Assembly and Analysis of the Genome Sequence of the Yeast Brettanomyces naardenensis CBS 7540

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation
2.2. Genome Sequencing
2.3. RNA Library Preparation and Sequencing
2.4. Genome Assembly
2.5. Transcriptome-Guided Genome Annotation
2.6. Functional Genome Annotation
2.7. SNP Analysis
2.8. Phylogenetic Tree Construction
2.9. Comparative Analysis of Gene Content
3. Results
3.1. Genome Structure
3.2. Genome Annotation
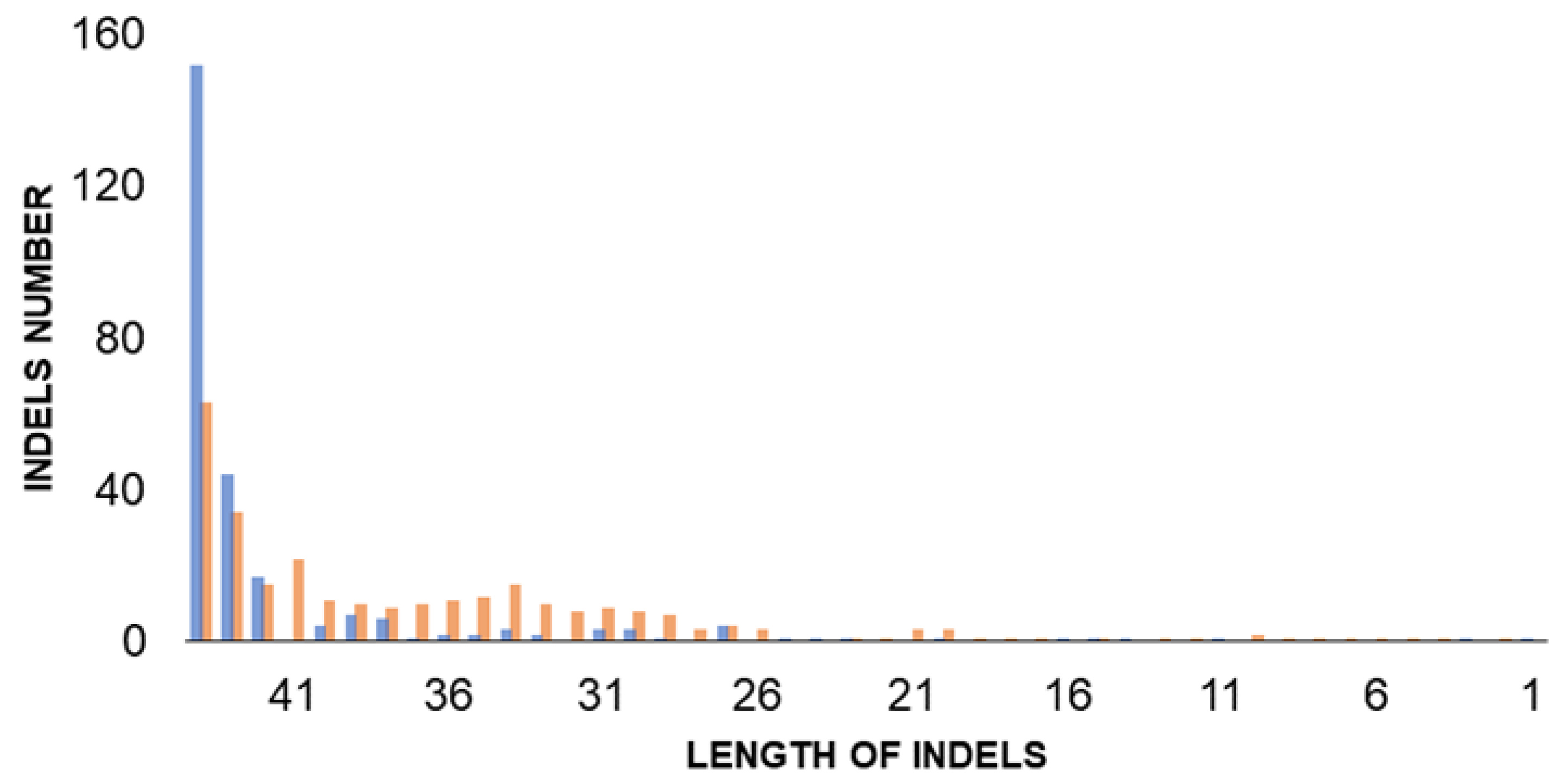
3.3. Heterozygosity
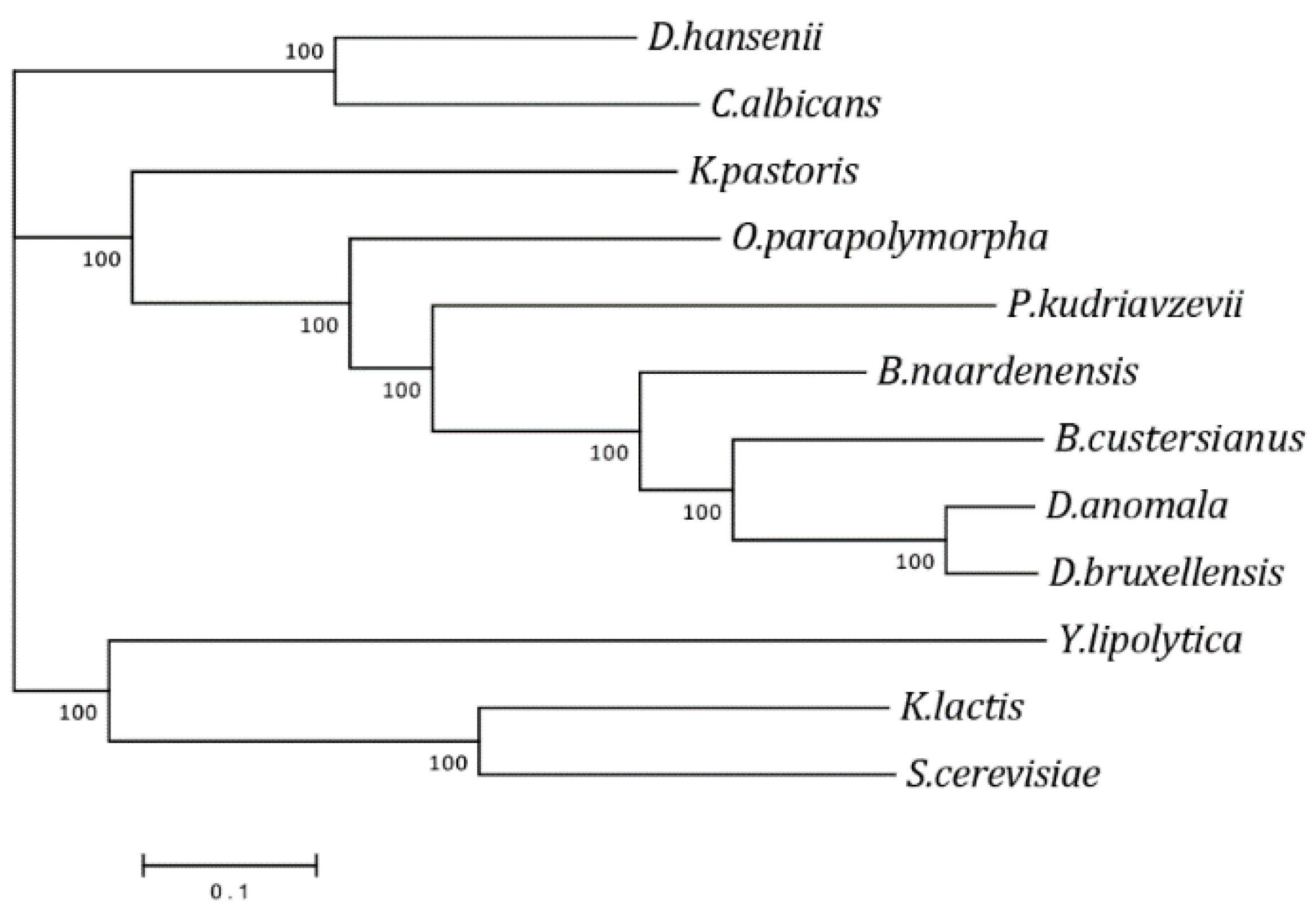
3.4. Phylogenetic Analysis
3.5. Genes Associated with Food-Related Traits of B. Naardenensis CBS 7540
3.5.1. Assimilatory Pathways
3.5.2. Genes Putatively Involved in Production of Volatiles
3.5.3. Putative Stress Tolerance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hawksworth, D.L.; Crous, P.W.; Redhead, S.A.; Reynolds, D.R.; Samson, R.A.; Seifert, K.A.; Taylor, J.W.; Wingfield, M.J.; Abaci, Ö.; Aime, C.; et al. The Amsterdam Declaration on Fungal Nomenclature. Ima. Fungus. 2011, 2, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.D.; Divol, B. Brettanomyces bruxellensis, a Survivalist Prepared for the Wine Apocalypse and Other Beverages. Food Microbiol. 2016, 59, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, J.; Passoth, V. Dekkera bruxellensis―Spoilage Yeast with Biotechnological Potential, and a Model for Yeast Evolution, Physiology and Competitiveness. FEMS Yeast Res. 2015, 15, fov021. [Google Scholar] [CrossRef] [PubMed]
- Schifferdecker, A.J.; Dashko, S.; Ishchuk, O.P.; Piškur, J. The Wine and Beer Yeast Dekkera bruxellensis. Yeast 2014, 31, 323–332. [Google Scholar] [CrossRef]
- Rozpedowska, E.; Hellborg, L.; Ishchuk, O.P.; Orhan, F.; Galafassi, S.; Merico, A.; Woolfit, M.; Compagno, C.; Piškur, J. Parallel Evolution of the Make-Accumulate-Consume Strategy in Saccharomyces and Dekkera Yeasts. Nat. Commun. 2011, 2, 302. [Google Scholar] [CrossRef]
- Steensels, J.; Daenen, L.; Malcorps, P.; Derdelinckx, G.; Verachtert, H.; Verstrepen, K.J. Brettanomyces Yeasts - From Spoilage Organisms to Valuable Contributors to Industrial Fermentations. Int. J. Food Microbiol. 2015, 206, 24–38. [Google Scholar] [CrossRef]
