Novel Bi-Factorial Strategy against Candida albicans Viability Using Carnosic Acid and Propolis: Synergistic Antifungal Action

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Culture Conditions
2.2. Natural Extracts
2.3. HPLC
2.4. Determination of Cell Viability
2.5. Morphological Analysis
2.6. Biofilm Formation
3. Results and Discussion
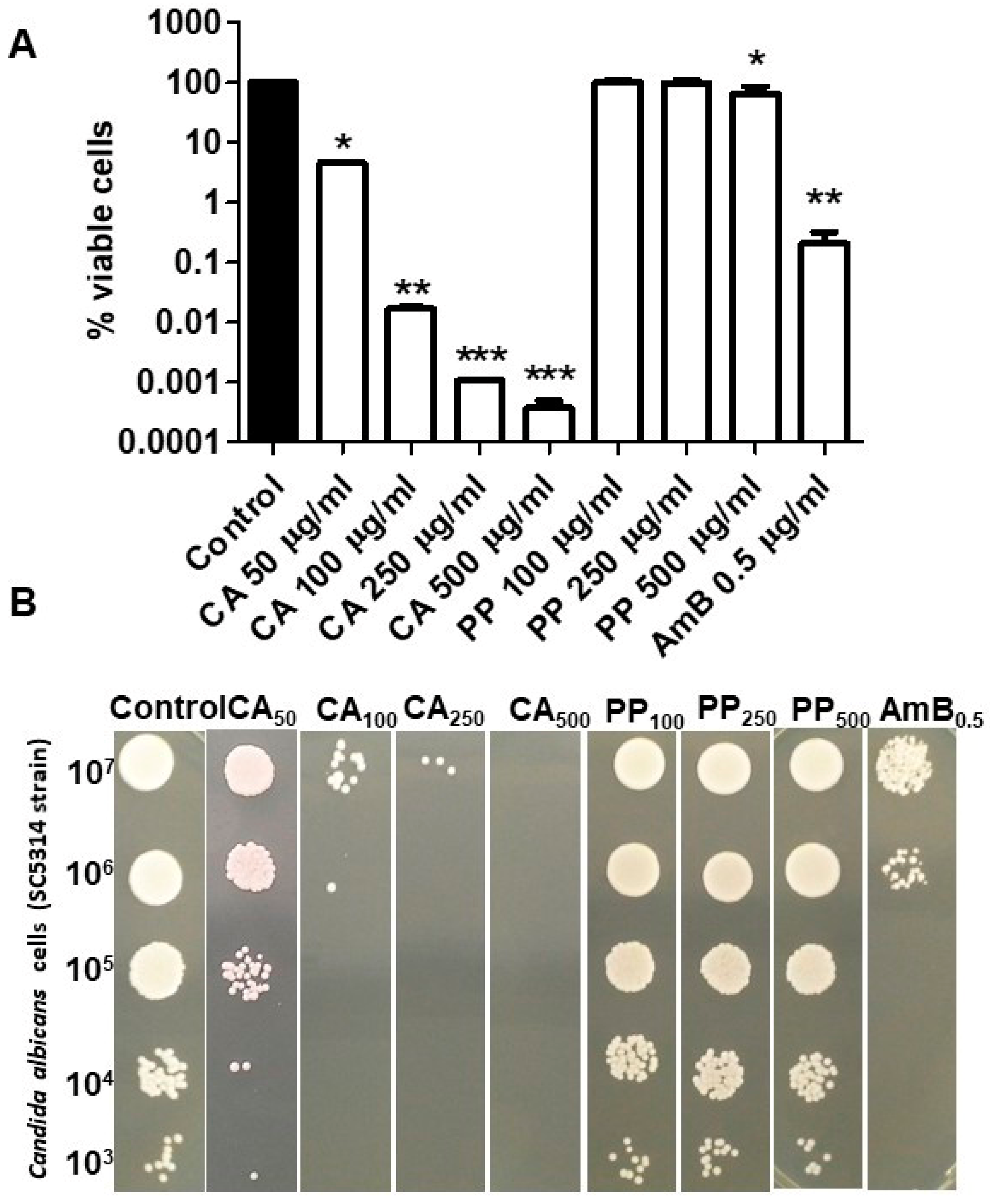
3.1. Single Antifungal Action of CA and PP on C. albicans
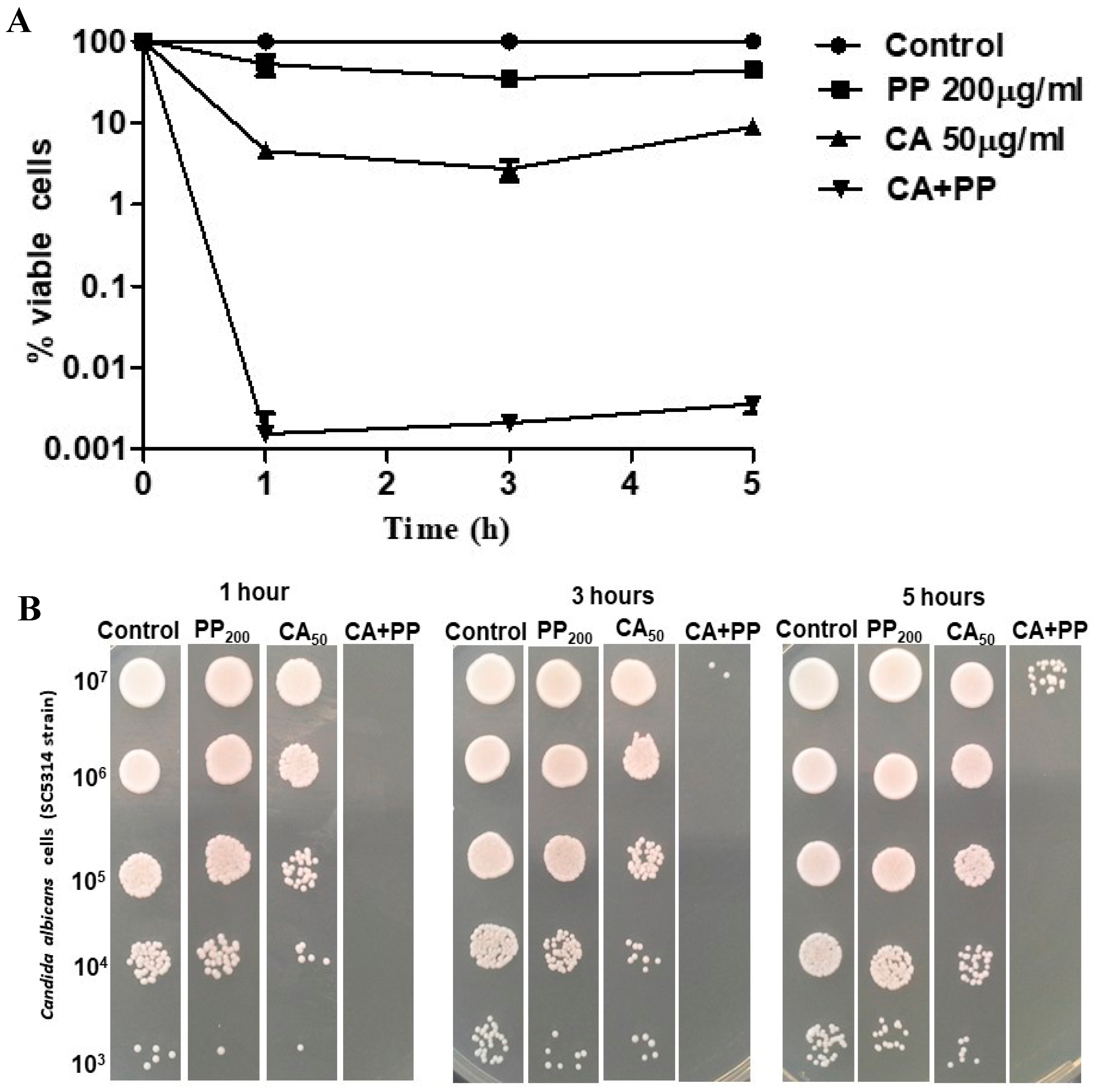
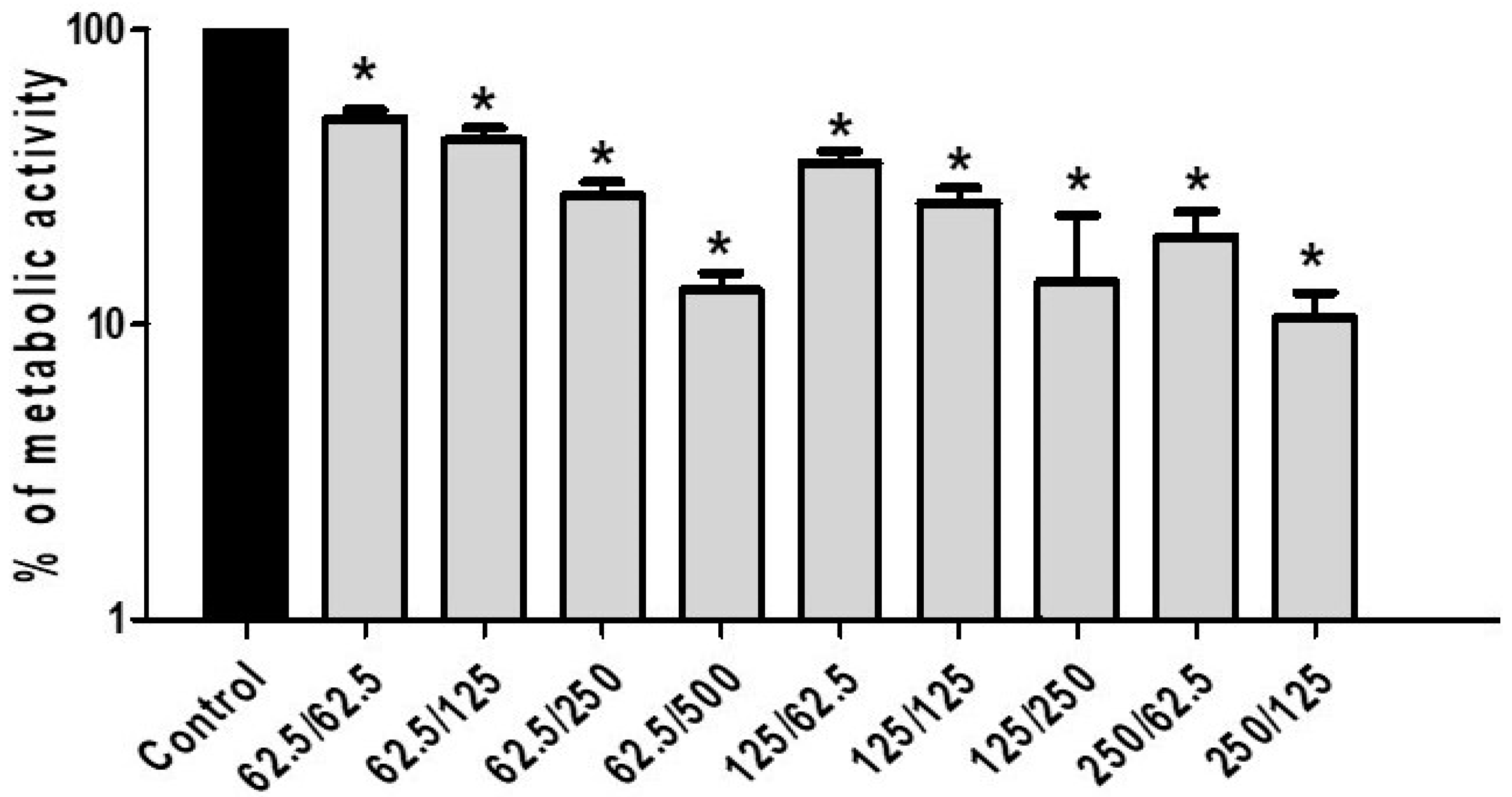
3.2. Antifungal Activity of CA and PP Mixtures on C. albicans
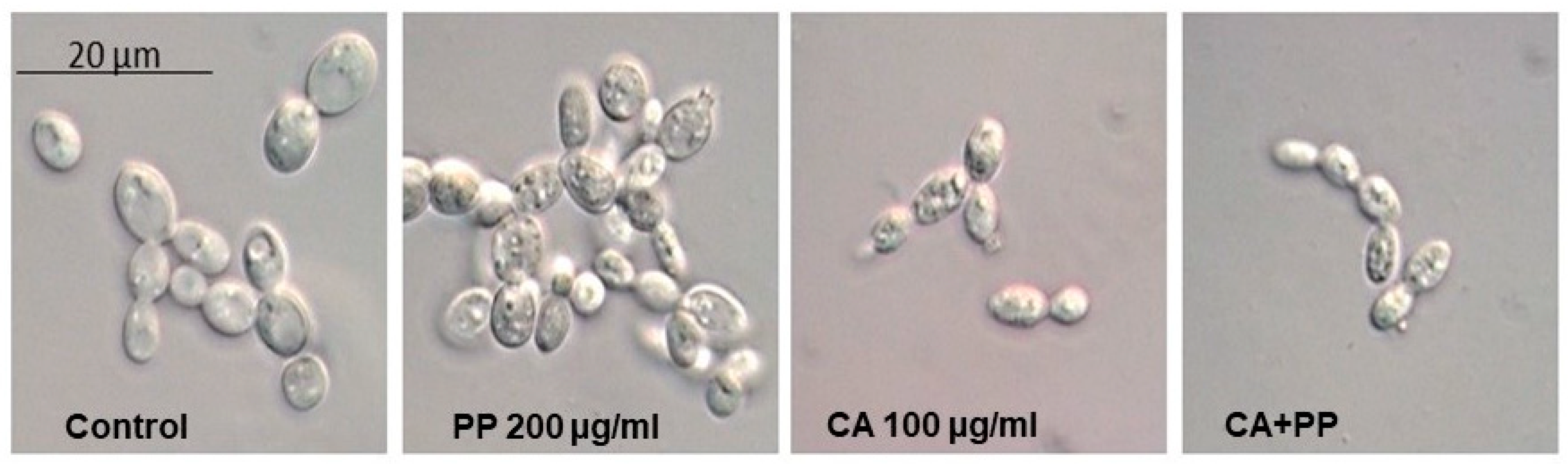
3.3. Morphological Changes Induced by Exposure to CA and PP
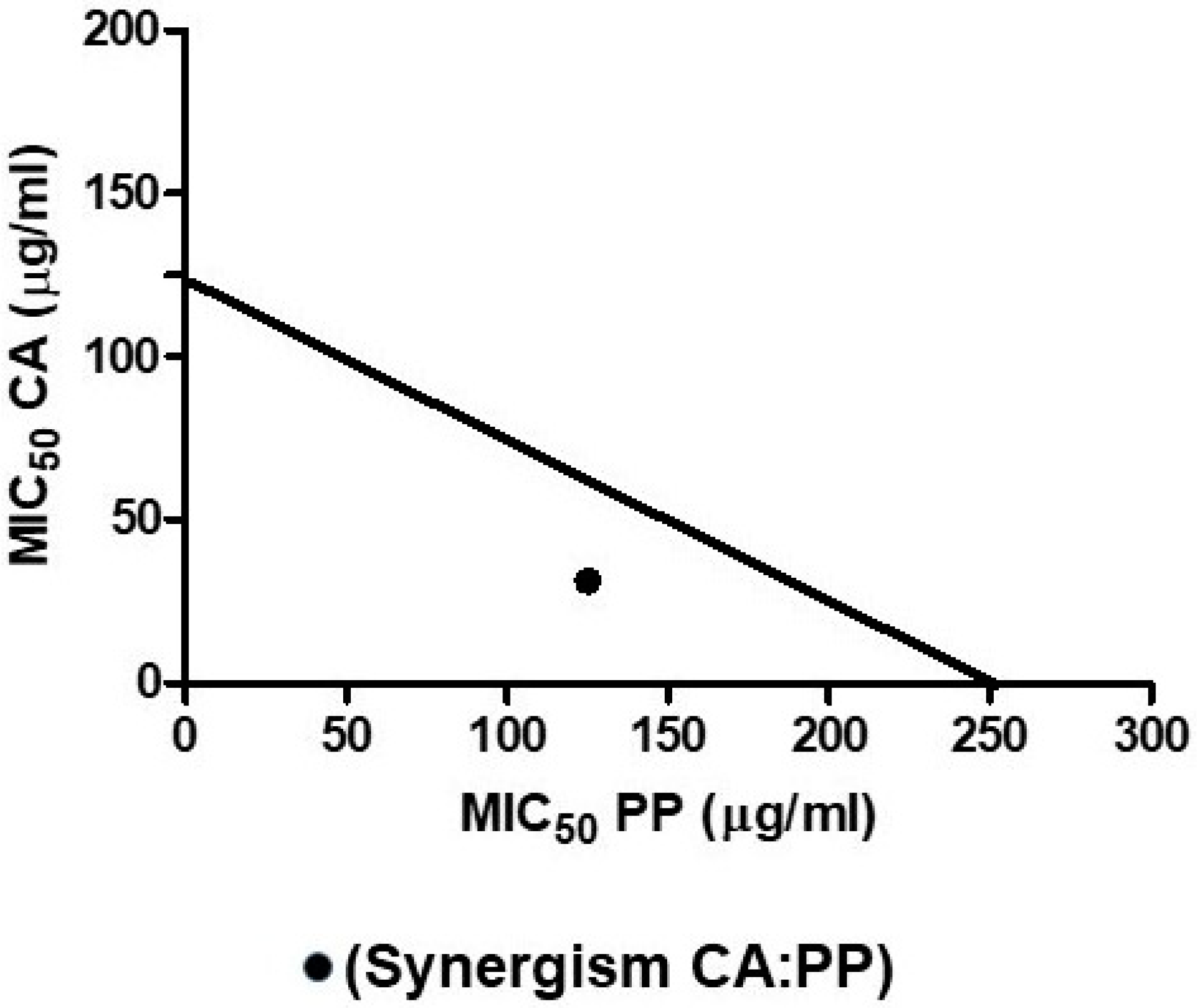
3.4. Isobologram Plot: Mathematical Demonstration of Synergism
3.5. The Capacity to Form Biofilms Is Impaired by the Addition of CA and PP
4. Conclusions
5. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bernardes, W.A.; Lucarini, R.; Tozatti, M.G.; Souza, M.G.; Andrade Silva, M.L.; da Silva Filho, A.A.; Martins, C.H.G.; Miller Crotti, A.E.; Pauletti, P.M.; Groppo, M.; et al. Antimicrobial activity of Rosmarinus officinalis against oral pathogens: Relevance of carnosic acid and carnosol. Chem. Biodiver. 