Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles


 ,
,  , ,
, ,  and
and 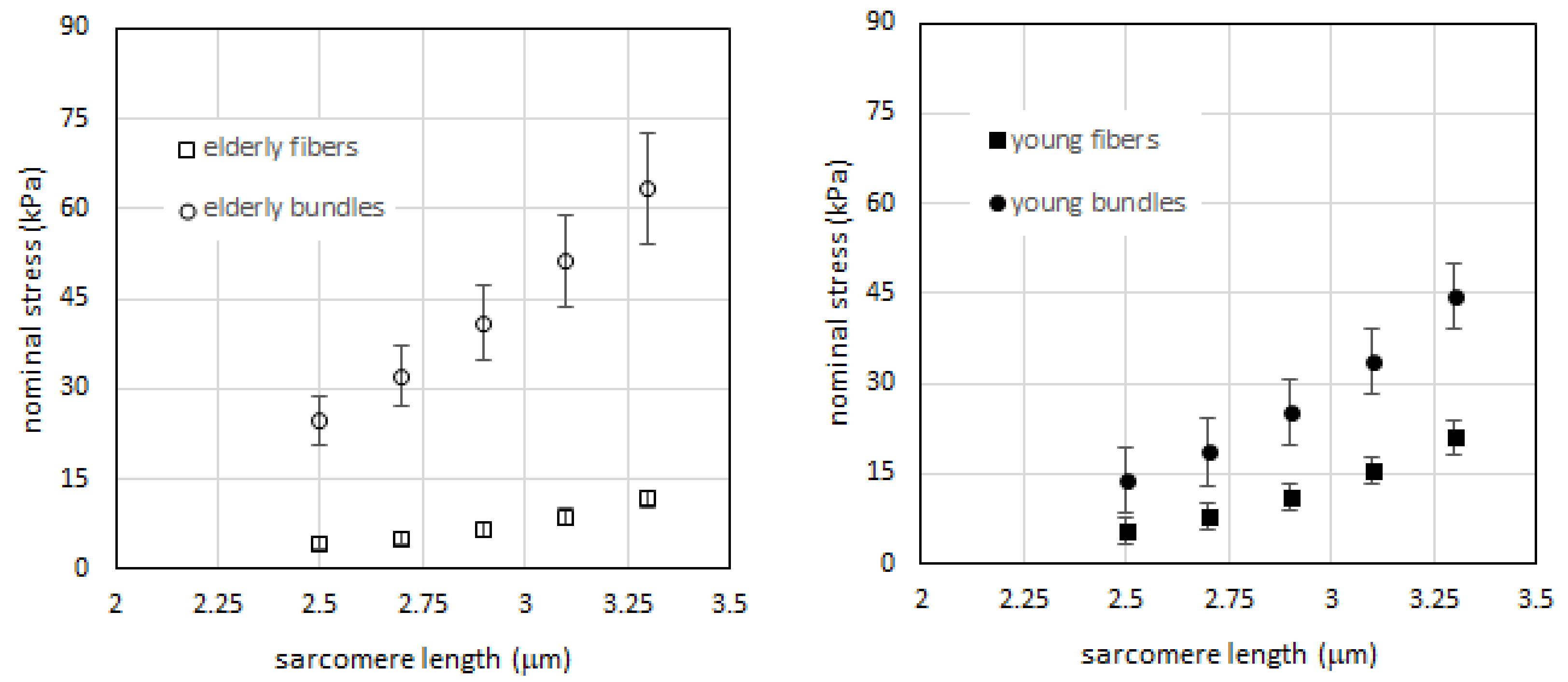{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
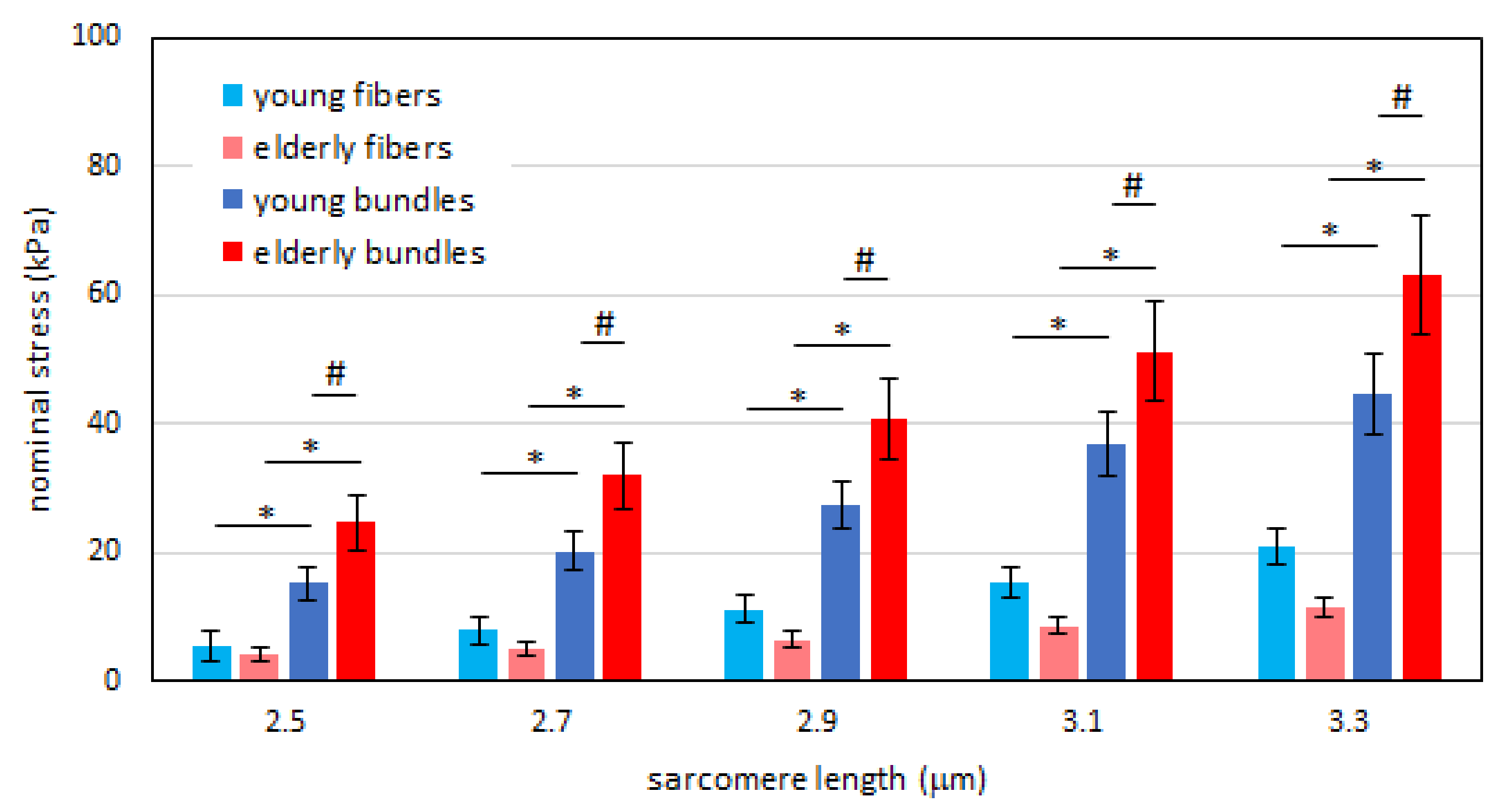
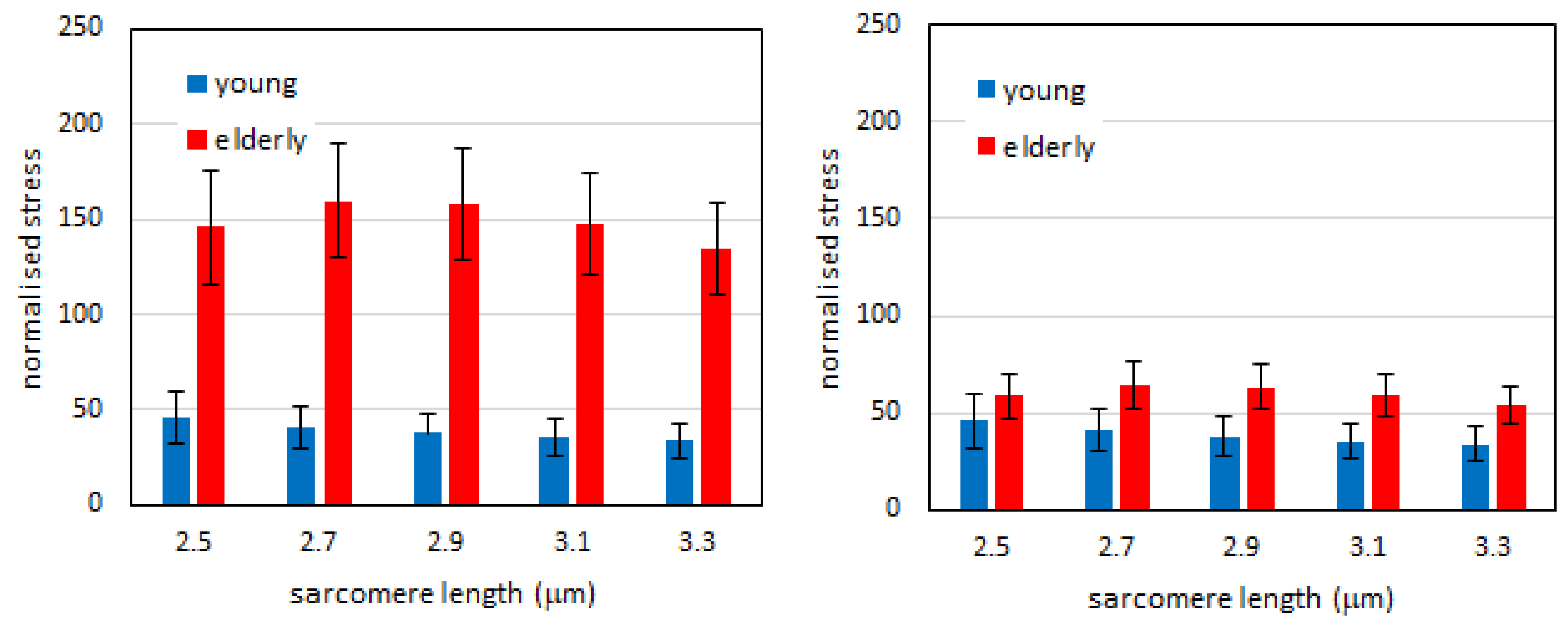
2.1. Passive Tension is Higher in Bundles than in Single Fibers in Both Young and Elderly Subjects
2.2. Passive Tension in Young Compared to Elderly Subjects Is Significantly Different in Bundles but Not in Single Muscle Fibers
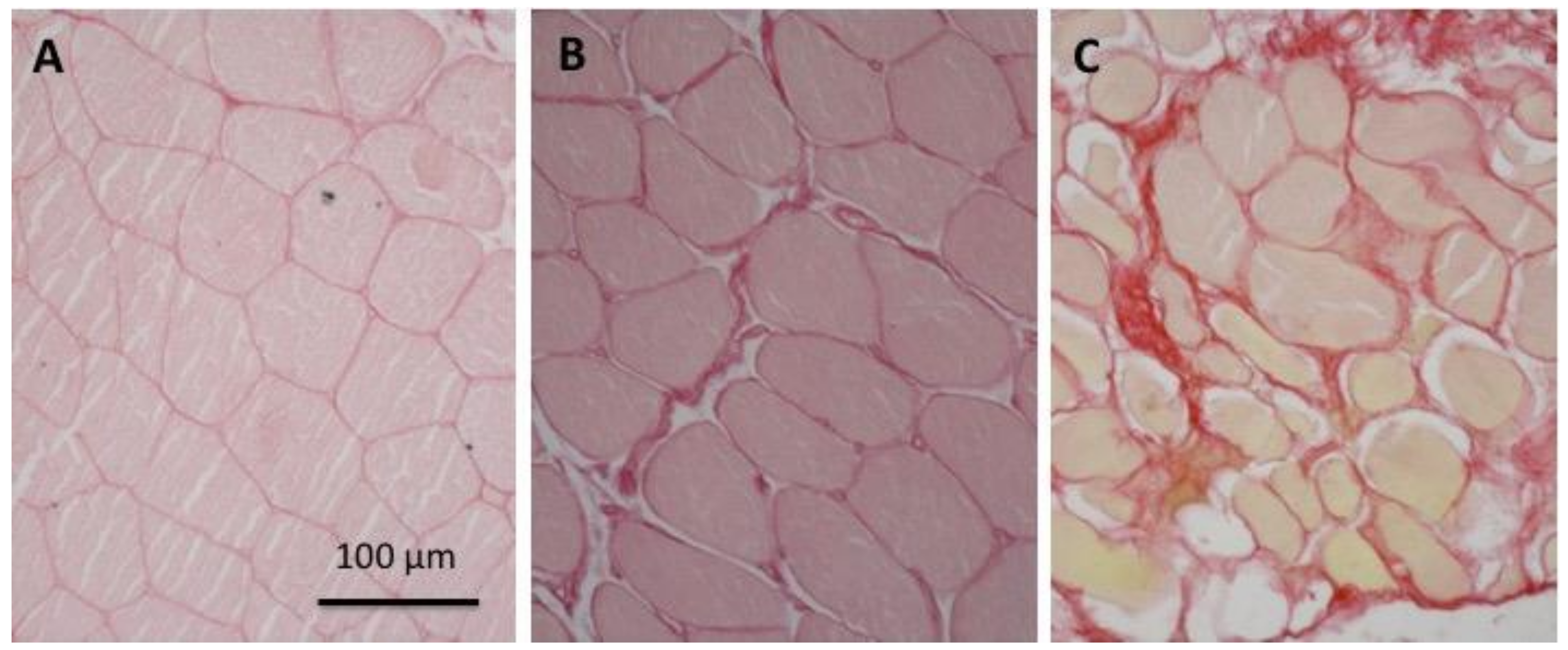
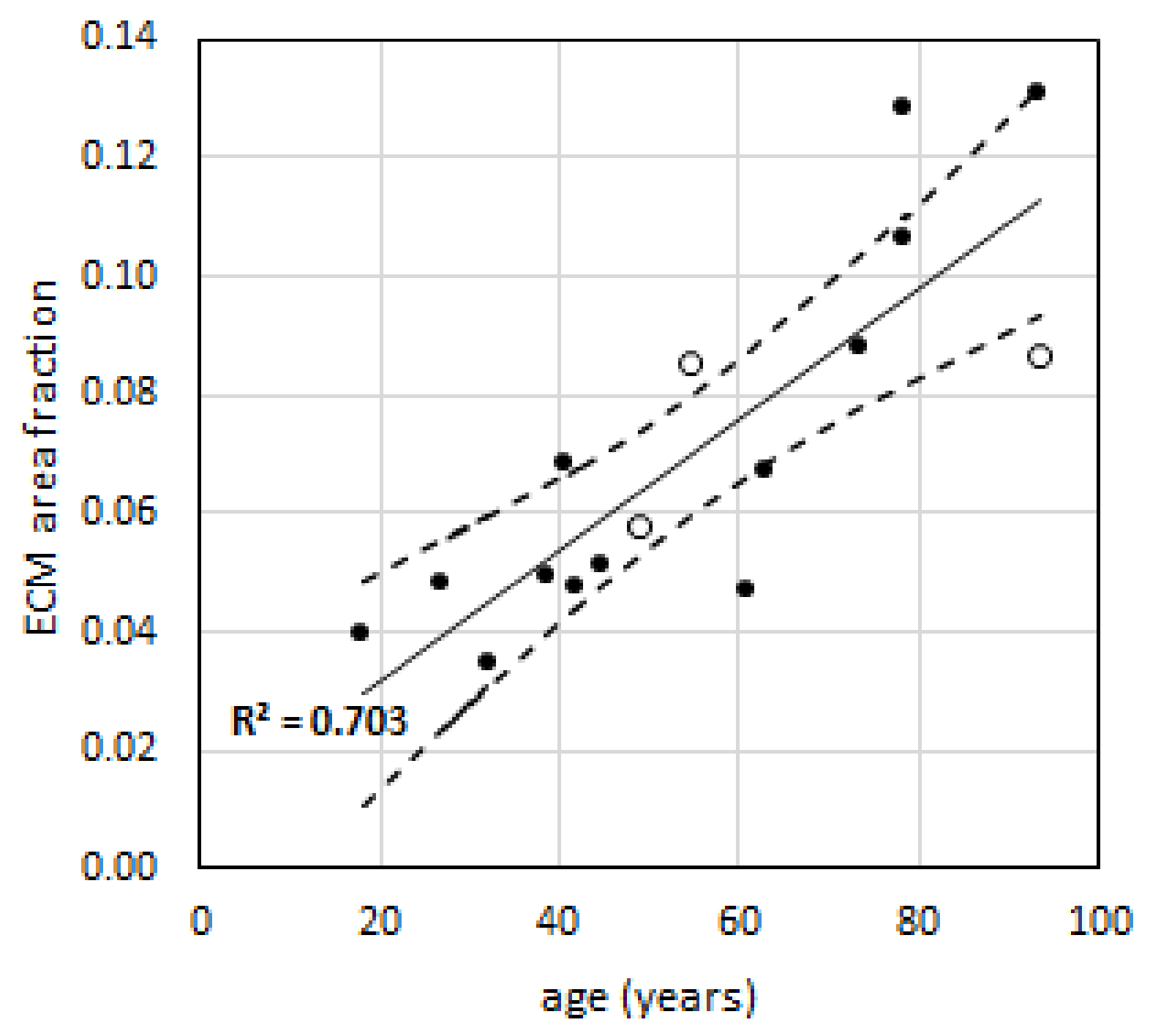
2.3. Relative Amount of Extra-Cellular Matrix is Higher in Elderly Than in Young Subjects
2.4. Extracellular Matrix Stiffness Is Not Significantly Different between Elderly or Young Subjects
3. Discussion
4. Experimental Procedures
4.1. Participants
4.2. Experimental Protocol
4.3. Histological Analysis
4.4. Tension Calculation at Each Elongation Step
4.5. Statistical Analysis
4.6. Ethics approval
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CSA | cross sectional area |
| E | elderly subjects |
| ECM | extracellular matrix |
| SL | sarcomere length |
| Y | young subjects |
Data Availability Statement
Permission to Reproduce Material from Other Sources
Clinical Trial Registration
References
- Gajdosik, R.L. Influence of Age on Calf Muscle Length and Passive Stiffness Variables at Different Stretch Velocities. Isokinet. Exerc. Sci. 1997, 6, 163–174. [Google Scholar] [CrossRef]
- Gajdosik, R.L. Passive extensibility of skeletal muscle: Review of the literature with clinical implications. Clin Biomech 2001, 16, 87–101. [Google Scholar] [CrossRef]
- Boyer, K.A.; Johnson, R.T.; Banks, J.J.; Jewell, C.; Hafer, J.F. Systematic review and meta-analysis of gait mechanics in young and older adults. Exp. Gerontol. 2017, 95, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Oosterwijk, A.M.; Nieuwenhuis, M.K.; van der Schans, C.P.; Mouton, L.J. Shoulder and elbow range of motion for the performance of activities of daily living: A systematic review. Physiother. Theory Pract. 2018, 34, 505–528. [Google Scholar] [CrossRef]