- Kolfschoten, G.A.; Yarrow, D. Brettanomyces naardenensis, a New Yeast from Soft Drinks. Antonie Leeuwenhoek 1970, 36, 458–460. [Google Scholar] [CrossRef]
- Wareing, P.; Davenport, R.R. Microbiology of Soft Drinks and Fruit Juices. In Chemistry and Technology of Soft Drinks and Fruit Juices, 2nd ed.; Blackwell Publishing Ltd.: Oxford, UK, 2007. [Google Scholar]
- Shi, X.; Chen, F.; Xu, Y.; Zheng, X.; Xiao, J. Aromatic Components Produced by Non-Saccharomyces cerevisiae Derived from Natural Fermentation of Grape. Nat. Prod. Res. 2015, 29, 1870–1873. [Google Scholar] [CrossRef]
- Galafassi, S.; Merico, A.; Pizza, F.; Hellborg, L.; Molinari, F.; Piškur, J.; Compagno, C. Dekkera/Brettanomyces Yeasts for Ethanol Production from Renewable Sources under Oxygen-Limited and Low-PH Conditions. J. Ind. Microbiol. Biotechnol. 2011, 38, 1079–1088. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dörnberg, T.; Jacob, F.; Methner, F.J.; Wagner, R.S.; Hutzler, M. Review: Pure Non-Saccharomyces Starter Cultures for Beer Fermentation with a Focus on Secondary Metabolites and Practical Applications. J. Inst. Brew. 2016, 122, 569–587. [Google Scholar] [CrossRef]
- De Barros Pita, W.; Leite, F.C.B.; De Souza Liberal, A.T.; Simões, D.A.; De Morais, M.A. The Ability to Use Nitrate Confers Advantage to Dekkera bruxellensis over S. cerevisiae and Can Explain Its Adaptation to Industrial Fermentation Processes. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2011, 100, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.H.; Jones, R.R.; Brender, J.D.; de Kok, T.M.; Weyer, P.J.; Nolan, B.T.; Villanueva, C.M.; van Breda, S.G. Drinking Water Nitrate and Human Health: An Updated Review. Int. J. Environ. Res. Public Health 2018, 15, 1557. [Google Scholar] [CrossRef] [PubMed]
- Joseph, C.M.L.; Albino, E.; Bisson, L.F. Creation and Use of a Brettanomyces Aroma Wheel. Catal. Discov. Pract. 2017, 1, 12–20. [Google Scholar] [CrossRef]
- Serra Colomer, M.; Funch, B.; Forster, J. The Raise of Brettanomyces Yeast Species for Beer Production. Curr. Opin. Biotechnol. 2019, 56, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Daenen, L.; Sterckx, F.; Delvaux, F.R.; Verachtert, H.; Derdelinckx, G. Evaluation of the Glycoside Hydrolase Activity of a Brettanomyces Strain on Glycosides from Sour Cherry (Prunus Cerasus L.) Used in the Production of Special Fruit Beers. FEMS Yeast Res. 2008, 8, 1103–1114. [Google Scholar] [CrossRef]
