NMR Structure of the FIV gp36 C-terminal Heptad Repeat and Membrane-Proximal External Region

 , , , ,
, , , ,  ,
,
Abstract
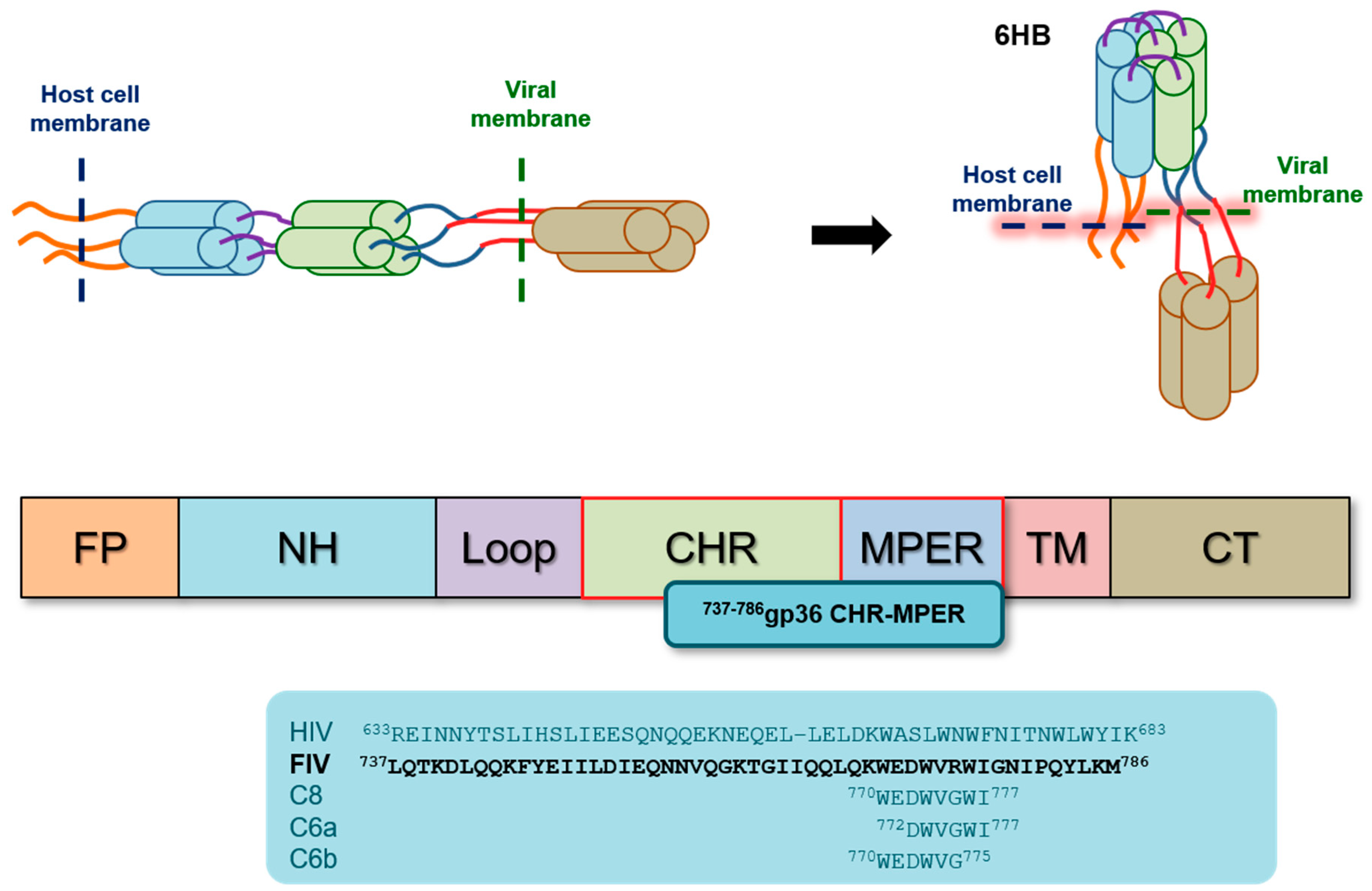
:1. Introduction
2. Results
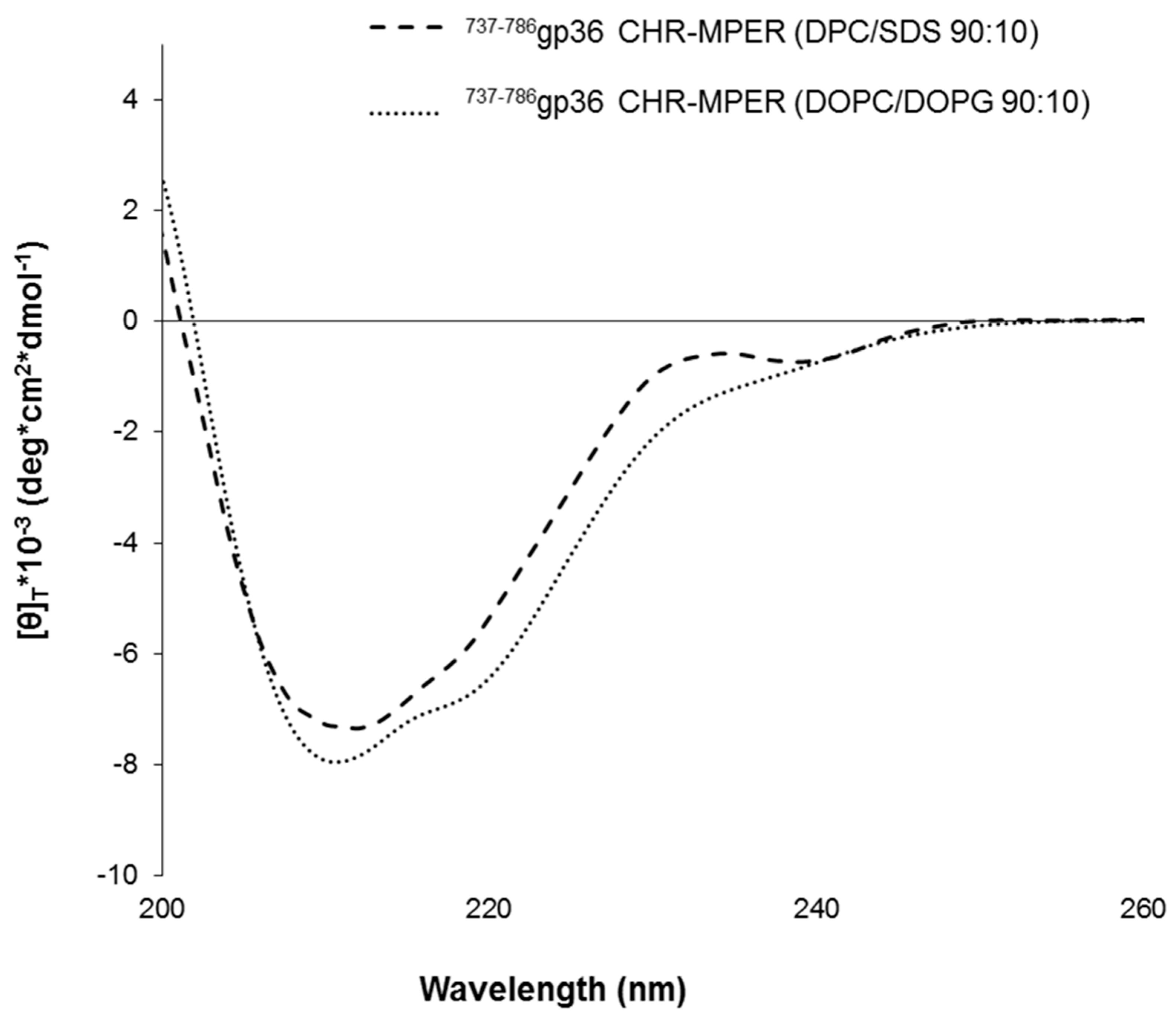
2.1. Circular Dichroism of 737-786gp36 CHR–MPER
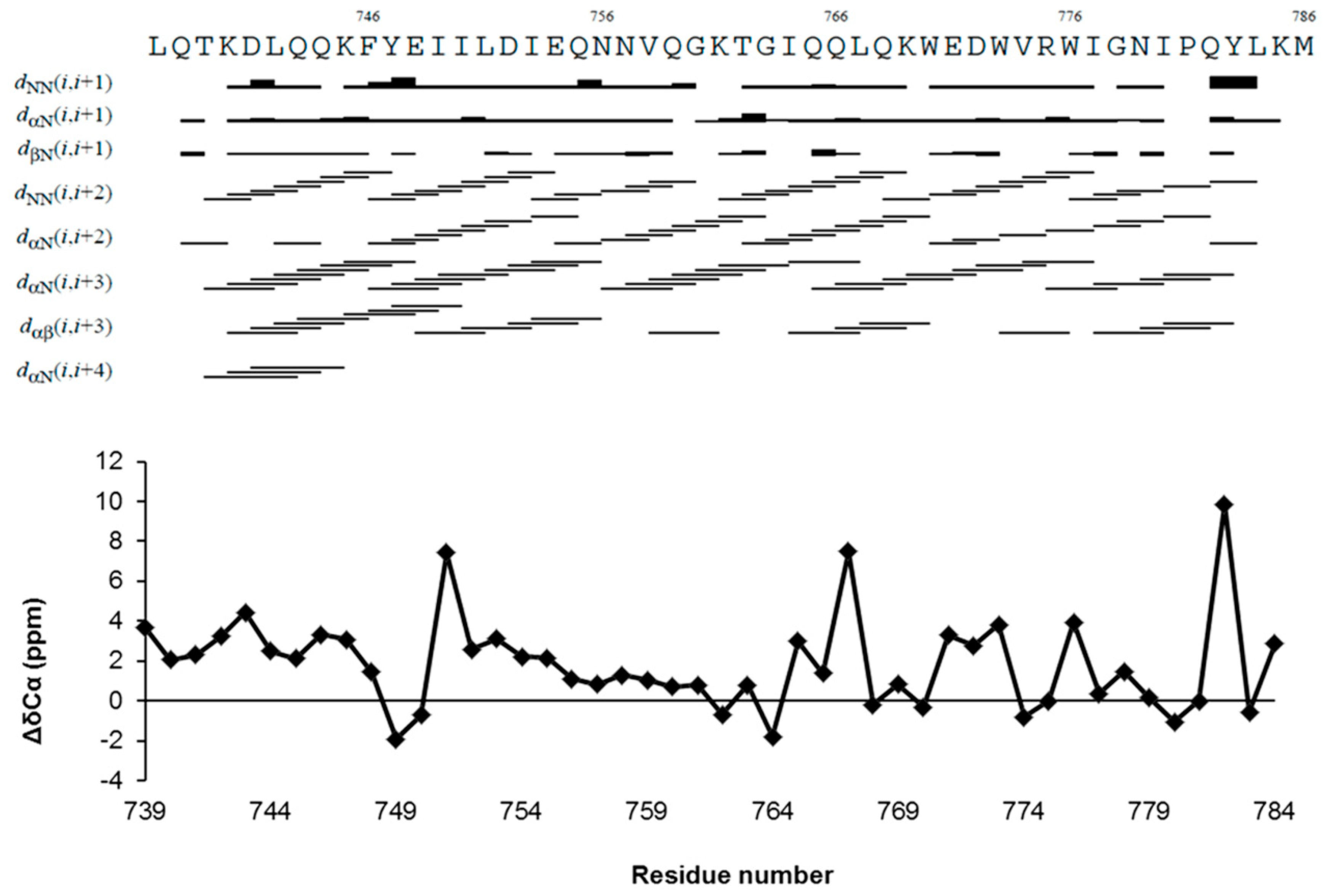
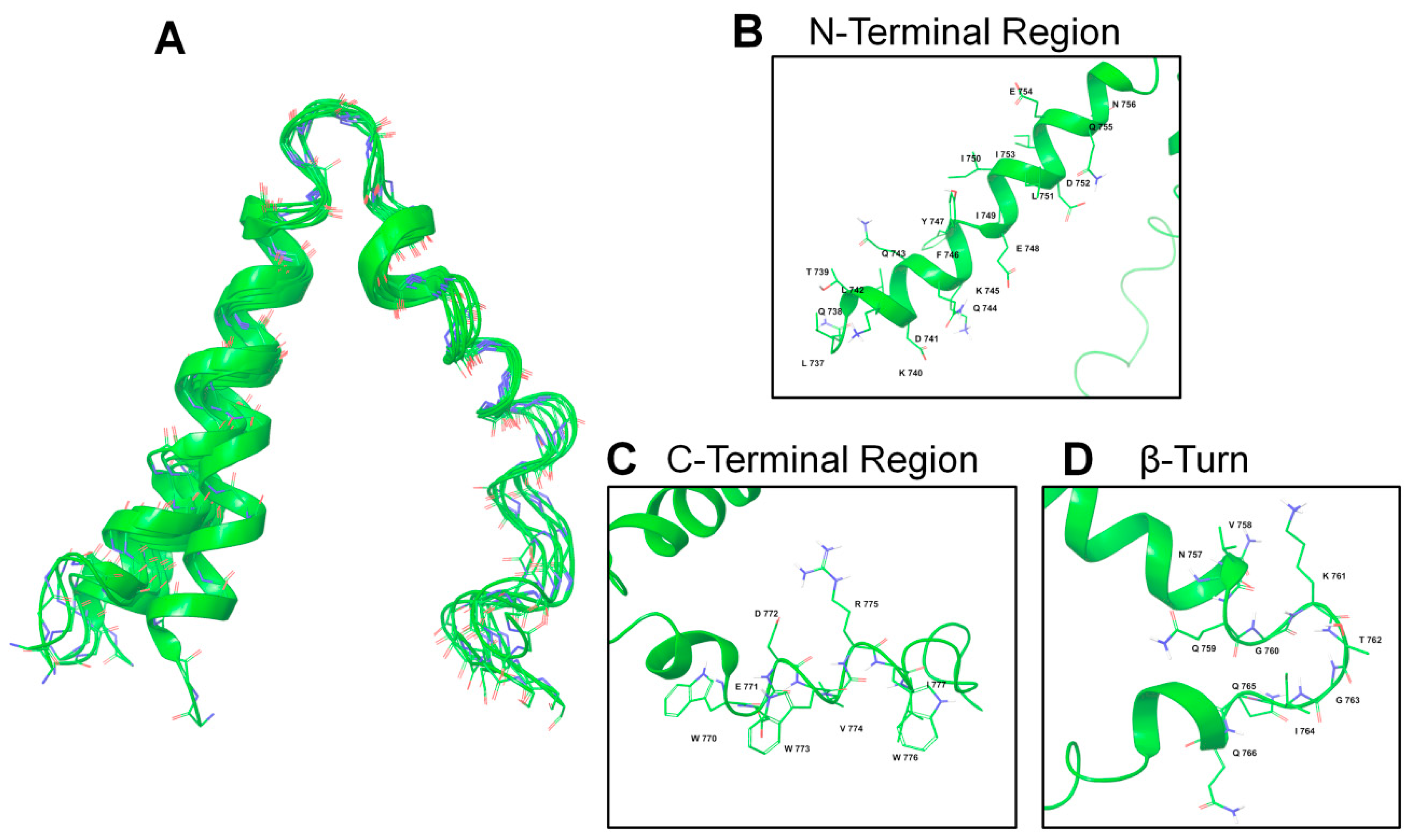
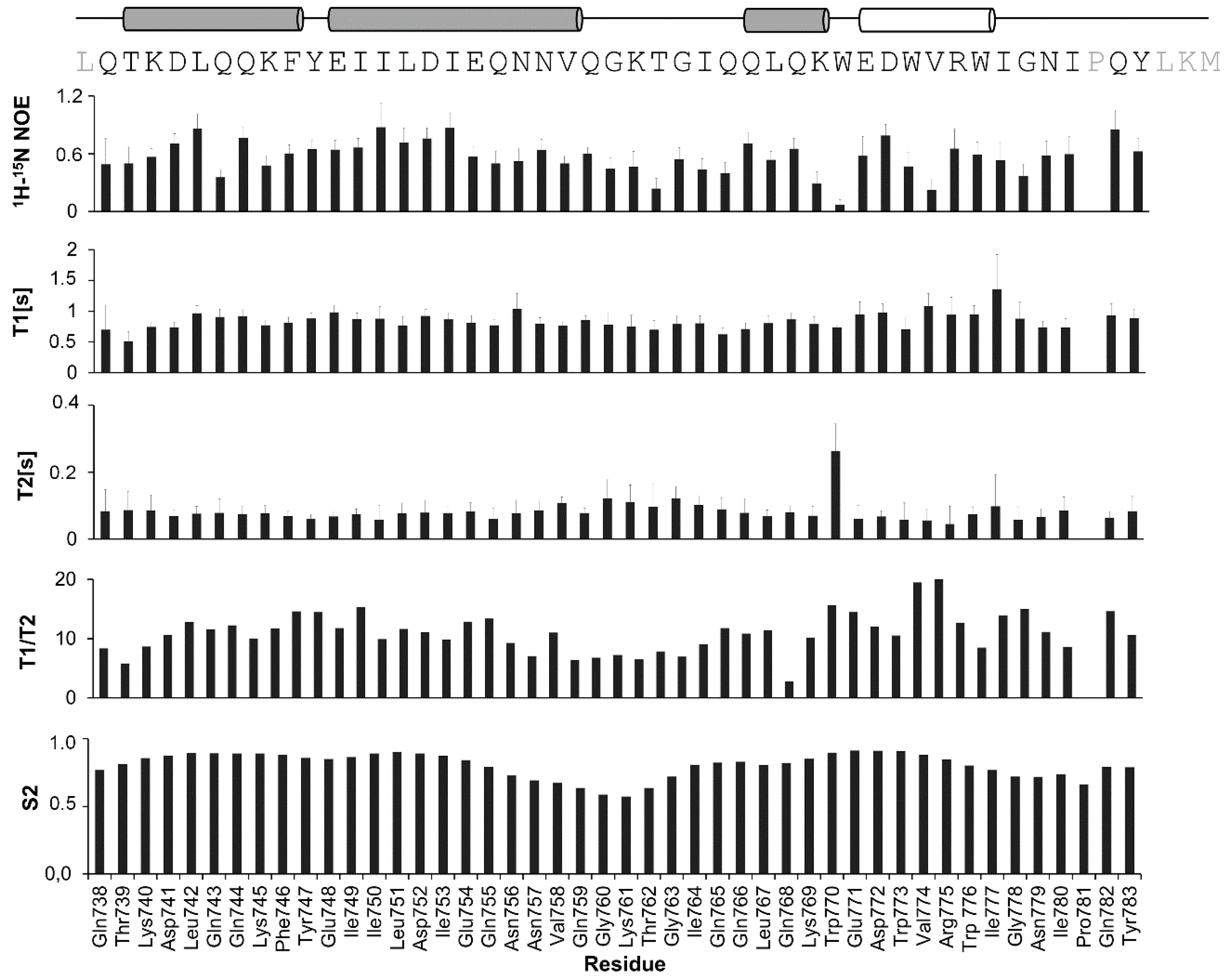
2.2. NMR Structure Determination
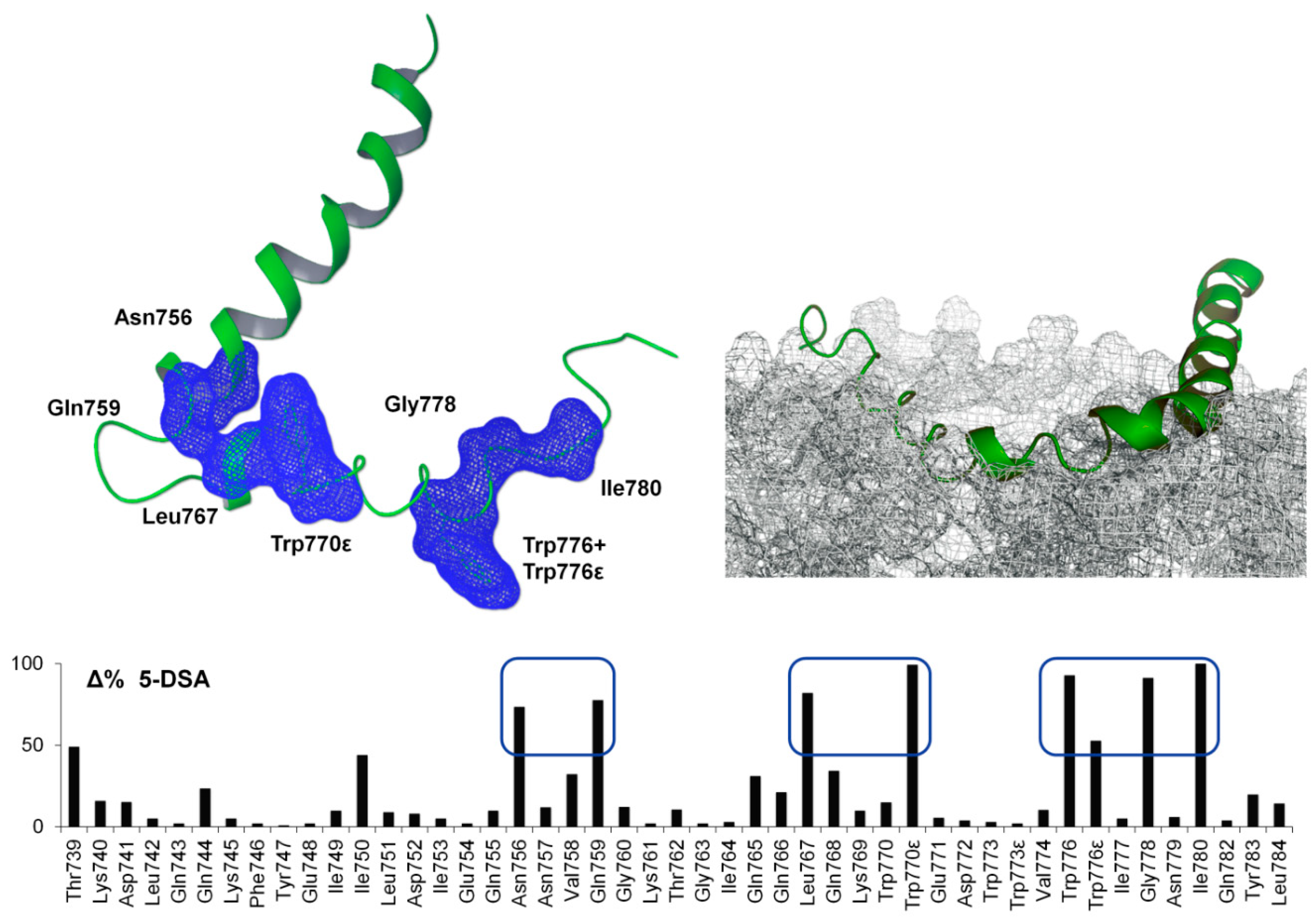
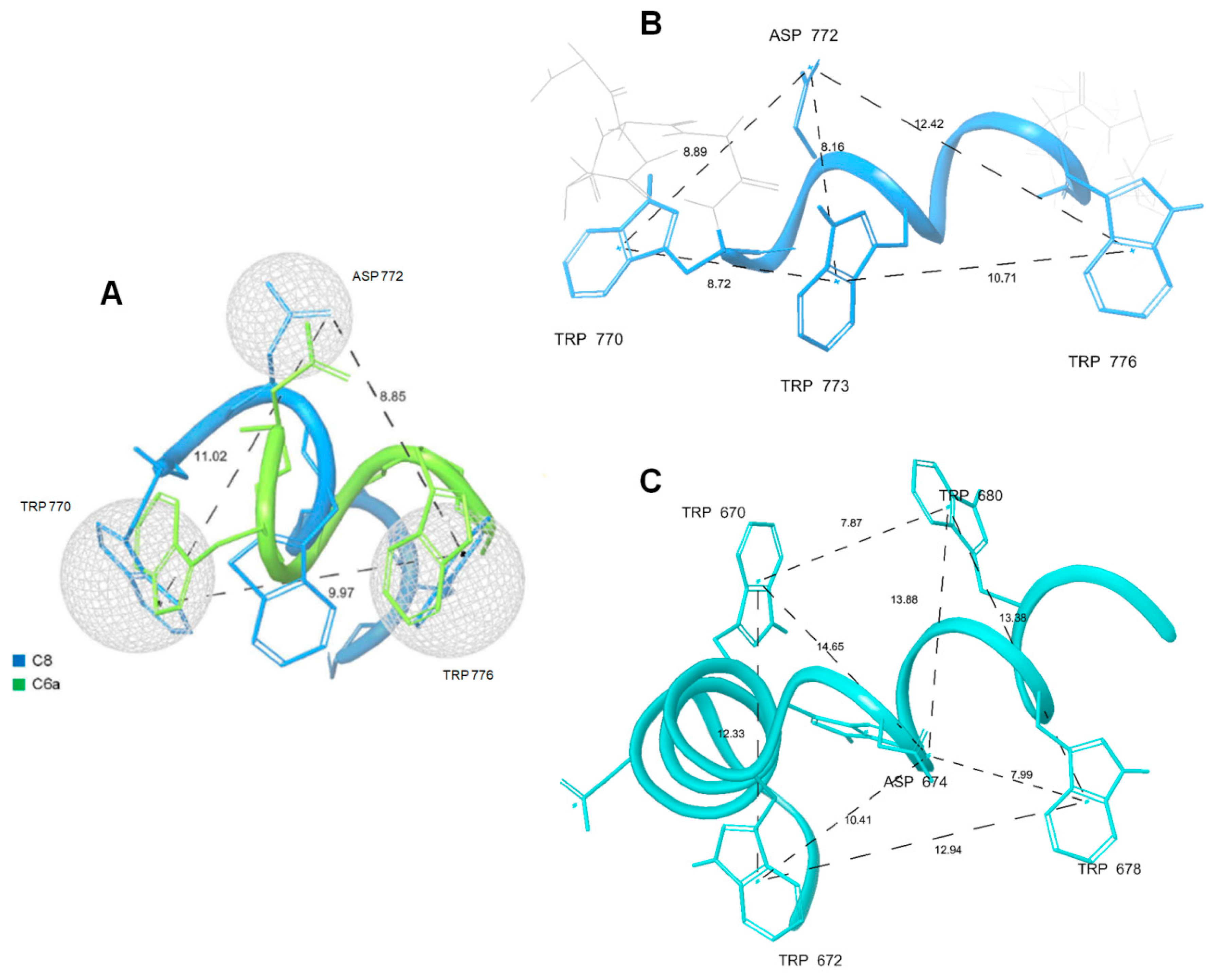
2.3. NMR Spin-Label Analysis
2.4. ESR Spin-Label Analysis
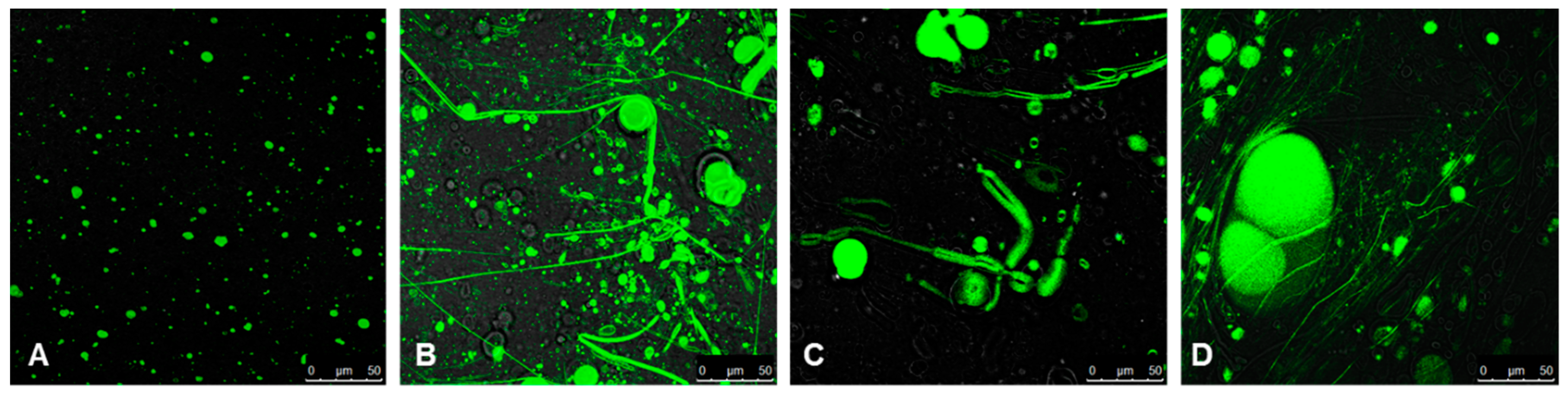
2.5. Confocal Microscopy Imaging
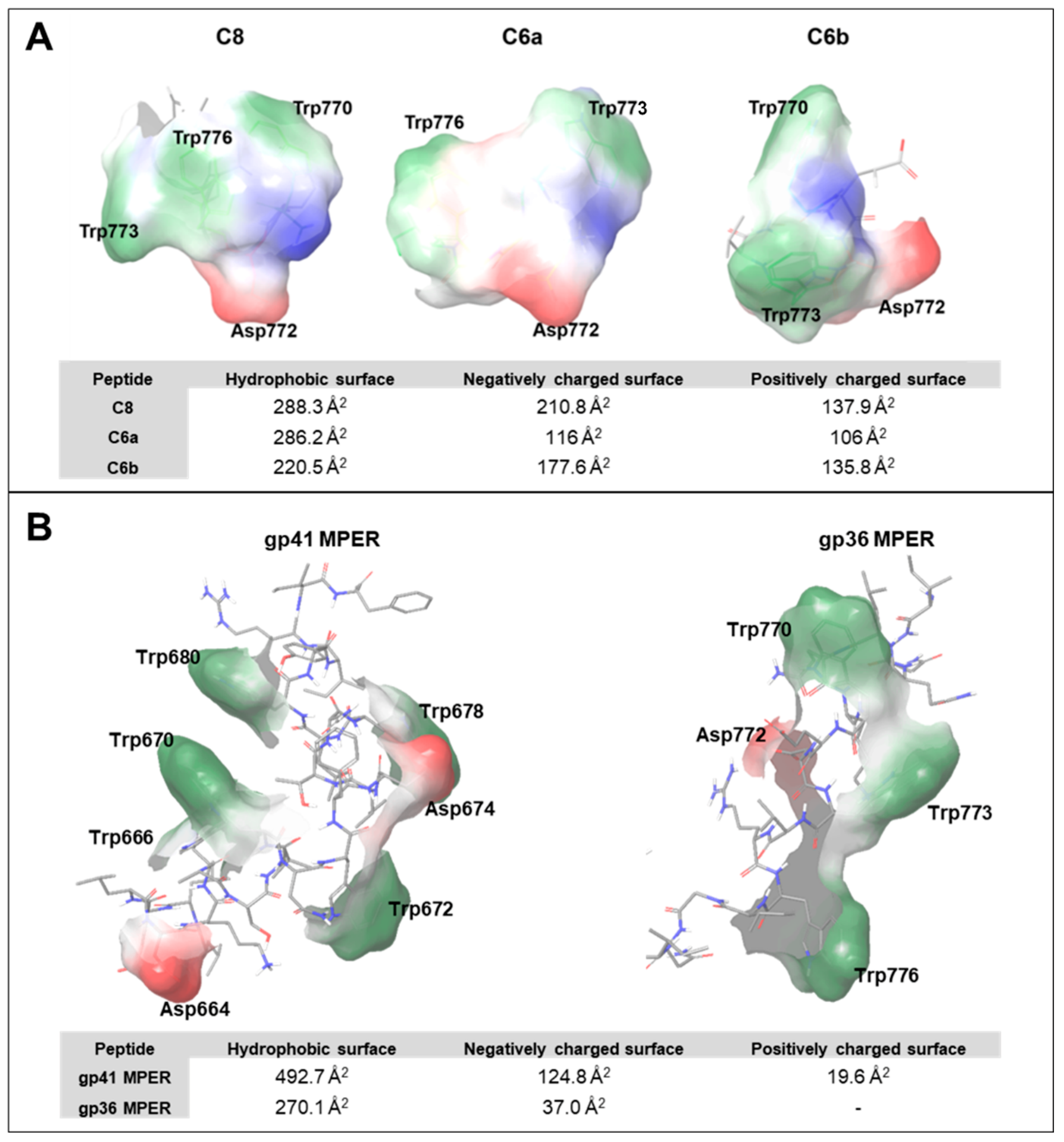
3. Discussion
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. Mass Spectrometry Analysis
4.3. Circular Dichroism Spectroscopy
4.4. NMR Spectroscopy
4.4.1. Spectra Acquisition
4.4.2. Assignment of NMR Resonances and TALOS+ Analysis
4.4.3. Structure Calculation
4.4.4. NMR Spin-Label Experiments
4.5. ESR Spectroscopy
4.6. Confocal Microscope Imaging
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pedersen, N.C.; Ho, E.W.; Brown, M.L.; Yamamoto, J.K. Isolation of a t-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome. Science 1987, 235, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Elder, J.H.; Phillips, T.R. Feline Immunodeficiency Virus As A Model For Development Of Molecular Approaches To Intervention Strategies Against Lentivirus Infections. Adv. Virus Res. 1995, 45, 225–247. [Google Scholar] [CrossRef]
- Lee, T.; Laco, G.S.; Torbett, B.E.; Fox, H.S.; Lerner, D.L.; Elder, J.H.; Wong, C.H. Analysis of the S3 and S3 ‘ subsite specificities of feline immunodeficiency virus (FIV) protease: Development of a broad-based protease inhibitor efficacious against FIV, SIV and HIV in vitro and ex vivo. Proc. Natl. Acad. Sci. USA 1998, 95, 939–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willett, B.J.; Flynn, J.N.; Hosie, M.J. FIV infection of the domestic cat: An animal model for AIDS. Immunol. Today 1997, 18, 182–189. [Google Scholar] [CrossRef]
- Molinos-Albert, L.M.; Clotet, B.; Blanco, J.; Carrillo, J. Immunologic Insights on the Membrane Proximal External Region: A Major Human Immunodeficiency Virus Type-1 Vaccine Target. Front. Immunol. 2017, 8, 1154. [Google Scholar] [CrossRef] [Green Version]
- Frey, S.C.S.; Hoover, E.A.; Mullins, J.I. Feline immunodeficiency virus cell entry. J. Virol. 2001, 75, 5433–5440. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, R.; Sodroski, J. The HIV-1 envelope glycoproteins: Fusogens, antigens, and immunogens. Science 1998, 280, 1884–1888. [Google Scholar] [CrossRef]
