NR1 and NR3B Composed Intranuclear N-methyl-d-aspartate Receptor Complexes in Human Melanoma Cells


Abstract
:1. Introduction
2. Results
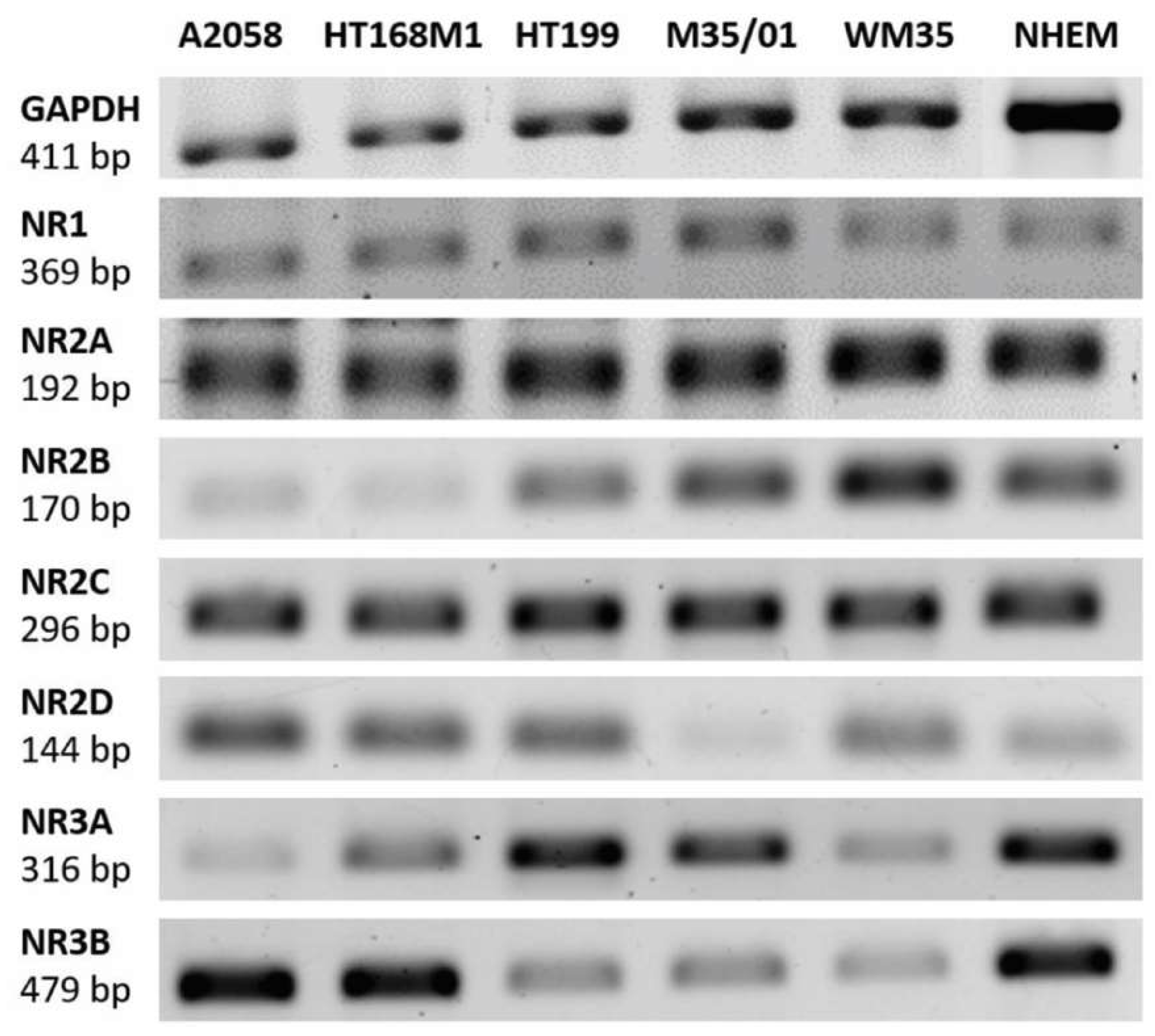
2.1. Melanocytes and Melanoma Cells Express NMDAR Subunit mRNAs
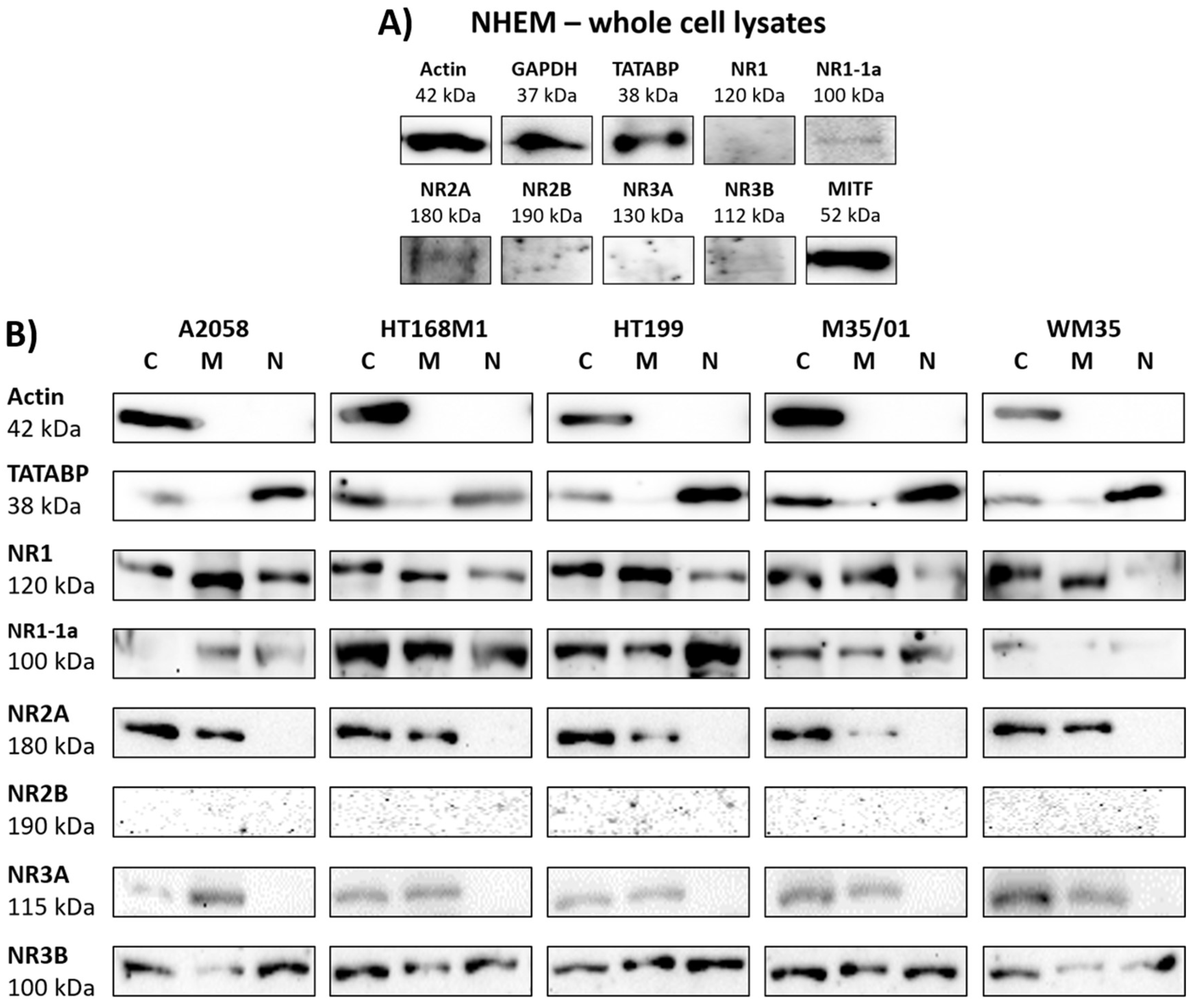
2.2. NR1 and NR3 Subunits Appear in Melanoma
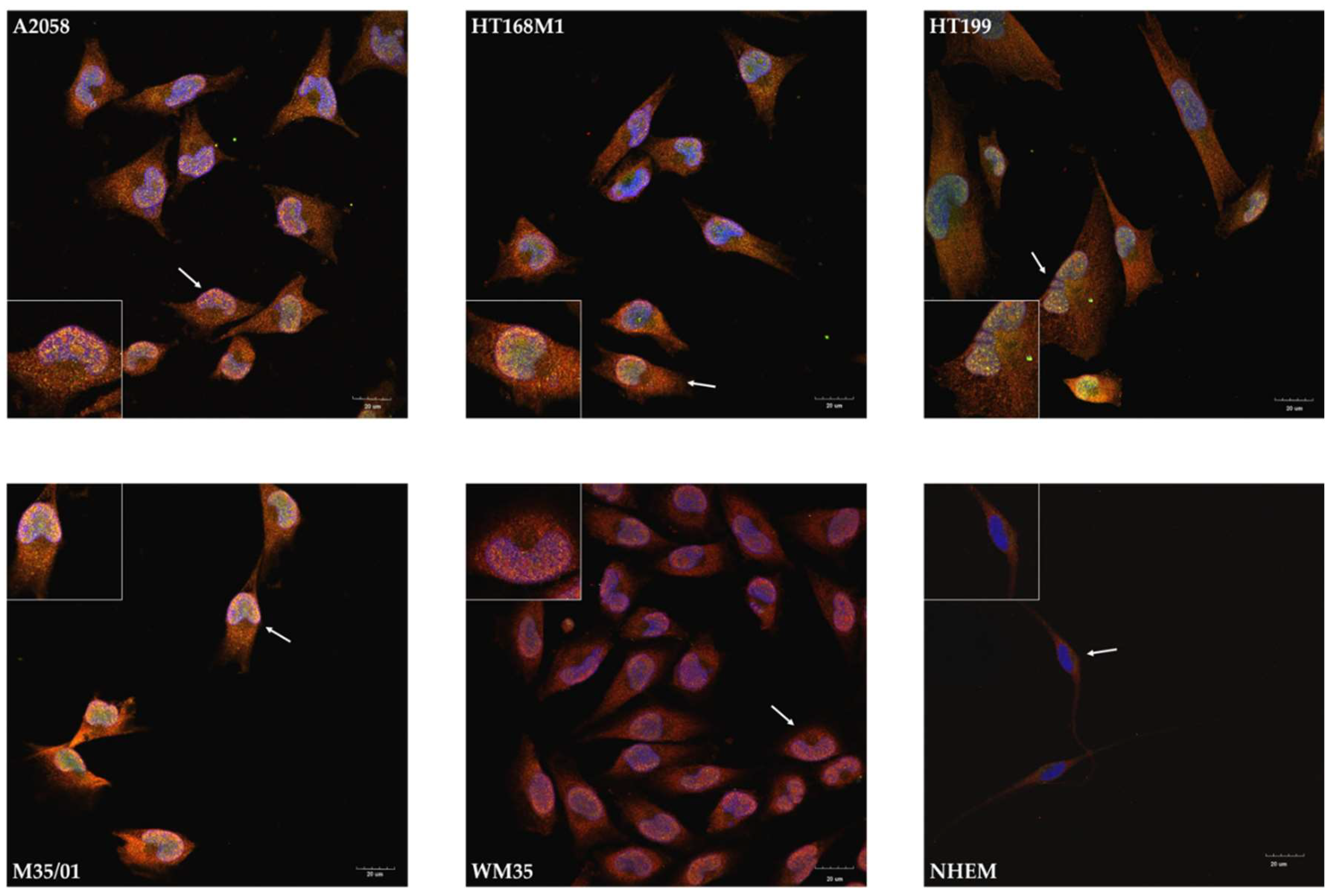
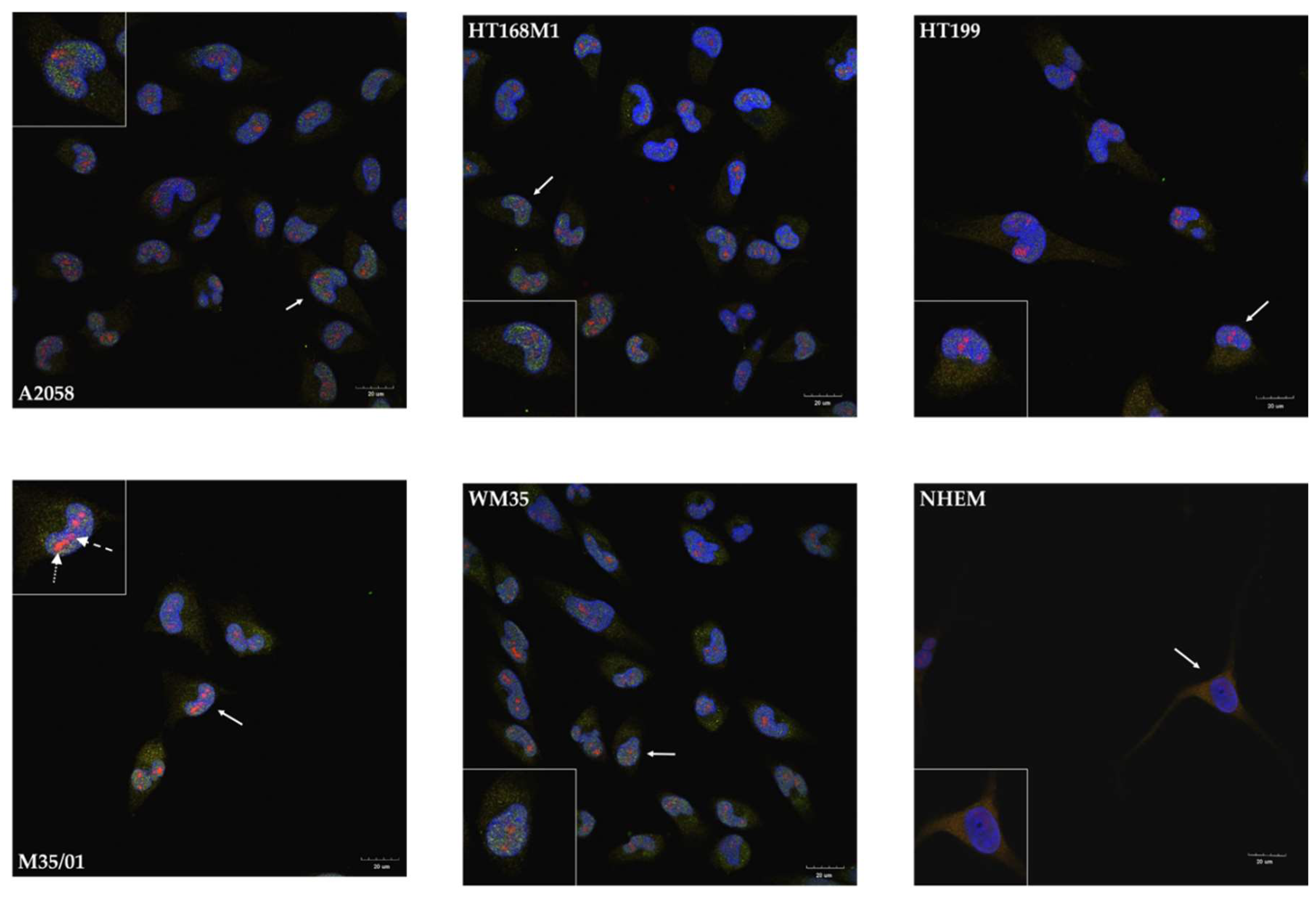
2.3. Unusual Nuclear Colocalization of Subunits in Cultured Melanoma Cells, But Not in Melanocytes
3. Discussion
4. Materials and Methods
4.1. Melanoma and Melanocyte Cell Cultures
4.2. mRNA Expression Analysis Using Reverse Transcription Followed by PCR (RT-PCR)
4.3. SDS-PAGE and Western Blot Analysis
4.4. Subcellular Fractionation of Melanoma Cells
4.5. Immunocytochemistry and Confocal Microscopy
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | central nervous system |
| iGluR | ionotropic glutamate receptor |
| mGluR | metabotropic glutamate receptor |
| NMDA | N-methyl-d-aspartate |
| NMDAR | N-methyl-d-aspartate receptor |
| NLS | Nuclear localization signal |
| NHEM | normal human epidermal melanocytes |
| RT-PCR | reverse transcription followed by polymerase chain reaction |
| dNTP | deoxynucleotide triphosphate |
| SDS-PAGE | sodium-dodecyl-sulphate polyacrylamide gel electrophoresis |
| RIPA | radio immuno precipitation assay |
| EDTA | ethylene diamine tetra-acetic acid |
| EGTA | ethylene glycol tetra-acetic acid |
| PBS | phosphate buffered saline |
| PBST | phosphate buffered saline supplemented with 1% Tween-20 |
| HRP | horse radish peroxidase |
| HEPES | hydroxyethyl-piperazine-ethanesulfonic acid |
| DTT | dithiothreitol |
| GAPDH | glycerine aldehyde phosphate dehydrogenase |
| MITF | microphthalmia associated transcription factor |
