Wood-Based Cellulose Nanofibrils: Haemocompatibility and Impact on the Development and Behaviour of Drosophila melanogaster

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
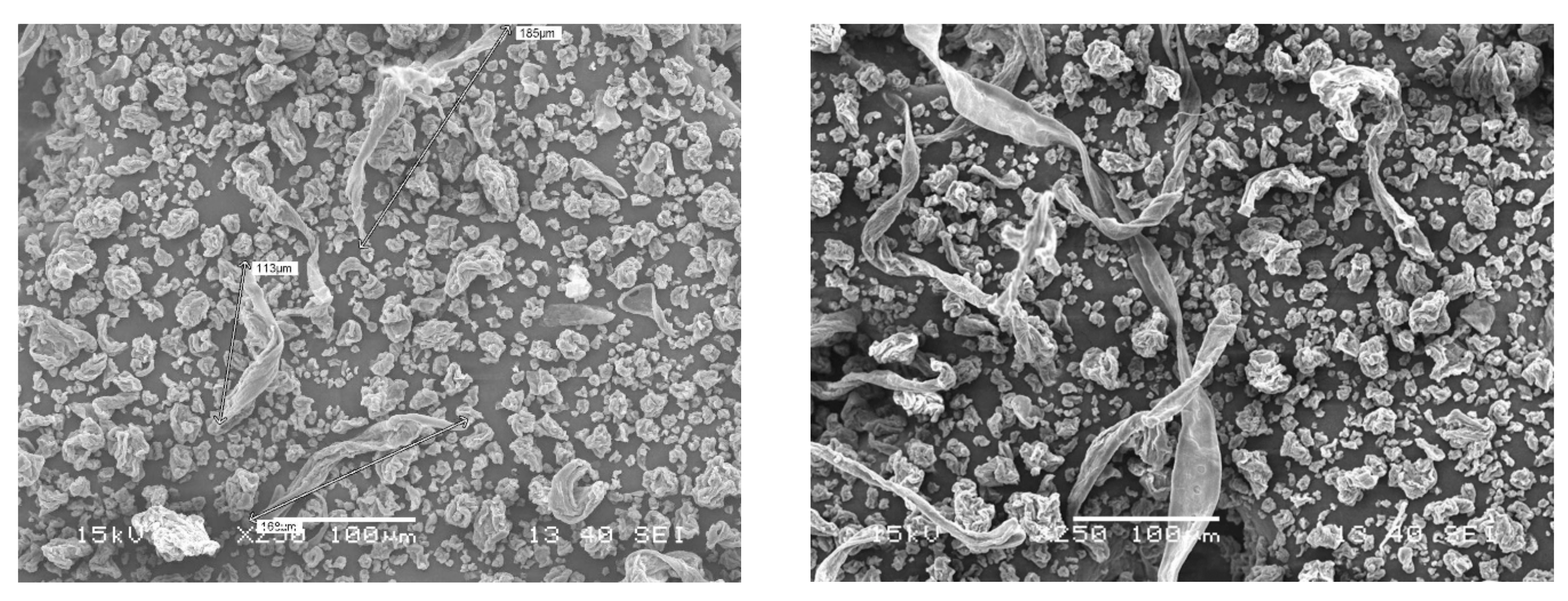
2.1. Materials
2.2. Fly Management
2.3. Treatment
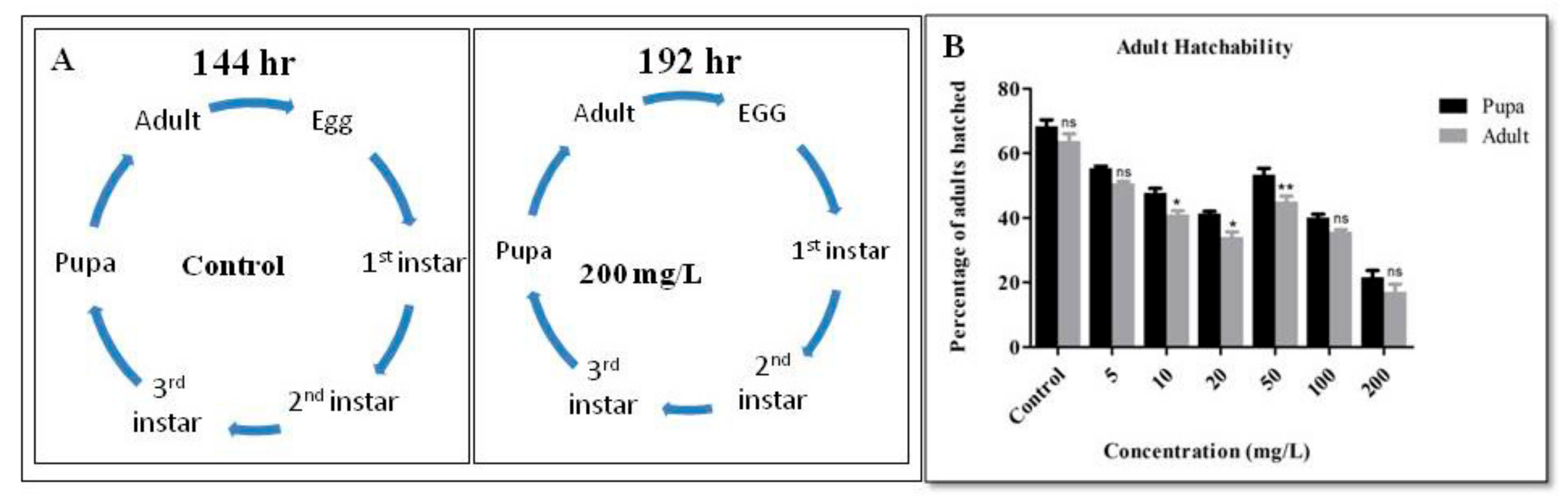
2.4. Developmental Cycle
2.5. Trypan Blue Exclusion Assay
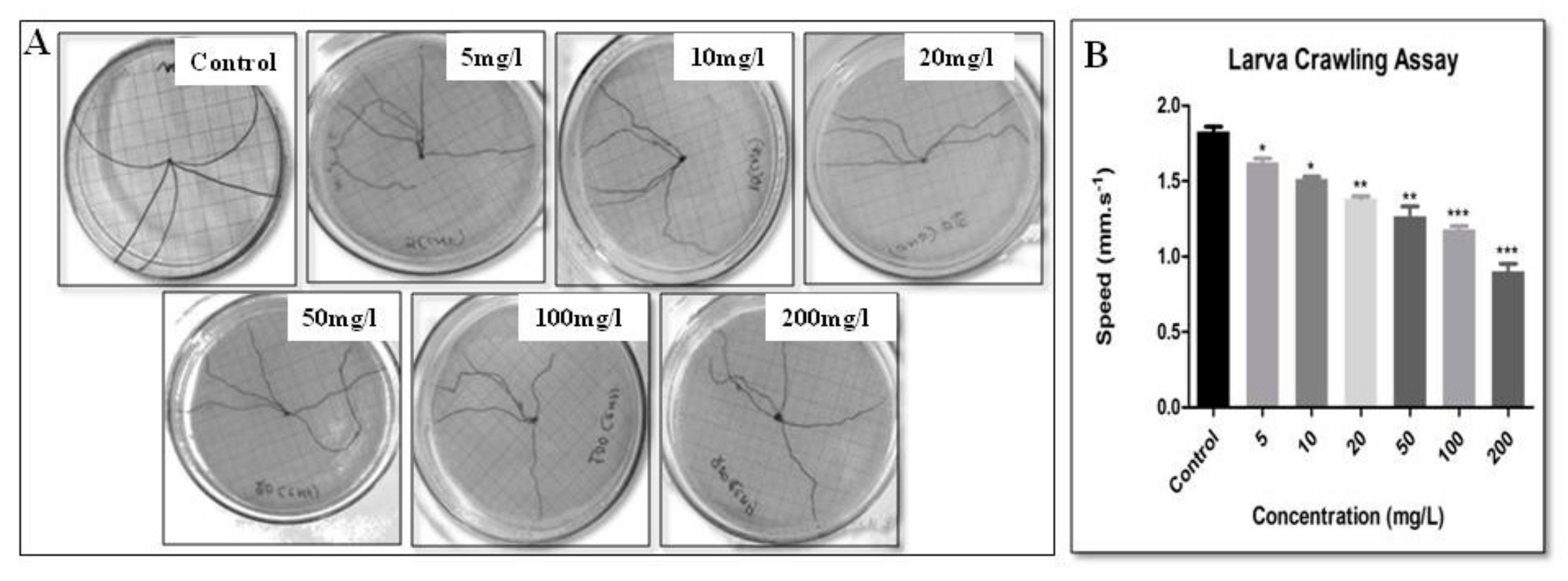