- Harrison, S.C. Viral membrane fusion. Nat. Struct. Mol. Biol. 2008, 15, 690–698. [Google Scholar] [CrossRef]
- Sackett, K.; Shai, Y. The HIV-1 gp41 N-terminal heptad repeat plays an essential role in membrane fusion. Biochemistry 2002, 41, 4678–4685. [Google Scholar] [CrossRef]
- Dwyer, J.J.; Wilson, K.L.; Davison, D.K.; Freel, S.A.; Seedorff, J.E.; Wring, S.A.; Tvermoes, N.A.; Matthews, T.J.; Greenberg, M.L.; Delmedico, M.K. Design of helical, oligomeric HIV-1 fusion inhibitor peptides with potent activity against enfuvirtide-resistant virus. Proc. Natl. Acad. Sci. USA 2007, 104, 12772–12777. [Google Scholar] [CrossRef] [Green Version]
- Steffen, I.; Poehlmann, S. Peptide-Based Inhibitors of the HIV Envelope Protein and Other Class I Viral Fusion Proteins. Curr. Pharm. Des. 2010, 16, 1143–1158. [Google Scholar] [CrossRef] [PubMed]
- Rowse, M. The Mechanism of Small Molecule Viral Fusion Inhibitors; The University of Alabama at Birmingham: Birmingham, AL, USA, 2015. [Google Scholar]
- Liu, H.; Su, X.; Si, L.; Lu, L.; Jiang, S. The development of HIV vaccines targeting gp41 membrane-proximal external region (MPER): Challenges and prospects. Protein Cell 2018, 9, 596–615. [Google Scholar] [CrossRef] [PubMed]
- Barbato, G.; Bianchi, E.; Ingallinella, P.; Hurni, W.H.; Miller, M.D.; Cilibertol, G.; Cortese, R.; Bazzo, R.; Shiver, J.W.; Pessi, A. Structural analysis of the epitope of the Anti-HIV antibody 2F5 sheds light into its mechanism of neutralization and HIV fusion. J. Mol. Biol. 2003, 330, 1101–1115. [Google Scholar] [CrossRef]
- Giannecchini, S.; Di Fenza, A.; D’Ursi, A.M.; Matteucci, D.; Rovero, P.; Bendinelli, M. Antiviral activity and conformational features of an octapeptide derived from the membrane-proximal ectodomain of the feline immunodeficiency virus transmembrane glycoprotein. J. Virol. 2003, 77, 3724–3733. [Google Scholar] [CrossRef] [Green Version]
- Mizukoshi, F.; Baba, K.; Goto, Y.; Setoguchi, A.; Fujino, Y.; Ohno, K.; Oishi, S.; Kodera, Y.; Fujii, N.; Tsujimoto, H. Antiviral activity of membrane fusion inhibitors that target gp40 of the feline immunodeficiency virus envelope protein. Vet. Microbiol. 2009, 136, 155–159. [Google Scholar] [CrossRef]
- Pancino, G.; Camoin, L.; Sonigo, P. Structural-analysis of the principal immunodominant domain of the feline immunodeficiency virus transmembrane glycoprotein. J. Virol. 1995, 69, 2110–2118. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, S.; Massi, C.; Indino, E.; LaRosa, C.; Mazzetti, P.; Falcone, M.L.; Rovero, P.; Fissi, A.; Pieroni, O.; Bandecchi, P.; et al. Inhibition of feline immunodeficiency virus infection in vitro by envelope glycoprotein synthetic peptides. Virology 1996, 220, 274–284. [Google Scholar] [CrossRef] [Green Version]