| TATABP | TATA box binding protein |
| InsP3R | inositol trisphosphate receptor |
References
- Hogan-Cann, A.D.; Anderson, C.M. Physiological Roles of Non-Neuronal NMDA Receptors. Trends Pharmacol. Sci. 2016, 37, 750–767. [Google Scholar] [CrossRef] [PubMed]
- Teh, J.L.; Chen, S. Glutamatergic signaling in cellular transformation. Pigment Cell Melanoma Res. 2012, 25, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traynelis, S.F.; Wollmuth, L.P.; McBain, C.J.; Menniti, F.S.; Vance, K.M.; Ogden, K.K.; Hansen, K.B.; Yuan, H.; Myers, S.J.; Dingledine, R. Glutamate receptor ion channels: Structure, regulation, and function. Pharmacol. Rev. 2010, 62, 405–496. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.L.; Westbrook, G.L. Permeation and block of N-methyl-d-aspartic acid receptor channels by divalent cations in mouse cultured central neurones. J. Physiol. 1987, 394, 501–527. [Google Scholar] [CrossRef] [PubMed]
- Malenka, R.C.; Nicoll, R.A. Long-term potentiation—A decade of progress? Science 1999, 285, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, P.; Bellone, C.; Zhou, Q. NMDA receptor subunit diversity: Impact on receptor properties, synaptic plasticity and disease. Nat. Rev. Neurosci. 2013, 14, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Ciabarra, A.M.; Sullivan, J.M.; Gahn, L.G.; Pecht, G.; Heinemann, S.; Sevarino, K.A. Cloning and characterization of chi-1: A developmentally regulated member of a novel class of the ionotropic glutamate receptor family. J. Neurosci. 1995, 15, 6498–6508. [Google Scholar] [CrossRef] [PubMed]
- Nishi, M.; Hinds, H.; Lu, H.P.; Kawata, M.; Hayashi, Y. Motoneuron-specific expression of NR3B, a novel NMDA-type glutamate receptor subunit that works in a dominant-negative manner. J. Neurosci. 2001, 21, RC185. [Google Scholar] [CrossRef] [PubMed]