- Takoi, D.K. Varietal Difference of Hop- Derived Flavour Compounds in Dry-Hopped Beers. Brauwelt Int. 2016, 69, 1–7. [Google Scholar]
- Kuo, H.P.; Wang, R.; Huang, C.Y.; Lai, J.T.; Lo, Y.C.; Huang, S.T. Characterization of an Extracellular β-Glucosidase from Dekkera bruxellensis for Resveratrol Production. J. Food Drug Anal. 2018, 26, 163–171. [Google Scholar] [CrossRef]
- Borneman, A.R.; Desany, B.A.; Riches, D.; Affourtit, J.P.; Forgan, A.H.; Pretorius, I.S.; Egholm, M.; Chambers, P.J. The Genome Sequence of the Wine Yeast VIN7 Reveals an Allotriploid Hybrid Genome with Saccharomyces cerevisiae and Saccharomyces kudriavzevii Origins. FEMS Yeast Res. 2012, 12, 88–96. [Google Scholar] [CrossRef]
- Crauwels, S.; Van Assche, A.; de Jonge, R.; Borneman, A.R.; Verreth, C.; Troels, P.; De Samblanx, G.; Marchal, K.; Van de Peer, Y.; Willems, K.A.; et al. Comparative Phenomics and Targeted Use of Genomics Reveals Variation in Carbon and Nitrogen Assimilation among Different Brettanomyces bruxellensis Strains. Appl. Microbiol. Biotechnol. 2015, 99, 9123–9134. [Google Scholar] [CrossRef]
- Crauwels, S.; Zhu, B.; Steensels, J.; Busschaert, P.; De Samblanx, G.; Marchal, K.; Willems, K.A.; Verstrepen, K.J.; Lievens, B. Assessing Genetic Diversity among Brettanomyces Yeasts by DNA Fingerprinting and Whole-Genome Sequencing. Appl. Environ. Microbiol. 2014, 80, 4398–4413. [Google Scholar] [CrossRef] [Green Version]
- Curtin, C.D.; Borneman, A.R.; Chambers, P.J.; Pretorius, I.S. De-Novo Assembly and Analysis of the Heterozygous Triploid Genome of the Wine Spoilage Yeast Dekkera bruxellensis AWRI1499. PLoS ONE 2012, 8, e33840. [Google Scholar] [CrossRef] [PubMed]
- Curtin, C.D.; Pretorius, I.S. Genomic Insights into the Evolution of Industrial Yeast Species Brettanomyces bruxellensis. FEMS Yeast Res. 2014, 14, 997–1005. [Google Scholar] [PubMed]
- Fournier, T.; Gounot, J.S.; Freel, K.; Cruaud, C.; Lemainque, A.; Aury, J.M.; Wincker, P.; Schacherer, J.; Friedrich, A. High-Quality de Novo Genome Assembly of the Dekkera bruxellensis Yeast Using Nanopore MinION Sequencing. Genes Genomes. Genet. 2017, 7, 3243–3250. [Google Scholar] [CrossRef] [PubMed]