- Giannecchini, S.; Bonci, F.; Pistello, M.; Matteucci, D.; Sichi, O.; Rovero, P.; Bendinelli, M. The membrane-proximal tryptophan-rich region in the transmembrane glycoprotein ectodomain of feline immunodeficiency virus is important for cell entry. Virology 2004, 320, 156–166. [Google Scholar] [CrossRef]
- Scrima, M.; Di Marino, S.; Grimaldi, M.; Campana, F.; Vitiello, G.; Piotto, S.P.; D’Errico, G.; D’Ursi, A.M. Structural features of the C8 antiviral peptide in a membrane-mimicking environment. BBA Biomembr. 2014, 1838, 1010–1018. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Deng, Y.; Dey, A.K.; Moore, J.P.; Lu, M. Structure of the HIV-1 gp41 membrane-proximal ectodomain region in a putative prefusion conformation. Biochemistry 2009, 48, 2915–2923. [Google Scholar] [CrossRef] [Green Version]
- Buzon, V.; Natrajan, G.; Schibli, D.; Campelo, F.; Kozlov, M.M.; Weissenhorn, W. Crystal structure of HIV-1 gp41 including both fusion peptide and membrane proximal external regions. PLoS Pathog. 2010, 6, e1000880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, W.; Bohon, J.; Han, D.; Habte, H.; Qin, Y.; Cho, M.W.; Chance, M.R. Structural characterization of HIV GP41 with the membrane proximal external region. J. Biol. Chem. 2010, 285, 24290–24298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Shaik, M.M.; Cai, Y.; Ghantous, F.; Piai, A.; Peng, H.; Rits-Volloch, S.; Liu, Z.; Harrison, S.C.; Seaman, M.S.; et al. Structure of the membrane proximal external region of HIV-1 envelope glycoprotein. Proc. Natl. Acad. Sci. USA 2018, 115, E8892–E8899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Errico, G.; D’Ursi, A.M.; Marsh, D. Interaction of a peptide derived from glycoprotein gp36 of feline immunodeficiency virus and its lipoylated analogue with phospholipid membranes. Biochemistry 2008, 47, 5317–5327. [Google Scholar] [CrossRef]
- Desmaris, F.; Lemaire, D.; Ricard-Blum, S.; Chatrenet, B.; Forest, E. Structural characterization of the feline-immunodeficiency-virus envelope glycoprotein 36 ectodomain for the development of new antivirals. Biochem. J. 2005, 389, 559–567. [Google Scholar] [CrossRef] [Green Version]