- Chatterton, J.E.; Awobuluyi, M.; Premkumar, L.S.; Takahashi, H.; Talantova, M.; Shin, Y.; Cui, J.; Tu, S.; Sevarino, K.A.; Nakanishi, N.; et al. Excitatory glycine receptors containing the NR3 family of NMDA receptor subunits. Nature 2002, 415, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Smothers, C.T.; Woodward, J.J. Pharmacological characterization of glycine-activated currents in HEK 293 cells expressing N-methyl-d-aspartate NR1 and NR3 subunits. J. Pharmacol. Exp. Ther. 2007, 322, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Hinoi, E.; Takarada, T.; Ueshima, T.; Tsuchihashi, Y.; Yoneda, Y. Glutamate signaling in peripheral tissues. Eur. J. Biochem. 2004, 271, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Stepulak, A.; Rola, R.; Polberg, K.; Ikonomidou, C. Glutamate and its receptors in cancer. J. Neural Transm. 2014, 121, 933–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummins, D.L.; Cummins, J.M.; Pantle, H.; Silverman, M.A.; Leonard, A.L.; Chanmugam, A. Cutaneous malignant melanoma. Mayo Clin. Proc. 2006, 81, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Erdei, E.; Torres, S.M. A new understanding in the epidemiology of melanoma. Expert Rev. Anticancer Ther. 2010, 10, 1811–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namkoong, J.; Shin, S.S.; Lee, H.J.; Marin, Y.E.; Wall, B.A.; Goydos, J.S.; Chen, S. Metabotropic glutamate receptor 1 and glutamate signaling in human melanoma. Cancer Res. 2007, 67, 2298–2305. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; He, C.D.; Liu, J.; Sun, C.; Lu, P.; Li, L.; Gao, L.; Zhang, Y.; Xu, Y.; Shan, L.; et al. Blocking glutamate-mediated signalling inhibits human melanoma growth and migration. Exp. Dermatol. 