- Piškur, J.; Ling, Z.; Marcet-Houben, M.; Ishchuk, O.P.; Aerts, A.; LaButti, K.; Copeland, A.; Lindquist, E.; Barry, K.; Compagno, C.; et al. The Genome of Wine Yeast Dekkera bruxellensis Provides a Tool to Explore Its Food-Related Properties. Int. J. Food Microbiol. 2012, 157, 202–209. [Google Scholar] [CrossRef]
- Valdes, J.; Tapia, P.; Cepeda, V.; Varela, J.; Godoy, L.; Cubillos, F.A.; Silva, E.; Martinez, C.; Ganga, M.A. Draft Genome Sequence and Transcriptome Analysis of the Wine Spoilage Yeast Dekkera bruxellensis LAMAP2480 Provides Insights into Genetic Diversity, Metabolism and Survival. Fems Microbiol. Lett. 2014, 361, 104–106. [Google Scholar] [CrossRef]
- Tiukova, I.A.; Pettersson, M.E.; Hoeppner, M.P.; Olsen, R.A.; Käller, M.; Nielsen, J.; Dainat, J.; Lantz, H.; Söderberg, J.; Passoth, V. Chromosomal Genome Assembly of the Ethanol Production Strain CBS 11270 Indicates a Highly Dynamic Genome Structure in the Yeast Species Brettanomyces bruxellensis. PLoS ONE 2019, 14, e0215077. [Google Scholar] [CrossRef]
- Shen, X.X.; Opulente, D.A.; Kominek, J.; Zhou, X.; Steenwyk, J.L.; Buh, K.V.; Haase, M.A.B.; Wisecaver, J.H.; Wang, M.; Doering, D.T.; et al. Tempo and Mode of Genome Evolution in the Budding Yeast Subphylum. Cell 2018, 175, 1533–1545. [Google Scholar] [CrossRef]
- Zhou, N.; Swamy, K.B.S.; Leu, J.Y.; McDonald, M.J.; Galafassi, S.; Compagno, C.; Piškur, J. Coevolution with Bacteria Drives the Evolution of Aerobic Fermentation in Lachancea kluyveri. PLoS ONE 2017, 12, e0173318. [Google Scholar] [CrossRef]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y. SOAPdenovo2: An Empirically Improved Memory-Efficient Short-Read de Novo Assembler. Giga Sci. 2012, 1, 2047. [Google Scholar] [CrossRef]
- Gu, S.; Fang, L.; Xu, X. Using SOAPaligner for Short Reads Alignment. Curr. Protoc. Bioinform. 2013, 44, 1–17. [Google Scholar]
- Norling, M.; Jareborg, N.; Dainat, J. EMBLmyGFF3: A Converter Facilitating Genome Annotation Submission to European Nucleotide Archive. BMC Res. Notes 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- H, L.; R, D. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar]