- Reeves, N.D.; Narici, M.V.; Maganaris, C.N. Myotendinous plasticity to ageing and resistance exercise in humans. Exp. Physiol. 2006, 91, 483–498. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, W.K.; Williams, J.; Atherton, P.; Larvin, M.; Lund, J.; Narici, M. Sarcopenia, Dynapenia, and the Impact of Advancing Age on Human Skeletal Muscle Size and Strength; a Quantitative Review. Front Physiol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.J.; Fielding, R.A. Neuromuscular contributions to age-related weakness. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Ochala, J.; Lambertz, D.; Pousson, M.; Goubel, F.; Hoecke, J.V. Changes in mechanical properties of human plantar flexor muscles in ageing. Exp. Gerontol. 2004, 39, 349–358. [Google Scholar] [CrossRef]
- Ochala, J.; Lambertz, D.; Van Hoecke, J.; Pousson, M. Changes in muscle and joint elasticity following long-term strength training in old age. Eur. J. Appl. Physiol. 2007, 100, 491–498. [Google Scholar] [CrossRef]
- Maganaris, C.N.; Narici, M.V.; Reeves, N.D. In vivo human tendon mechanical properties: Effect of resistance training in old age. J. Musculoskelet Neuronal Interact. 2004, 4, 204–208. [Google Scholar]
- Onambele, G.L.; Narici, M.V.; Maganaris, C.N. Calf muscle-tendon properties and postural balance in old age. J. Appl. Physiol. 2006, 100, 2048–2056. [Google Scholar] [CrossRef] [PubMed]
- Roig, M.; Macintyre, D.L.; Eng, J.J.; Narici, M.V.; Maganaris, C.N.; Reid, W.D. Preservation of eccentric strength in older adults: Evidence, mechanisms and implications for training and rehabilitation. Exp. Gerontol. 2010, 45, 400–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alnaqeeb, M.A.; Al Zaid, N.S.; Goldspink, G. Connective tissue changes and physical properties of developing and ageing skeletal muscle. J. Anat. 1984, 139, 677–689. [Google Scholar]