2012, 21, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Walia, V.; Lin, J.C.; Teer, J.K.; Prickett, T.D.; Gartner, J.; Davis, S.; Program, N.C.S.; Stemke-Hale, K.; Davies, M.A.; et al. Exome sequencing identifies GRIN2A as frequently mutated in melanoma. Nat. Genet. 2011, 43, 442–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Mello, S.A.; Flanagan, J.U.; Green, T.N.; Leung, E.Y.; Askarian-Amiri, M.E.; Joseph, W.R.; McCrystal, M.R.; Isaacs, R.J.; Shaw, J.H.; Furneaux, C.E.; et al. Evidence That GRIN2A Mutations in Melanoma Correlate with Decreased Survival. Front. Oncol. 2014, 3, 333. [Google Scholar] [CrossRef] [PubMed]
- Prickett, T.D.; Zerlanko, B.J.; Hill, V.K.; Gartner, J.J.; Qutob, N.; Jiang, J.; Simaan, M.; Wunderlich, J.; Gutkind, J.S.; Rosenberg, S.A.; et al. Somatic mutation of GRIN2A in malignant melanoma results in loss of tumor suppressor activity via aberrant NMDAR complex formation. J. Investig. Dermatol. 2014, 134, 2390–2398. [Google Scholar] [CrossRef] [PubMed]
- Hoogduijn, M.J.; Hitchcock, I.S.; Smit, N.P.; Gillbro, J.M.; Schallreuter, K.U.; Genever, P.G. Glutamate receptors on human melanocytes regulate the expression of MiTF. Pigment Cell Res. 2006, 19, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Wang, N.; Gao, L.; Li, L.; Zheng, S.; Liu, Y.; Ozukum, M.; Nikiforova, A.; Zhao, G.; Song, Z. The effect of the NMDA receptor-dependent signaling pathway on cell morphology and melanosome transfer in melanocytes. J. Dermatol. Sci. 2016, 84, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.D.; Mattar, P.; Marsh, D.R.; Jordan, V.; Weaver, L.C.; Dekaban, G.A. The C-terminal C1 cassette of the N-methyl-d-aspartate receptor 1 subunit contains a bi-partite nuclear localization sequence. J. Neurochem. 