- Olsen, R.A.; Bunikis, I.; Tiukova, I.; Holmberg, K.; Lötstedt, B.; Pettersson, O.V.; Passoth, V.; Käller, M.; Vezzi, F. De Novo Assembly of Dekkera bruxellensis: A Multi Technology Approach Using Short and Long-Read Sequencing and Optical Mapping. Giga Sci. 2015, 4, s13742-015-0094-1. [Google Scholar] [CrossRef] [PubMed]
- Röder, C.; König, H.; Fröhlich, J. Species-Specific Identification of Dekkera/Brettanomyces Yeasts by Fluorescently Labeled DNA Probes Targeting the 26S RRNA. FEMS Yeast Res. 2007, 7, 1013–1026. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification and Phylogeny of Ascomycetous Yeasts from Analysis of Nuclear Large Subunit (26S) Ribosomal DNA Partial Sequences. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Toivola, A.; Yarrow, D.; Van Den Bosch, E. Alcoholic Fermentation of D-Xylose by Yeasts. Appl. Environ. Microbiol. 1984, 47, 1221–1223. [Google Scholar]
- De Barros Pita, W.; Tiukova, I.; Leite, F.C.B.; Passoth, V.; Simões, D.A.; De Morais, M.A. The Influence of Nitrate on the Physiology of the Yeast Dekkera bruxellensis Grown under Oxygen Limitation. Yeast 2013, 30, 111–117. [Google Scholar] [CrossRef]
- Woolfit, M.; Rozpȩdowska, E.; Piškur, J.; Wolfe, K.H. Genome Survey Sequencing of the Wine Spoilage Yeast Dekkera (Brettanomyces) bruxellensis. Eukaryot. Cell 2007, 6, 21–733. [Google Scholar] [CrossRef]
- Blomqvist, J.; Nogué, V.S.; Gorwa-Grauslund, M.; Passoth, V. Physiological Requirements for Growth and Competitiveness of Dekkera bruxellensis under Oxygen-Limited or Anaerobic Conditions. Yeast 2012, 29, 265–274. [Google Scholar] [CrossRef]
- Passoth, V.; Schäfer, B.; Liebel, B.; Weierstall, T.; Klinner, U. Molecular Cloning of Alcohol Dehydrogenase Genes of the Yeast Pichia stipitis and Identification of the Fermentative ADH. Yeast 1998, 14, 1311–1325. [Google Scholar] [CrossRef]