2.6. Larva Crawling Assay
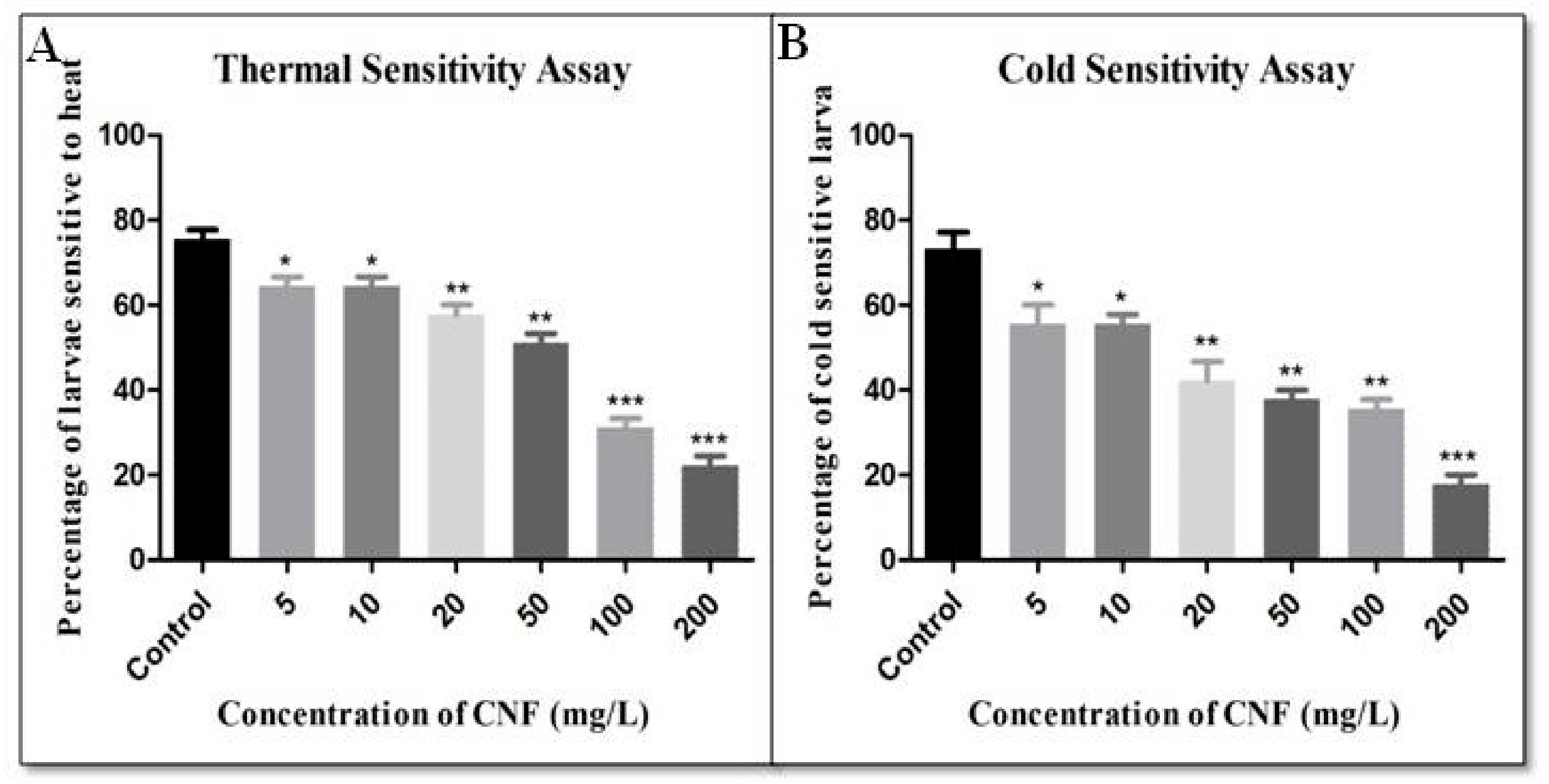
2.7. Thermal Sensitivity Assay
2.8. Cold Sensitivity Assay
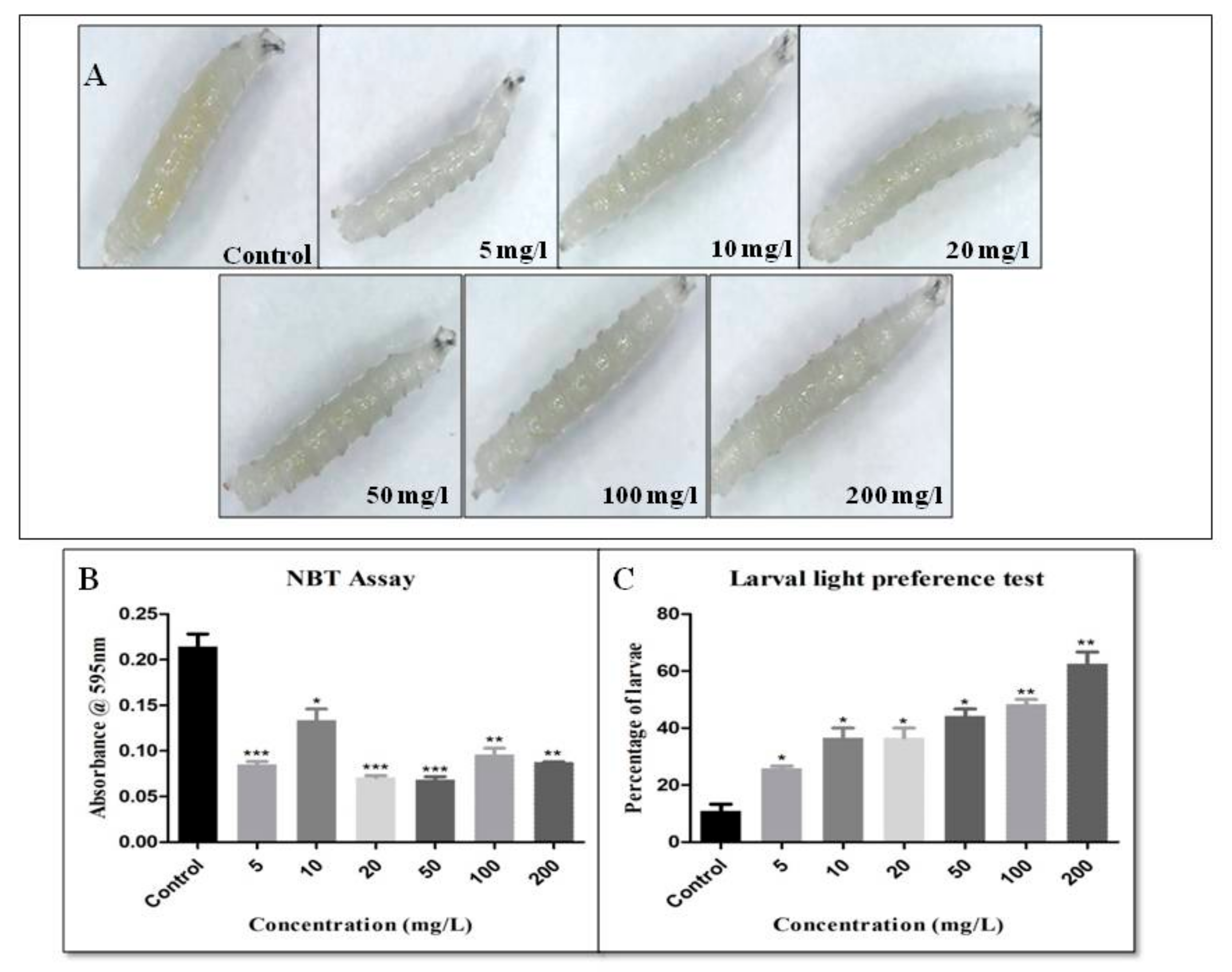
2.9. Larval Light Preference Test
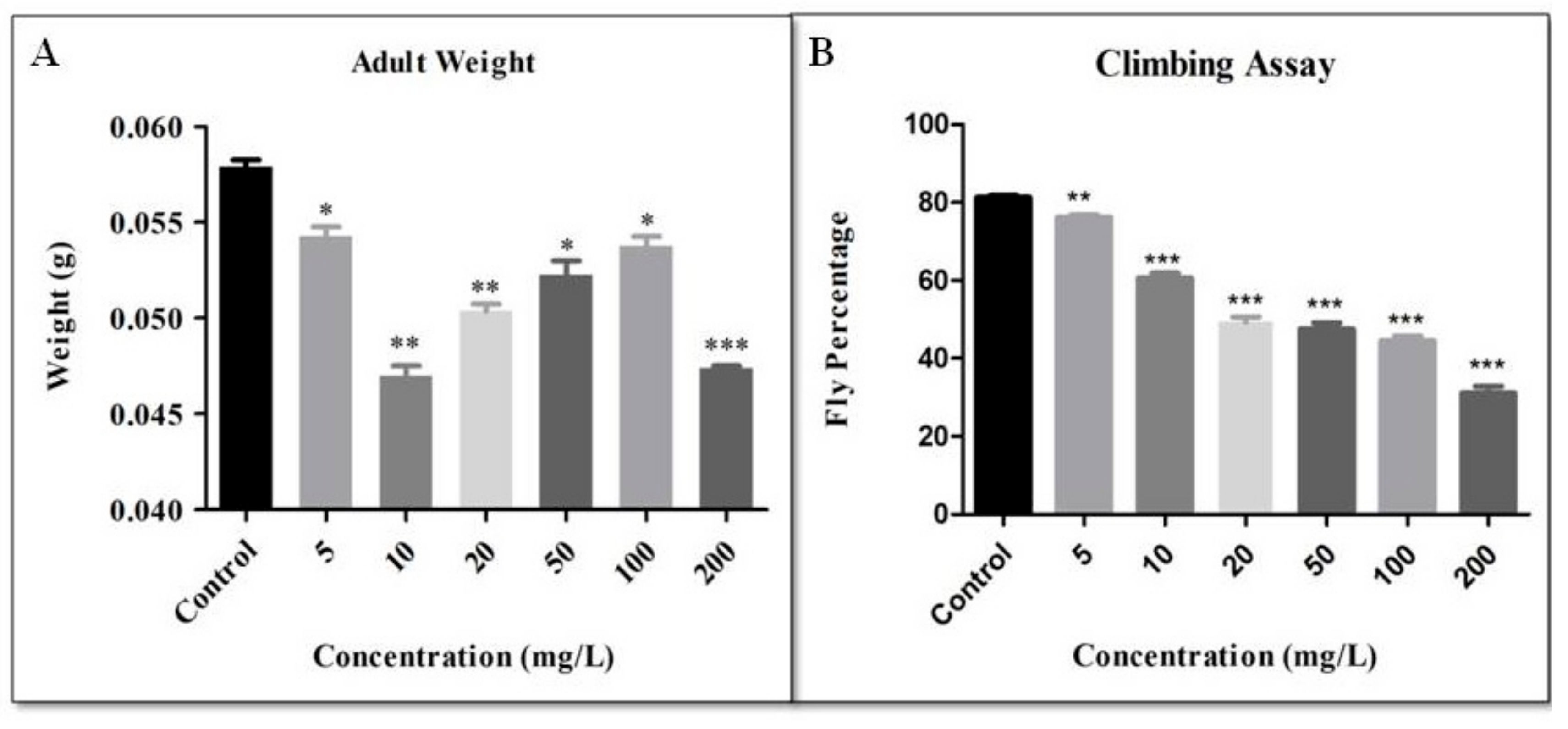
2.10. Climbing Behaviour
2.11. Nitroblue Tetrazolium Reduction Assay
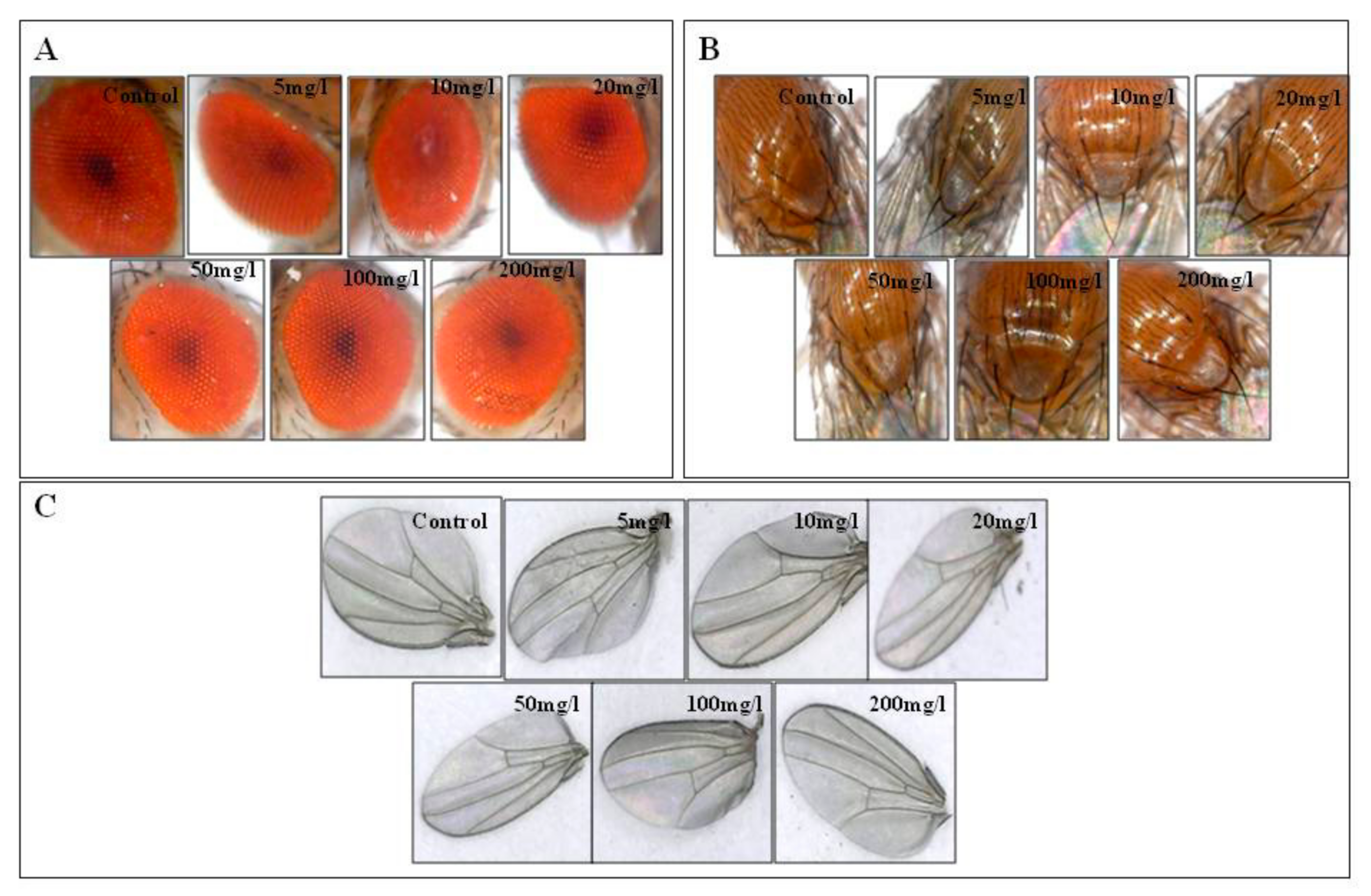
2.12. Adult Phenotype
2.13. Adult Weight