- Stearns-Reider, K.M.; D’Amore, A.; Beezhold, K.; Rothrauff, B.; Cavalli, L.; Wagner, W.R.; Vorp, D.A.; Tsamis, A.; Shinde, S.; Zhang, C.; et al. Aging of the skeletal muscle extracellular matrix drives a stem cell fibrogenic conversion. Aging Cell 2017, 16, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Wood, L.K.; Kayupov, E.; Gumucio, J.P.; Mendias, C.L.; Claflin, D.R.; Brooks, S.V. Intrinsic stiffness of extracellular matrix increases with age in skeletal muscles of mice. J. Appl. Physiol. 2014, 117, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Huijing, P.A.; Baan, G.C.; Rebel, G.T. Non-myotendinous force transmission in rat extensor digitorum longus muscle. J. Exp. Biol. 1998, 201, 683–691. [Google Scholar]
- Ramaswamy, K.S.; Palmer, M.L.; van der Meulen, J.H.; Renoux, A.; Kostrominova, T.Y.; Michele, D.E.; Faulkner, J.A. Lateral transmission of force is impaired in skeletal muscles of dystrophic mice and very old rats. J. Physiol. 2011, 589, 1195–1208. [Google Scholar] [CrossRef]
- Street, S.F. Lateral transmission of tension in frog myofibers: A myofibrillar network and transverse cytoskeletal connections are possible transmitters. J. Cell. Physiol. 1983, 114, 346–364. [Google Scholar] [CrossRef]
- Moore, A.Z.; Caturegli, G.; Metter, E.J.; Makrogiannis, S.; Resnick, S.M.; Harris, T.B.; Ferrucci, L. Difference in muscle quality over the adult life span and biological correlates in the Baltimore Longitudinal Study of Aging. J. Am. Geriatr. Soc. 2014, 62, 230–236. [Google Scholar] [CrossRef]
- Meyer, G.; Lieber, R.L. Muscle fibers bear a larger fraction of passive muscle tension in frogs compared with mice. J. Exp. Biol. 2018, 221, jeb182089. [Google Scholar] [CrossRef] [Green Version]