2010, 7, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Birtić, S.; Dussort, P.; Pierre, F.-X.; Bily, A.C.; Roller, M. Carnosic acid. Phytochemistry 2015, 115, 9–19. [Google Scholar]
- Jordán, M.J.; Castillo, J.; Bañón, S.; Martínez-Conesa, C.; Sotomayor, J.A. Relevance of the carnosic acid/carnosol ratio for the level of rosemary diterpene transfer and for improving lamb meat antioxidant status. Food Chem. 2014, 151, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Bankova, V. Chemical diversity of propolis and the problem of standardization. J. Ethnopharm. 2005, 100, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.G.; Antunes, M.D. Is propolis safe as an alternative medicine? J. Pharm. Bioal. Sci. 2011, 3, 479. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, C.-P.; Wang, K.; Li, G.Q.; Hu, F.L. Recent advances in the chemical composition of Propolis. Molecules 2014, 19, 19610–19632. [Google Scholar] [CrossRef] [Green Version]
- Cigut, T.; Polak, T.Z.; Ga, L.; Raspor, P.; Jamnik, P. Antioxidative activity of Propolis extract in yeast cells. J. Agric. Food Chem. 2011, 59, 11449–11455. [Google Scholar] [CrossRef]
- De Castro, P.A.; Bom, V.L.; Brown, N.A.; de Almeida, R.S.C.; Ramalho, L.N.Z.; Savoldi, M.; Goldman, M.H.S.; Berretta, A.A.; Goldman, G.H. Identification of the cell targets important for propolis-induced cell death in Candida albicans. Fungal Genet. Biol. 2013, 60, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Sardi, J.C.; Giannini, M.J.; Bernardi, T.; Fusco-Almeida, A.M.; Giannini, M.M. Candida species: Current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. J. Med. Microbiol. 2013, 62, 10–24. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Jones, R.N.; Castanheira, M. Regional data analysis of Candida non-albicans strains collected in United States medical sites over a 6-year period, 2006–2011. Mycoses 2014, 57, 602–611. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Castanheira, M. Nosocomial candidiasis: Antifungal stewardship and the importance of rapid diagnosis. Med. Mycol. 