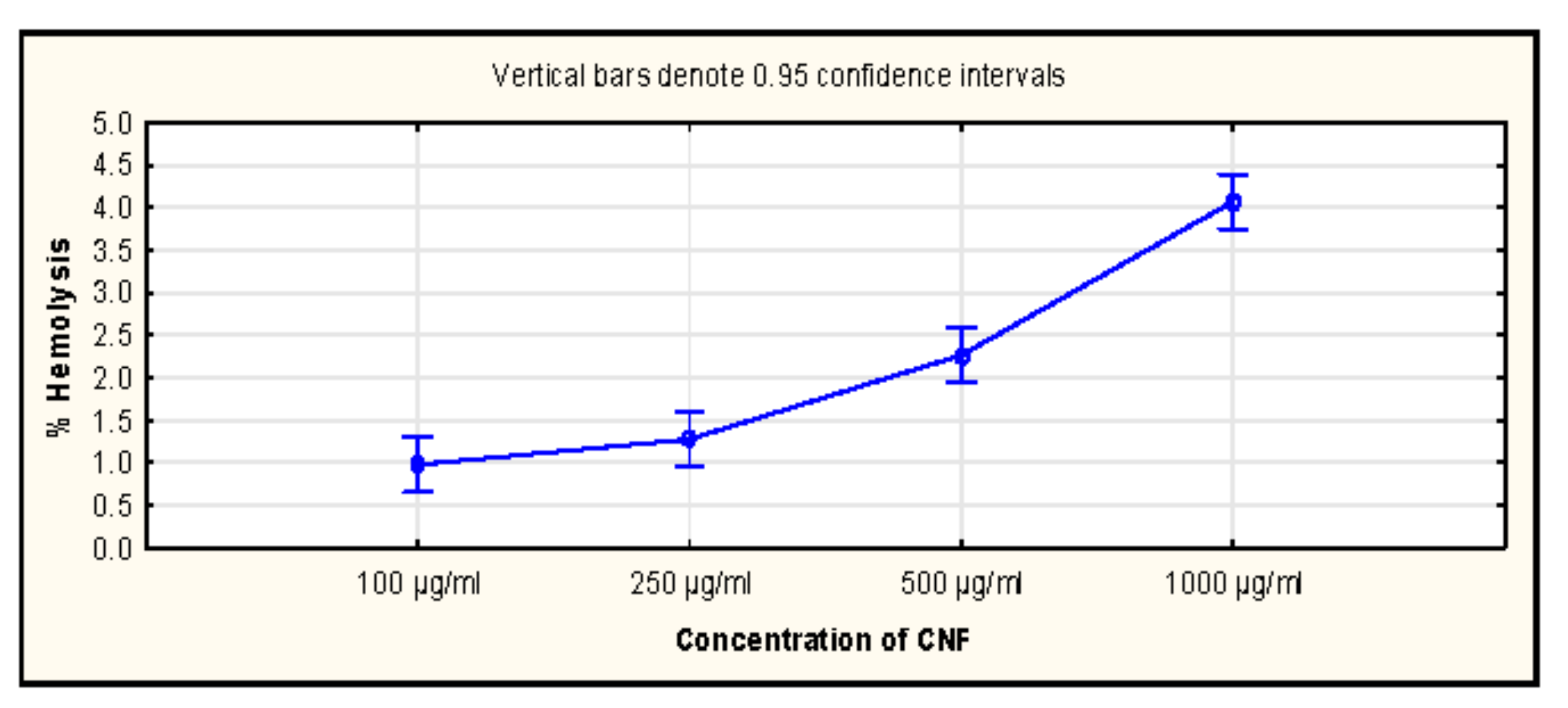
2.14. Haemocompatibility Assay
2.15. Statistical Analysis
3. Results
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mishra, P.K.; Mishra, H.; Ekielski, A.; Talegaonkar, S.; Vaidya, B. Zinc oxide nanoparticles: A promising nanomaterial for biomedical applications. Drug Discov. Today 2017, 22, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Ekielski, A. The self-assembly