- Fredlund, E.; Beerlage, C.; Melin, P.; Schnürer, J.; Passoth, V. Oxygen and Carbon Source-Regulated Expression of PDC and ADH Genes in the Respiratory Yeast Pichia anomala. Yeast 2006, 23, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Romano, D.; Valdetara, F.; Zambelli, P.; Galafassi, S.; De Vitis, V.; Molinari, F.; Compagno, C.; Foschino, R.; Vigentini, I. Cloning the Putative Gene of Vinyl Phenol Reductase of Dekkera bruxellensis in Saccharomyces cerevisiae. Food Microbiol. 2017, 63, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Smith, M. Brettanomyces. In The Yeasts: A Taxonomic Study, 5th ed.; Elsevier: Oxford, UK, 2011. [Google Scholar]
- Silva, P.; Cardoso, H.; Gerós, H. Studies on the Wine Spoilage Capacity of Brettanomyces/Dekkera spp. Am. J. Enol. Vitic. 2004, 55, 65–72. [Google Scholar]
- Wijsman, M.R.; van Dijken, J.P.; van Kleeff, B.H.A.; Scheffers, W.A. Inhibition of Fermentation and Growth in Batch Cultures of the Yeast Brettanomyces intermedius upon a Shift from Aerobic to Anaerobic Conditions (Custers Effect). Antonie Leeuwenhoek 1984, 50, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Scheffers, W.A. Stimulation of Fermentation in Yeasts by Acetoin and Oxygen. Nature 1966, 210, 533–534. [Google Scholar] [CrossRef] [Green Version]
- Tiukova, I.A.; Petterson, M.E.; Tellgren-Roth, C.; Bunikis, I.; Eberhard, T.; Pettersson, O.V.; Passoth, V. Transcriptome of the Alternative Ethanol Production Strain Dekkera bruxellensis CBS 11270 in Sugar Limited, Low Oxygen Cultivation. PLoS ONE 2013, 8, e58455. [Google Scholar] [CrossRef]
- Russell, J.B. Another Explanation for the Toxicity of Fermentation Acids at Low PH: Anion Accumulation versus Uncoupling. J. Appl. Bacteriol. 1992, 73, 363–370. [Google Scholar] [CrossRef]
- Imai, T.; Ohno, T. The Relationship between Viability and Intracellular PH in the Yeast Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1995, 61, 3604–3608. [Google Scholar]
- Ludovico, P.; Sousa, M.J.; Silva, M.T.; Leão, C.; Côrte-Real, M. Saccharomyces cerevisiae Commits to a Programmed Cell Death Process in Response to Acetic Acid. Microbiology 2001, 147, 2409–2415. [Google Scholar] [CrossRef]
- Meijnen, J.P.; Randazzo, P.; Foulquié-Moreno, M.R.; Van Den Brink, J.; Vandecruys, P.; Stojiljkovic, M.; Dumortier, F.; Zalar, P.; Boekhout, T.; Gunde-Cimerman, N.; et al. Polygenic Analysis and Targeted Improvement of the Complex Trait of High Acetic Acid Tolerance in the Yeast Saccharomyces cerevisiae. Biotechnol. Biofuels 2016, 9, 5. [Google Scholar] [CrossRef]