2002, 81, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Resende, R.R.; Andrade, L.M.; Oliveira, A.G.; Guimaraes, E.S.; Guatimosim, S.; Leite, M.F. Nucleoplasmic calcium signaling and cell proliferation: Calcium signaling in the nucleus. Cell Commun. Signal. CCS 2013, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Bootman, M.D.; Fearnley, C.; Smyrnias, I.; MacDonald, F.; Roderick, H.L. An update on nuclear calcium signalling. J. Cell Sci. 2009, 122, 2337–2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, M.T.; Garcia-Sancho, J. Nuclear Ca(2+) signalling. Cell Calcium 2011, 49, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Adebanjo, O.A.; Biswas, G.; Moonga, B.S.; Anandatheerthavarada, H.K.; Sun, L.; Bevis, P.J.; Sodam, B.R.; Lai, F.A.; Avadhani, N.G.; Zaidi, M. Novel biochemical and functional insights into nuclear Ca(2+) transport through IP(3)Rs and RyRs in osteoblasts. Am. J. Physiol. Renal Physiol. 2000, 278, F784–F791. [Google Scholar] [CrossRef] [PubMed]
- Gerasimenko, O.V.; Gerasimenko, J.V.; Tepikin, A.V.; Petersen, O.H. ATP-dependent accumulation and inositol trisphosphate- or cyclic ADP-ribose-mediated release of Ca2+ from the nuclear envelope. Cell 1995, 80, 439–444. [Google Scholar] [CrossRef]
- Shirakawa, H.; Miyazaki, S. Spatiotemporal analysis of calcium dynamics in the nucleus of hamster oocytes. J. Physiol. 1996, 494 Pt 1, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Yang, D.; Lan, X.; Li, K.; Li, X.; Chen, J.; Zhang, Y.; Xiao, R.P.; Han, Q.; Cheng, H. Nuclear Ca2+ sparks and waves mediated by inositol 1,4,5-trisphosphate receptors in neonatal rat cardiomyocytes. Cell Calcium 2008, 43, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Campden, R.; Audet, N.; Hebert, T.E. Nuclear G protein signaling: New tricks for old dogs. J. Cardiovasc. Pharmacol. 