- Meyer, G.A.; Lieber, R.L. Elucidation of extracellular matrix mechanics from muscle fibers and fiber bundles. J. Biomech. 2011, 44, 771–773. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-Y.; Choi, S.J.; Widrick, J.J.; Phillips, E.M.; Frontera, W.R. Passive force and viscoelastic properties of single fibers in human aging muscles. Graefe’s Arch. Clin. Exp. Ophthalmol. 2019, 119, 2339–2348. [Google Scholar] [CrossRef] [PubMed]
- Marcucci, L.; Bondì, M.; Randazzo, G.; Reggiani, C.; Natali, A.N.; Pavan, P.G. Fibre and extracellular matrix contributions to passive forces in human skeletal muscles: An experimental based constitutive law for numerical modelling of the passive element in the classical Hill-type three element model. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horowits, R. Passive force generation and titin isoforms in mammalian skeletal muscle. Biophys. J. 1992, 61, 392–398. [Google Scholar] [CrossRef] [Green Version]
- Olsson, M.C.; Krüger, M.; Meyer, L.-H.; Ahnlund, L.; Gransberg, L.; Linke, W.A.; Larsson, L. Fibre type-specific increase in passive muscle tension in spinal cord-injured subjects with spasticity. J. Physiol. 2006, 577, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.R.; Winters, T.M.; O’Connor, S.M.; Lieber, R.L. Non-linear Scaling of Passive Mechanical Properties in Fibers, Bundles, Fascicles and Whole Rabbit Muscles. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.R.; Lee, K.S.; Ward, S.R.; Chambers, H.G.; Lieber, R.L. Hamstring contractures in children with spastic cerebral palsy result from a stiffer extracellular matrix and increased in vivo sarcomere length. J. Physil. 2011, 589, 2625–2639. [Google Scholar] [CrossRef]
- Gosselin, L.E.; Adams, C.; Cotter, T.A.; McCormick, R.J.; Thomas, D.P. Effect of exercise training on passive stiffness in locomotor skeletal muscle: Role of extracellular matrix. J. Appl. Physiol. 1998, 85, 1011–1016. [Google Scholar] [CrossRef]
- Brown, M.; Fisher, J.S.; Salsich, G. Stiffness and muscle function with age and reduced muscle use. J. Orthop. Res. 1999, 17, 409–414. [Google Scholar] [CrossRef]