- Passoth, V.; Blomqvist, J.; Schnürer, J. Dekkera bruxellensis and Lactobacillus vini Form a Stable Ethanol-Producing Consortium in a Commercial Alcohol Production Process. Appl. Env. Microbiol. 2007, 73, 354–4356. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.T.; Batenburg-van der Vegte, W.H.; Scheffers, W.A. Eeniella, a New Yeast Genus of the Torulopsidales. Int. J. Syst. Bacteriol. 1981, 31, 196–203. [Google Scholar] [CrossRef]
- Yamada, Y.; Matsuda, M.; Mikata, K. The Phylogenetic Relationships of Eeniella nana Smith, Batenburg-van Der Vegte et Scheffers Based on the Partial Sequences of 18S and 26S Ribosomal RNAs (Candidaceae). J. Ind. Microbiol. 1995, 14, 456–460. [Google Scholar] [CrossRef]
- Avramova, M.; Cibrario, A.; Peltier, E.; Coton, M.; Coton, E.; Schacherer, J.; Spano, G.; Capozzi, V.; Blaiotta, G.; Salin, F.; et al. Brettanomyces bruxellensis Population Survey Reveals a Diploid-Triploid Complex Structured According to Substrate of Isolation and Geographical Distribution. Sci. Rep. 2018, 8, 4136. [Google Scholar] [CrossRef] [PubMed]
- Shimotsu, S.; Asano, S.; Iijima, K.; Suzuki, K.; Yamagishi, H.; Aizawa, M. Investigation of Beer-Spoilage Ability of Dekkera/Brettanomyces Yeasts and Development of Multiplex PCR Method for Beer-Spoilage Yeasts. J. Inst. Brew. 2015, 121, 177–180. [Google Scholar] [CrossRef]
- Curtin, C.; Kennedy, E.; Henschke, P.A. Genotype-Dependent Sulphite Tolerance of Australian Dekkera (Brettanomyces) bruxellensis Wine Isolates. Lett. Appl. Microbiol. 2012, 55, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Jong, S.C.; Lee, F.L. The New Species Dekkera naardenensis, Teleomorph of Brettanomyces naardenensis. Mycotaxon 1986, 25, 147–152. [Google Scholar]
- Smith, M.T.; Yamazaki, M.; Poot, G.A. Dekkera, Brettanomyces and Eeniella: Electrophoretic Comparison of Enzymes and DNA–DNA Homology. Yeast 1990, 6, 299–310. [Google Scholar] [CrossRef]
- Legras, J.L.; Merdinoglu, D.; Cornuet, J.M.; Karst, F. Bread, Beer and Wine: Saccharomyces cerevisiae Diversity Reflects Human History. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef]