- D’Ursi, A.M.; Giannecchini, S.; Esposito, C.; Alcaro, M.C.; Sichi, O.; Armenante, M.R.; Carotenuto, A.; Papini, A.M.; Bendinelli, M.; Rovero, P. Development of antiviral fusion inhibitors: Short modified peptides derived from the transmembrane glycoprotein of feline immunodeficiency virus. ChemBioChem 2006, 7, 774–779. [Google Scholar] [CrossRef]
- D’Ursi, A.M.; Giannecchini, S.; Di Fenza, A.; Esposito, C.; Armenante, M.R.; Carotenuto, A.; Bendinelli, M.; Rovero, P. Retroinverso analogue of the antiviral octapeptide C8 inhibits feline immunodeficiency virus in serum. J. Med. Chem. 2003, 46, 1807–1810. [Google Scholar] [CrossRef]
- Giannecchini, S.; D’Ursi, A.M.; Esposito, C.; Scrima, M.; Zabogli, E.; Freer, G.; Rovero, P.; Bendinelli, M. Antibodies generated in cats by a lipopeptide reproducing the membrane-proximal external region of the feline immunodeficiency virus transmembrane enhance virus infectivity. Clin. Vaccine Immunol. 2007, 14, 944–951. [Google Scholar] [CrossRef] [Green Version]
- Esposito, C.; D’Errico, G.; Armenante, M.R.; Giannecchini, S.; Bendinelli, M.; Rovero, P.; Anna, M. Physicochemical characterization of a peptide deriving from the glycoprotein gp36 of the feline immunodeficiency virus and its lipoylated analogue in micellar systems. BBA-Biomembranes 2006, 1758, 1653–1661. [Google Scholar] [CrossRef] [Green Version]
- Merlino, A.; Vitiello, G.; Grimaldi, M.; Sica, F.; Busi, E.; Basosi, R.; D’Ursi, A.M.; Fragneto, G.; Paduano, L.; D’Errico, G. Destabilization of lipid membranes by a peptide derived from glycoprotein gp36 of feline immunodeficiency virus: A combined molecular dynamics/experimental study. J. Phys. Chem. B 2011, 116, 401–412. [Google Scholar] [CrossRef]
- Giannecchini, S.; Alcaro, M.C.; Isola, P.; Sichi, O.; Pistello, M.; Papini, A.M.; Rovero, P.; Bendinelli, M. Feline immunodeficiency virus plasma load reduction by a retroinverso octapeptide reproducing the Trp-rich motif of the transmembrane glycoprotein. Antivir. Ther. 2005, 10, 671–680. [Google Scholar] [PubMed]
- Oliva, R.; Del Vecchio, P.; Stellato, M.I.; D’Ursi, A.M.; D’Errico, G.; Paduano, L.; Petraccone, L. A thermodynamic signature of lipid segregation in biomembranes induced by a short peptide derived from glycoprotein gp36 of feline immunodeficiency virus. BBA Biomembr. 2015, 1848, 510–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.; Alcaro, M.C.; Scrima, M.; Peroni, E.; Paolini, I.; Di Marino, S.; Barbetti, F.; Carotenuto, A.; Novellino, E.; Papini, A.M.; et al. Designed glucopeptides mimetics of myelin protein epitopes as synthetic probes for the detection of autoantibodies, biomarkers of multiple sclerosis. J. Med. Chem. 2012, 55, 10437–10447. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, M.; Stillitano, I.; Amodio, G.; Santoro, A.; Buonocore, M.; Moltedo, O.; Remondelli, P.; D’Ursi, A.M. Structural basis of antiviral activity of peptides from MPER of FIV gp36. PLoS ONE 2018, 13, e0204042. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.E.; Perkins, R.; Zschunke, M.; Hoerl, B.; Maercklein, P. Plasma membrane vesiculation in 3T3 and SV3T3 cells. I. Morphological and biochemical characterization. J. Cell Sci. 1979, 35, 229–243. [Google Scholar]
- Del Piccolo, N.; Placone, J.; He, L.; Agudelo, S.C.; Hristova, K. Production of plasma membrane vesicles with chloride salts and their utility as a cell membrane mimetic for biophysical characterization of membrane protein interactions. Anal. Chem. 2012, 84, 8650–8655. [Google Scholar] [CrossRef] [Green Version]
- Scott, R.E. Plasma membrane vesiculation: A new technique for isolation of plasma membranes. Science 1976, 194, 743–745. [Google Scholar] [CrossRef]
- Scott, R.E.; Maercklein, P.B. Plasma membrane vesiculation in 3T3 and SV3T3 cells. II. Factors affecting the process of vesiculation. J. Cell Sci. 1979, 35, 245–252. [Google Scholar]
- Mannhold, R.; Kubinyi, H.; Folkers, G.; Zerbe, O. BioNMR in Drug Research; Wiley-VCH: Zurich, Switzerland, 2006; Volume 16. [Google Scholar]
- Zerbe, O. BioNMR in Drug Research Methods and Principles in Medicinal Chemistry; Wiley-VCH: Zurich, Switzerland, 2003. [Google Scholar]
- Whitmore, L.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, W668–W673. [Google Scholar] [CrossRef] [Green Version]
- Weller, K.; Lauber, S.; Lerch, M.; Renaud, A.; Merkle, H.P.; Zerbe, O. Biophysical and biological studies of end-group-modified derivatives of Pep-1. Biochemistry 2005, 44, 15799–15811. [Google Scholar] [CrossRef]
- Goddard, T.; Kneller, D. SPARKY 3. Ph.D. Thesis, University of California, San Francisco, CA, USA, 2004. [Google Scholar]
- Wüthrich, K. NMR with Proteins and Nucleic Acids. Europhys. News 1986, 17, 11–13. [Google Scholar] [CrossRef]
- Cavanagh, J.; Fairbrother, W.J.; Palmer, A.G., III; Skelton, N.J. Protein NMR Spectroscopy: Principles and Practice; Elsevier: Amsterdam, The Netherlands, 1995. [Google Scholar]
- Clore, G.M.; Gronenborn, A.M. NMR of Proteins; Macmillan International Higher Education: New York, NY, USA, 1993. [Google Scholar]
- Wishart, D.S.; Sykes, B.D. The 13 C chemical-shift index: A simple method for the identification of protein secondary structure using 13 C chemical-shift data. J. Biomol. NMR 1994, 4, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Kjaergaard, M.; Brander, S.; Poulsen, F.M. Random coil chemical shift for intrinsically disordered proteins: Effects of temperature and pH. J. Biomol. NMR 2011, 49, 139–149. [Google Scholar] [CrossRef]