2015, 65, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Matzke, A.J.; Weiger, T.M.; Matzke, M. Ion channels at the nucleus: Electrophysiology meets the genome. Mol. Plant 2010, 3, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Avedanian, L.; Jacques, D. Nuclear membrane receptors and channels as targets for drug development in cardiovascular diseases. Can. J. Physiol. Pharmacol. 2009, 87, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Jong, Y.J.; Kumar, V.; Kingston, A.E.; Romano, C.; O’Malley, K.L. Functional metabotropic glutamate receptors on nuclei from brain and primary cultured striatal neurons. Role of transporters in delivering ligand. J. Biol. Chem. 2005, 280, 30469–30480. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Jong, Y.J.; O’Malley, K.L. Activated nuclear metabotropic glutamate receptor mGlu5 couples to nuclear Gq/11 proteins to generate inositol 1,4,5-trisphosphate-mediated nuclear Ca2+ release. J. Biol. Chem. 2008, 283, 14072–14083. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.F.; Thrower, E.C.; Echevarria, W.; Koulen, P.; Hirata, K.; Bennett, A.M.; Ehrlich, B.E.; Nathanson, M.H. Nuclear and cytosolic calcium are regulated independently. Proc. Natl. Acad. Sci. USA 2003, 100, 2975–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, C.M.; Wee, K.S. New insights into the not-so-new NR3 subunits of N-methyl-D-aspartate receptor: Localization, structure, and function. Mol. Pharmacol. 2010, 78, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cokol, M.; Nair, R.; Rost, B. Finding nuclear localization signals. EMBO Rep. 2000, 1, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bading, H. Nuclear calcium signalling in the regulation of brain function. Nat. Rev. Neurosci. 