- Blanpied, P.; Smidt, G.L. The difference in stiffness of the active plantarflexors between young and elderly human females. J. Gerontol. 1993, 48, M58–M63. [Google Scholar] [CrossRef]
- Sobolewski, E.J.; Ryan, E.D.; Thompson, B.J.; McHugh, M.P.; Conchola, E.C. The influence of age on the viscoelastic stretch response. J. Strength Cond. Res. 2014, 28, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Palmer, T.B.; Thompson, B.J. Influence of age on passive stiffness and size, quality, and strength characteristics. Muscle Nerve 2017, 55, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Valour, D.; Pousson, M. Compliance changes of the series elastic component of elbow flexor muscles with age in humans. Pflugers Arch. 2003, 445, 721–727. [Google Scholar] [CrossRef]
- Linke, W.A. Titin Gene and Protein Functions in Passive and Active Muscle. Annu. Rev.Physiol. 2018, 80, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado, L.G.; Makarenko, I.; Andresen, C.; Krüger, M.; Opitz, C.A.; Linke, W.A. Isoform diversity of giant proteins in relation to passive and active contractile properties of rabbit skeletal muscles. J. Gen. Physiol. 2005, 126, 461–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; McCarter, R.; Wright, J.; Beverly, J.; Ramirez-Mitchell, R. Regulation of skeletal muscle stiffness and elasticity by titin isoforms: A test of the segmental extension model of resting tension. Proc. Natl Acad. Sci. USA 1991, 88, 7101–7105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freundt, J.K.; Linke, W.A. Titin as a force-generating muscle protein under regulatory control. J. Appl. Physiol. 2019, 126, 1474–1482. [Google Scholar] [CrossRef] [PubMed]
- Labeit, D.; Watanabe, K.; Witt, C.; Fujita, H.; Wu, Y.; Lahmers, S.; Funck, T.; Labeit, S.; Granzier, H. Calcium-dependent molecular spring elements in the giant protein titin. Proc. Natl. Acad. Sci. USA. 2003, 100, 13716–13721. [Google Scholar] [CrossRef] [Green Version]