2016, 54, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yapar, N. Epidemiology and risk factors for invasive candidiasis. Therap. Clin. Risk Manag. 2014, 10, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campoy, S.; Adrio, J.L. Antifungals. Biochem. Pharmacol. 2017, 133, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Guirao-Abad, J.P.; Sánchez-Fresneda, R.; Alburquerque, B.; Hernández, J.A.; Argüelles, J.C. ROS formation is a differential contributory factor to the fungicidal action of Amphotericin B and Micafungin in Candida albicans. Internatl. J. Med. Microbiol. 2017, 307, 241–248. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Tomic, G.; Nikolic, I.; Nerantzaki, A.A.; Sayyad, N.; Stosic-Grujicic, S.; Stojanovic, I.; Gerothanassis, I.P.; Tzakos, A.G. Phytochemical profile of Rosmarinus officinalis and Salvia officinalis extracts and correlation to their antioxidant and anti-proliferative activity. Food Chem. 2013, 136, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Erkan, N.; Ayranci, G.; Ayranci, E. Antioxidant activities of rosemary (Rosmarinus Officinalis L.) extract, blackseed (Nigella sativa L.) essential oil, carnosic acid, rosmarinic acid and sesamol. Food Chem. 2008, 110, 76–82. [Google Scholar] [CrossRef]
- Benavente-García, O.; Castillo, J.; Lorente, J.; Ortuño, A.D.R.J.; Del Rio, J.A. Antioxidant activity of phenolics extracted from Olea europaea L. leaves. Food Chem. 2000, 68, 457–462. [Google Scholar] [CrossRef]
- Badiee, P.; Reza Nasirzadeh, A.; Motaffaf, M. Comparison of Salvia officinalis L. essential oil and antifungal agents against Candida species. J. Pharm. Technol. Drug Res. 2012, 1, 1–5. [Google Scholar] [CrossRef]
- Scazzocchio, F.; D’auria, F.D.; Alessandrini, D.; Pantanella, F. Multifactorial aspects of antimicrobial activity of propolis. Microbiol. Res. 2006, 161, 327–333. [Google Scholar] [CrossRef]
- Freires, I.A.; Queiroz, V.C.; Furletti, V.F.; Ikegaki, M.; de Alencar, S.M.; Duarte, M.C.T.; Rosalen, P.L. Chemical composition and antifungal potential of Brazilian propolis against Candida spp. J. Mycol. Medical. 2016, 26, 122–132. [Google Scholar] [CrossRef]
- Stepanović, S.; Antić, N.; Dakić, I.; Švabić-Vlahović, M. In vitro antimicrobial activity of propolis and synergism between propolis and antimicrobial drugs. Microbiol. Res. 2003, 158, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Pertino, M.W.; Theoduloz, C.; Butassi, E.; Zacchino, S.; Schmeda-Hirschmann, G. Synthesis, antiproliferative and antifungal activities of 1,2,3-Triazole-substituted carnosic acid and carnosol derivatives. Molecules 2015, 20, 8666–8686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siqueira, A.B.; Rodriguez, L.R.; Santos, R.K.; Marinho, R.R.B.; Abreu, S.; Peixoto, R.F.; Gurgel, B.C.D.V. Antifungal activity of propolis against Candida species isolated from cases of chronic periodontitis. Braz. Oral Res. 2015, 29, 1–6. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Antifungal Agents Breakpoint Tables for Interpretation of MICs European Committee on Antimicrobial Susceptibility Testing Antifungal Agents Breakpoint Tables for Interpretation of MICs. Available online: http://www.eucast.org/mic_distributions_and_ecoffs/ (accessed on 17 April 2020).