- Shen, Y.; Delaglio, F.; Cornilescu, G.; Bax, A. TALOS+: A hybrid method for predicting protein backbone torsion angles from NMR chemical shifts. J. Biomol. NMR 2009, 44, 213–223. [Google Scholar] [CrossRef]
- Guntert, P. Automated NMR structure calculation with CYANA. Methods Mol. Biol. 2004, 278, 353–378. [Google Scholar]
- Dongre, R.; Folkers, G.E.; Gualerzi, C.O.; Boelens, R.; Wienk, H. A model for the interaction of the G3-subdomain of Geobacillus stearothermophilus IF2 with the 30S ribosomal subunit. Protein Sci. 2016, 25, 1722–1733. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, E.G.; Thornton, J.M. PROMOTIF-A program to identify and analyze structural motifs in proteins. Protein Sci. 1996, 5, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metcalfe, E.E.; Zamoon, J.; Thomas, D.D.; Veglia, G. 1H/15N heteronuclear NMR spectroscopy shows four dynamic domains for phospholamban reconstituted in dodecylphosphocholine micelles. Biophys. J. 2004, 87, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, M.; Scrima, M.; Esposito, C.; Vitiello, G.; Ramunno, A.; Limongelli, V.; D’Errico, G.; Novellino, E.; D’Ursi, A.M. Membrane charge dependent states of the β-amyloid fragment Aβ (16–35) with differently charged micelle aggregates. Biochim. Biophys. Acta BBA Biomembr. 2010, 1798, 660–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jesus, A.J.; Allen, T.W. The role of tryptophan side chains in membrane protein anchoring and hydrophobic mismatch. BBA Biomembr. 2013, 1828, 864–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.-Y.J.; Cheng, Y.; Kim, M.; Song, L.; Choi, J.; Kudahl, U.J.; Brusic, V.; Chowdhury, B.; Yu, L.; Seaman, M.S. Disruption of helix-capping residues 671 and 674 reveals a role in HIV-1 entry for a specialized hinge segment of the membrane proximal external region of gp41. J. Mol. Biol. 2014, 426, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reardon, P.N.; Sage, H.; Dennison, S.M.; Martin, J.W.; Donald, B.R.; Alam, S.M.; Haynes, B.F.; Spicer, L.D. Structure of an HIV-1–neutralizing antibody target, the lipid-bound gp41 envelope membrane proximal region trimer. Proc. Natl. Acad. Sci. USA 2014, 111, 1391–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.-Y.J.; Oh, K.J.; Kim, M.; Yu, J.; Brusic, V.; Song, L.; Qiao, Z.; Wang, J.