- Kjaer, M. Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol. Rev. 2004, 84, 649–698. [Google Scholar] [CrossRef]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junqueira, L.C.U.; Cossermelli, W.; Brentani, R. Differential Staining of Collagens Type I, II and III by Sirius Red and Polarization Microscopy. Arch. Histol. Jap., Arch. Histol. Jpn 1978, 41, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcucci, L.; Reggiani, C. Mechanosensing in Myosin Filament Solves a 60 Years Old Conflict in Skeletal Muscle Modeling between High Power Output and Slow Rise in Tension. Front Physiol 2016, 7, 427. [Google Scholar] [CrossRef] [Green Version]
- Marcucci, L.; Washio, T.; Yanagida, T. Titin-mediated thick filament activation, through a mechanosensing mechanism, introduces sarcomere-length dependencies in mathematical models of rat trabecula and whole ventricle. Scientific Reports 2017, 7, 5546. [Google Scholar] [CrossRef] [Green Version]
- Marcucci, L.; Washio, T.; Yanagida, T. Including thermal fluctuations in actomyosin stable states increases the predicted force per motor and macroscopic efficiency in muscle modelling. PLoS Comput. Biol. 2016, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huijing, P.A. Muscle as a collagen fiber reinforced composite: A review of force transmission in muscle and whole limb. J. Biomech. 1999, 32, 329–345. [Google Scholar] [CrossRef]
- Blemker, S.S.; Pinsky, P.M.; Delp, S.L. A 3D model of muscle reveals the causes of nonuniform strains in the biceps brachii. J. Biomech. 2005, 38, 657–665. [Google Scholar] [CrossRef]
- Bleiler, C.; Ponte Castañeda, P.; Röhrle, O. A microstructurally-based, multi-scale, continuum-mechanical model for the passive behaviour of skeletal muscle tissue. J. Mech. Behav. Biomed. Mater. 2019, 97, 171–186. [Google Scholar] [CrossRef]
- Sharafi, B.; Blemker, S.S. A mathematical model of force transmission from intrafascicularly terminating muscle fibers. J. Biomech. 2011, 44, 2031–2039. [Google Scholar] [CrossRef] [Green Version]