- Mortimer, R.K.; Romano, P.; Suzzi, G.; Polsinelli, M. Genome Renewal: A New Phenomenon Revealed from a Genetic Study of 43 Strains of Saccharomyces cerevisiae Derived from Natural Fermentation of Grape Musts. Yeast 1994, 10, 1543–1552. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Assembly Characteristic | Description |
|---|---|
| Number of contigs | 76 |
| Number of contigs >1 kb | 76 |
| Number of contigs >10 kb | 54 |
| N50 | 395,348 bp |
| N90 | 87,012 bp |
| Number of nucleotides | 11,283,072 bp |
| Number of Ns nucleotides | 752 bp |
| Number of N-regions | 45 |
| GC-content | 44.5% |
| Annotation Feature | Counts |
|---|---|
| Genes | 5168 |
| mRNAs | 5181 |
| Exons | 5692 |
| Introns | 496 |
| Mean intron frequency per gene | 0.1 |
| Mean intron length (bp) | 146 |
| Mean CDS length (bp) | 1488 |
| Mean mRNA length (bp) | 1528 |
| Total exon length (bp) | 7,840,339 |
| Total intron length (bp) | 72,570 |
| Variant Type | Counts |
|---|---|
| SNP | 323 |
| Indel | 533 |
| Total variant | 851 |
| SNP Type | Counts |
|---|---|
| A/C | 19 |
| A/G | 41 |
| T/G | 6 |
| T/A | 25 |
| C/T | 51 |
| T/C | 30 |
| C/A | 43 |
| G/T | 18 |
| A/T | 24 |
| C/G | 18 |
| G/C | 11 |
| G/A | 37 |
| Number of Variants | Gene Counts |
|---|---|
| 24 | 1 |
| 14 | 1 |
| 12 | 1 |
| 10 | 1 |
| 7 | 1 |
| 6 | 1 |
| 5 | 1 |
| 4 | 1 |
| 3 | 4 |
| 2 | 7 |
| 1 | 30 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiukova, I.A.; Jiang, H.; Dainat, J.; Hoeppner, M.P.; Lantz, H.; Piskur, J.; Sandgren, M.; Nielsen, J.; Gu, Z.; Passoth, V. Assembly and Analysis of the Genome Sequence of the Yeast Brettanomyces naardenensis CBS 7540. Microorganisms 2019, 7, 489. https://doi.org/10.3390/microorganisms7110489
Tiukova IA, Jiang H, Dainat J, Hoeppner MP, Lantz H, Piskur J, Sandgren M, Nielsen J, Gu Z, Passoth V. Assembly and Analysis of the Genome Sequence of the Yeast Brettanomyces naardenensis CBS 7540. Microorganisms. 2019; 7(11):489. https://doi.org/10.3390/microorganisms7110489
Chicago/Turabian StyleTiukova, Ievgeniia A., Huifeng Jiang, Jacques Dainat, Marc P. Hoeppner, Henrik Lantz, Jure Piskur, Mats Sandgren, Jens Nielsen, Zhenglong Gu, and Volkmar Passoth. 2019. "Assembly and Analysis of the Genome Sequence of the Yeast Brettanomyces naardenensis CBS 7540" Microorganisms 7, no. 11: 489. https://doi.org/10.3390/microorganisms7110489