, M.; Pignitter, M.; Kopic, A.; Keßler, C.; Hochkogler, C.; Kretschy, N.; Somoza, M.M.; Somoza, V. Exposure of Human Gastric Cells to Oxidized Lipids Stimulates Pathways of Amino Acid Biosynthesis on a Genomic and Metabolomic Level. Molecules 2019, 24, 4111. https://doi.org/10.3390/molecules24224111
Zaunschirm M, Pignitter M, Kopic A, Keßler C, Hochkogler C, Kretschy N, Somoza MM, Somoza V. Exposure of Human Gastric Cells to Oxidized Lipids Stimulates Pathways of Amino Acid Biosynthesis on a Genomic and Metabolomic Level. Molecules. 2019; 24(22):4111. https://doi.org/10.3390/molecules24224111
Chicago/Turabian StyleZaunschirm, Mathias, Marc Pignitter, Antonio Kopic, Claudia Keßler, Christina Hochkogler, Nicole Kretschy, Mark Manuel Somoza, and Veronika Somoza. 2019. "Exposure of Human Gastric Cells to Oxidized Lipids Stimulates Pathways of Amino Acid Biosynthesis on a Genomic and Metabolomic Level" Molecules 24, no. 22: 4111. https://doi.org/10.3390/molecules24224111