- Teklemariam, A.; Hodson-Tole, E.; Reeves, N.D.; Cooper, G. A micromechanical muscle model for determining the impact of motor unit fiber clustering on force transmission in aging skeletal muscle. Biomech. Model Mechanobiol. 2019, 18, 1401–1413. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Gao, Y. Finite element analysis of mechanics of lateral transmission of force in single muscle fiber. J. Biomech. 2012, 45, 2001–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, J.-S.; He, Q.; He, X.; Basava, R.R.; Hodgson, J.; Sinha, U.; Sinha, S. Microstructural analysis of skeletal muscle force generation during aging. Int. J. Numer. Method Biomed. Eng 2020, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcucci, L.; Reggiani, C.; Natali, A.N.; Pavan, P.G. From single muscle fiber to whole muscle mechanics: A finite element model of a muscle bundle with fast and slow fibers. Biomec. Model. Mechan. 2017, 16, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- Virgilio, K.M.; Martin, K.S.; Peirce, S.M.; Blemker, S.S. Multiscale models of skeletal muscle reveal the complex effects of muscular dystrophy on tissue mechanics and damage susceptibility. Interface Focus 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wineman, A.S.; Waas, A.M. Mechanics of muscle injury induced by lengthening contraction. Ann. Biomed. Eng. 2008, 36, 1615–1623. [Google Scholar] [CrossRef]
- Harridge, S.D.R.; Lazarus, N.R. Physical Activity, Aging, and Physiological Function. Physiology 2017, 32, 152–161. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, S.D.; McCormick, R.J.; Vadlamudi, R.K.; Thomas, D.P. Age and training alter collagen characteristics in fast- and slow-twitch rat limb muscle. J. Appl. Physiol. 1993, 75, 1670–1674. [Google Scholar] [CrossRef]
- Kovanen, V.; Suominen, H.; Peltonen, L. Effects of aging and life-long physical training on collagen in slow and fast skeletal muscle in rats. A morphometric and immuno-histochemical study. Cell Tissue Res. 1987, 248, 247–255. [Google Scholar] [CrossRef]