- Wuyts, J.; Van Dijck, P.; Holtappels, M. Fungal persister cells: The basis for recalcitrant infections? PLoS Path. 2018, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Suleman, T.; van Vuuren, S.; Sandasi, M.; Viljoen, A.M. Antimicrobial activity and chemometric modelling of South African propolis. J. Appl. Microbiol. 2015, 119, 981–990. [Google Scholar] [CrossRef]
- Tallarida, R.J. Quantitative Methods for Assessing Drug Synergism. Genes Cancer. 2011, 2, 1003–1008. [Google Scholar] [CrossRef] [Green Version]
- Pierce, C.G.; Saville, S.P.; Lopez-Ribot, J.L. High-content phenotypic screenings to identify inhibitors of Candida albicans biofilm formation and filamentation. Pathogens Dis. 2014, 70, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Manke, M.B.; Raut, J.S.; Dhawale, S.C.; Karuppayil, S.M. Antifungal activity of Helicteres isora Linn. Fruit extracts against planktonic and biofilm growth of Candida albicans. J. Biol. Active Prod. Nat. 2015, 5, 357–364. [Google Scholar]
- Mukherjee, P.K.; Chandra, J. Candida Biofilms: Development, architecture, and resistance. Microbiol. Spectr. 2015, 3, 1–14. [Google Scholar]
- Scudellari, M. The science myths that will not die. Nature 2015, 528, 322–325. [Google Scholar] [CrossRef] [Green Version]
- Metodiewa, D.; Jaiswal, A.K.; Cenas, N.; Dickancaité, E.; Segura-Aguilar, J. Quercetin may act as a cytotoxic prooxidant after its metabolic activation to semiquinone and quinoidal product. Free Radic. Biol. Med. 1999, 26, 107–116. [Google Scholar] [CrossRef]
- Halliwell, B. Are polyphenols antioxidants or pro-oxidants? What do we learn from cell culture and in vivo studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar] [CrossRef]
- Vella, F.; Ferry, G.; Delagrange, P.; Boutin, J.A. NRH:quinone reductase 2: An enzyme of surprises and mysteries. Biochem. Pharmacol. 2005, 71, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Collado-González, M.; Guirao-Abad, J.P.; Sánchez-Fresneda, R.; Belchí-Navarro, S.; Argüelles, J.C. Resveratrol lacks antifungal activity against Candida albicans. World J. Microbiol. Biotechnol. 2012, 28, 2441–2446. [Google Scholar]
- Guirao-Abad, J.P.; Sánchez-Fresneda, R.; Valentín, E.; Martínez-Esparza, M.; Argüelles, J.C. Analysis of validamycin as a potential antifungal compound against Candida albicans. Internatl. Microbiol. 2013, 16, 217–225. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | % |
|---|---|
| Carnosic acid | 70%–75% (mean 72%) |
| Minor diterpenes (carnosol, rosmanol, epirosmanol and 12-methyl carnosate) | 3%–7% |
| Other plant lipids | 5%–10% |
| Plant carbohydrates | 2%–5% |
| Proteins | 0%–0.5% |
| Water | 2%–4% |
| Mineral salts | 1%–2% |
| Other plant materials | 2%–4% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Argüelles, A.; Sánchez-Fresneda, R.; Guirao-Abad, J.P.; Belda, C.; Lozano, J.A.; Solano, F.; Argüelles, J.-C. Novel Bi-Factorial Strategy against Candida albicans Viability Using Carnosic Acid and Propolis: Synergistic Antifungal Action. Microorganisms 2020, 8, 749. https://doi.org/10.3390/microorganisms8050749
Argüelles A, Sánchez-Fresneda R, Guirao-Abad JP, Belda C, Lozano JA, Solano F, Argüelles J-C. Novel Bi-Factorial Strategy against Candida albicans Viability Using Carnosic Acid and Propolis: Synergistic Antifungal Action. Microorganisms. 2020; 8(5):749. https://doi.org/10.3390/microorganisms8050749
Chicago/Turabian StyleArgüelles, Alejandra, Ruth Sánchez-Fresneda, José P. Guirao-Abad, Cristóbal Belda, José Antonio Lozano, Francisco Solano, and Juan-Carlos Argüelles. 2020. "Novel Bi-Factorial Strategy against Candida albicans Viability Using Carnosic Acid and Propolis: Synergistic Antifungal Action" Microorganisms 8, no. 5: 749. https://doi.org/10.3390/microorganisms8050749