2013, 14, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.P.; Custodio, J.B.; Santos, A.E. Ionotropic glutamate receptor antagonists and cancer therapy: Time to think out of the box? Cancer Chemother. Pharmacol. 2017, 79, 219–225. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Nucleotide Sequence (5′→3′) | GenBank ID | Annealing Temperature | Amplimer Size (bp) |
|---|---|---|---|---|---|
| NR1 (GRIN1) | sense | CCA CGC TGA GTG ATG GGA CA (1568–1587) | NM_007327.3 | 56.5 °C | 369 |
| antisense | GGT ACT TGA AGG GCT TGG AAA A (1936–1915) | ||||
| NR2A (GRIN2A) | sense | TGC CAC AAC GAG AAG AAC (2946–2963) | NM_001134407.2 | 57 °C | 192 |
| antisense | GAT GGA GAA GAG CAA CCC (3137–3120) | ||||
| NR2B (GRIN2B) | sense | CAA GAG GCG TAA GCA GC (3413–3429) | NM_000834.3 | 55 °C | 170 |
| antisense | CCA GGT AGA AGT CCC GTA G (3582–3564) | ||||
| NR2C (GRIN2C) | sense | ACG TCC ACG GCA TTG TCT (676–693) | NM_000835.4 | 61 °C | 296 |
| antisense | GGA TGT CAT TCC CAT AAC CA (952–971) | ||||
| NR2D (GRIN2D) | sense | TGG CAA GCA CGG AAA GAA GAT C (1618–1639) | NM_000836.2 | 62.5 °C | 144 |
| antisense | TCC ACG AAG GGG ACG GAG AAG T (1761–1740) | ||||
| NR3A (GRIN3A) | sense | ACA CAA AAC CCA CTT CCA ACA TCC (2107–2130) | NM_133445.2 | 58 °C | 316 |
| antisense | TGC TCC ATA CTT TCC ATC CCC TAC (2422–2399) | ||||
| NR3B (GRIN3B) | sense | CGC AAG TGC TGC TAC GGC TAC (1436–1456) | NM_138690.2 | 61 °C | 479 |
| antisense | ACG GTG CGT CTG AAG AGG ATG (1914–1894) | ||||
| GAPDH | sense | CCA GAA GAC TGT GGA TGG CC (740–759) | NM_002046.5 | 54 °C | 411 |
| antisense | CTG TAG CCA AAT TCG TTG TC (1150–1131) |
| Antibody | Host Animal | Dilution for Western Blot | Dilution for Immunocytochemistry | Distributor, Cat. Num. |
|---|---|---|---|---|
| Anti-NR1 | rabbit, monoclonal | 1:250 | 1:50 | Cell Signaling Technology, Danvers, MO, USA |
| #5704S lot:2 | ||||