- Rejc, E.; Floreani, M.; Taboga, P.; Botter, A.; Toniolo, L.; Cancellara, L.; Narici, M.; Šimunič, B.; Pišot, R.; Biolo, G.; et al. Loss of maximal explosive power of lower limbs after 2 weeks of disuse and incomplete recovery after retraining in older adults. J. Physiol. 2018, 596, 647–665. [Google Scholar] [CrossRef]
- Chen, X.; Sanchez, G.N.; Schnitzer, M.J.; Delp, S.L. Changes in sarcomere lengths of the human vastus lateralis muscle with knee flexion measured using in vivo microendoscopy. J. Biomech. 2016, 49, 2989–2994. [Google Scholar] [CrossRef] [Green Version]
- Lieber, R.L.; Runesson, E.; Einarsson, F.; Fridén, J. Inferior mechanical properties of spastic muscle bundles due to hypertrophic but compromised extracellular matrix material. Muscle Nerve 2003, 28, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Caetano, G.F.; Frade, M.A.C.; Andrade, T.A.M.; Leite, M.N.; Bueno, C.Z.; Moraes, Â.M.; Ribeiro-Paes, J.T. Chitosan-alginate membranes accelerate wound healing. J. Biomed. Mater. Res. Part B Appl. Biomater. 2015, 103, 1013–1022. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavan, P.; Monti, E.; Bondí, M.; Fan, C.; Stecco, C.; Narici, M.; Reggiani, C.; Marcucci, L. Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles. Int. J. Mol. Sci. 2020, 21, 3992. https://doi.org/10.3390/ijms21113992
Pavan P, Monti E, Bondí M, Fan C, Stecco C, Narici M, Reggiani C, Marcucci L. Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles. International Journal of Molecular Sciences. 2020; 21(11):3992. https://doi.org/10.3390/ijms21113992
Chicago/Turabian StylePavan, Piero, Elena Monti, Michela Bondí, Chenglei Fan, Carla Stecco, Marco Narici, Carlo Reggiani, and Lorenzo Marcucci. 2020. "Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles" International Journal of Molecular Sciences 21, no. 11: 3992. https://doi.org/10.3390/ijms21113992