-H.; Wagner, G.; Reinherz, E.L. HIV-1 broadly neutralizing antibody extracts its epitope from a kinked gp41 ectodomain region on the viral membrane. Immunity 2008, 28, 52–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, B.; Lee, M.; Waring, A.J.; Hong, M. Oligomeric structure and three-dimensional fold of the HIV gp41 membrane-proximal external region and transmembrane domain in phospholipid bilayers. J. Am. Chem. Soc. 2018, 140, 8246–8259. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Nyenhuis, D.A.; Nelson, E.A.; Cafiso, D.S.; White, J.M.; Tamm, L.K. Structure of the Ebola virus envelope protein MPER/TM domain and its interaction with the fusion loop explains their fusion activity. Proc. Natl. Acad. Sci. USA 2017, 114, E7987–E7996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuliopulos, A.; Walsh, C.T. Production, purification, and cleavage of tandem repeats of recombinant peptides. J. Am. Chem. Soc. 1994, 116, 4599–4607. [Google Scholar] [CrossRef]
- Maniatis, T. Molecular cloning. In Decontamination of Dilute Solutions of Ethidium Bromide; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Rodriguez, J.C.; Wong, L.; Jennings, P.A. The solvent in CNBr cleavage reactions determines the fragmentation efficiency of ketosteroid isomerase fusion proteins used in the production of recombinant peptides. Protein Expr. Purif. 2003, 28, 224–231. [Google Scholar] [CrossRef]
- Pellegrini, M.; Mierke, D.F. Structural characterization of peptide hormone/receptor interactions by NMR spectroscopy. Pept. Sci. 1999, 51, 208–220. [Google Scholar] [CrossRef]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Tedeschi, A.; Franco, L.; Ruzzi, M.; Paduano, L.; Corvaja, C.; D’Errico, G. Micellar aggregation of alkyltrimethylammonium bromide surfactants studied by electron paramagnetic resonance of an anionic nitroxide. Phys. Chem. Chem. Phys. 2003, 5, 4204–4209. [Google Scholar] [CrossRef]
- Moltedo, O.; Remondelli, P.; Amodio, G. The Mitochondria-Endoplasmic Reticulum Contacts and their critical role in ageing and age-associated diseases. Front. Cell Dev. Biol. 2019, 7, 172. [Google Scholar] [CrossRef] [PubMed]
- Vitiello, G.; Falanga, A.; Petruk, A.A.; Merlino, A.; Fragneto, G.; Paduano, L.; Galdiero, S.; D’Errico, G. Fusion of raft-like lipid bilayers operated by a membranotropic domain of the HSV-type I glycoprotein gH occurs through a cholesterol-dependent mechanism. Soft Matter 2015, 11, 3003–3016. [Google Scholar] [CrossRef] [PubMed]