| Anti-NR1-1a | rabbit, polyclonal | 1:200 | 1:50 | Merck-Millipore, Billerica, MA, USA |
| AB5046P | ||||
| Anti-NR2A | rabbit, polyclonal | 1:200 | - | Cell Signaling Technology, Danvers, MO, USA |
| #4205S lot:1 | ||||
| Anti-NR2B | rabbit, polyclonal | 1:200 | - | Cell Signaling Technology, Danvers, MO, USA |
| #4207S lot:2 | ||||
| Anti-NR3A | rabbit, polyclonal | 1:300 | - | Merck-Millipore, Billerica, MA, USA 07-356 |
| Anti-NR3A | rabbit, polyclonal | 1:500 | - | Alomone Labs, Jerusalem, Israel AGC-030 |
| Anti-NR3B | rabbit, polyclonal | 1:600 | - | Abcam, Cambridge, UK |
| ab35677-100 | ||||
| Anti-NR3B | rabbit, polyclonal | 1:250 | 1:50 | Alomone Labs, Jerusalem, Israel AGC-031 |
| Anti-TATABP | mouse, monoclonal | 1:500 | - | Abcam, Cambridge, UK ab51841 |
| Anti-GAPDH | rabbit, polyclonal | 1:500 | - | Abcam, Cambridge, UK Ab9485 |
| Anti-β-Actin | mouse, monoclonal | 1:10.000 | - | Sigma-Aldrich, St. Louis, MO, USA A5441 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajdú, T.; Juhász, T.; Szűcs-Somogyi, C.; Rácz, K.; Zákány, R. NR1 and NR3B Composed Intranuclear N-methyl-d-aspartate Receptor Complexes in Human Melanoma Cells. Int. J. Mol. Sci. 2018, 19, 1929. https://doi.org/10.3390/ijms19071929
Hajdú T, Juhász T, Szűcs-Somogyi C, Rácz K, Zákány R. NR1 and NR3B Composed Intranuclear N-methyl-d-aspartate Receptor Complexes in Human Melanoma Cells. International Journal of Molecular Sciences. 2018; 19(7):1929. https://doi.org/10.3390/ijms19071929
Chicago/Turabian StyleHajdú, Tibor, Tamás Juhász, Csilla Szűcs-Somogyi, Kálmán Rácz, and Róza Zákány. 2018. "NR1 and NR3B Composed Intranuclear N-methyl-d-aspartate Receptor Complexes in Human Melanoma Cells" International Journal of Molecular Sciences 19, no. 7: 1929. https://doi.org/10.3390/ijms19071929