- Caggiano, R.; Cattaneo, F.; Moltedo, O.; Esposito, G.; Perrino, C.; Trimarco, B.; Ammendola, R.; Faraonio, R. miR-128 is implicated in stress responses by targeting MAFG in skeletal muscle cells. Oxid. Med. Cell. Longev. 2017, 2017, 9308310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amodio, G.; Moltedo, O.; Fasano, D.; Zerillo, L.; Oliveti, M.; Di Pietro, P.; Faraonio, R.; Barone, P.; Pellecchia, M.T.; De Rosa, A. PERK-mediated unfolded protein response activation and oxidative stress in PARK20 fibroblasts. Front. Neurosci. 2019, 13, 673. [Google Scholar] [CrossRef] [Green Version]
- Fasano, D.; Parisi, S.; Pierantoni, G.M.; De Rosa, A.; Picillo, M.; Amodio, G.; Pellecchia, M.T.; Barone, P.; Moltedo, O.; Bonifati, V.; et al. Alteration of endosomal trafficking is associated with early-onset parkinsonism caused by SYNJ1 mutations. Cell Death Dis. 2018, 9, 385. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of NOESY Peaks | |
| Total | 457 |
| Number of Experimental Restraints after CYANA | |
| Total NOEs | 445 |
| Intra-residual | 294 |
| Sequential | 122 |
| Medium-range | 20 |
| Long-range | 9 |
| Dihedral angles | 73 |
| RMSD | |
| bb/heavy Å | 1.38/1.98 |
| Ramachandran Analysis | |
| Favourable regions | 78.40% |
| Additionally allowed regions | 19.30% |
| Generously allowed regions | 2.30% |
| Disallowed regions | 0% |
| AN/G | τC × 1010/s | 2Amax/G | |
|---|---|---|---|
| 5-DSA | |||
| DPC/SDS 90:10 | 14.5 ± 0.1 | 27 ± 2 | - |
| DPC/SDS 90:10 + 737-786gp36 CHR–MPER | 14.6 ± 0.1 | 42 ± 3 | - |
| 5-PCSL | |||
| DOPC/DOPG 90:10 | - | - | 51.2 ± 0.1 |
| DOPC/DOPG 90:10 + 737-786gp36 CHR–MPER | - | - | 52.4 ± 0.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grimaldi, M.; Buonocore, M.; Scrima, M.; Stillitano, I.; D’Errico, G.; Santoro, A.; Amodio, G.; Eletto, D.; Gloria, A.; Russo, T.; et al. NMR Structure of the FIV gp36 C-terminal Heptad Repeat and Membrane-Proximal External Region. Int. J. Mol. Sci. 2020, 21, 2037. https://doi.org/10.3390/ijms21062037
Grimaldi M, Buonocore M, Scrima M, Stillitano I, D’Errico G, Santoro A, Amodio G, Eletto D, Gloria A, Russo T, et al. NMR Structure of the FIV gp36 C-terminal Heptad Repeat and Membrane-Proximal External Region. International Journal of Molecular Sciences. 2020; 21(6):2037. https://doi.org/10.3390/ijms21062037
Chicago/Turabian StyleGrimaldi, Manuela, Michela Buonocore, Mario Scrima, Ilaria Stillitano, Gerardino D’Errico, Angelo Santoro, Giuseppina Amodio, Daniela Eletto, Antonio Gloria, Teresa Russo, and et al. 2020. "NMR Structure of the FIV gp36 C-terminal Heptad Repeat and Membrane-Proximal External Region" International Journal of Molecular Sciences 21, no. 6: 2037. https://doi.org/10.3390/ijms21062037