of lignin and its application in nanoparticle synthesis: A short review. Nanomaterials 2019, 9, 243. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Mishra, P.K.; Talegaonkar, S.; Vaidya, B. Metal nanoparticles: A theranostic nanotool against cancer. Drug Discov. Today 2015, 20, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.C.; Harshita; Mishra, P.K.; Talegaonkar, S. Ceramic nanoparticles: Fabrication methods and applications in drug delivery. Curr. Pharm. Des. 2015, 21, 6165–6188. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Ekielski, A. A simple method to synthesize lignin nanoparticles. Colloids Interfaces 2019, 3, 52. [Google Scholar] [CrossRef]
- Klemm, D.; Heublein, B.; Fink, H.-P.; Bohn, A. Cellulose: Fascinating biopolymer and sustainable raw material. Angew. Chem. Int. Ed. 2005, 44, 3358–3393. [Google Scholar] [CrossRef] [PubMed]
- Klemm, D.; Kramer, F.; Moritz, S.; Lindström, T.; Ankerfors, M.; Gray, D.; Dorris, A. Nanocelluloses: A new family of nature-based materials. Angew. Chem. Int. Ed. 2011, 50, 5438–5466. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.K.; Gregor, T.; Wimmer, R. Utilising brewer’s spent grain as a source of cellulose nanofibres following separation of protein-based biomass. BioResources 2017, 12, 107–116. [Google Scholar] [CrossRef]
- Azizi Samir, M.A.S.; Alloin, F.; Dufresne, A. Review of recent research into cellulosic whiskers, their properties and their application in nanocomposite field. Biomacromolecules 2005, 6, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Klemm, D.; Schumann, D.; Kramer, F.; Heßler, N.; Koth, D.; Sultanova, B. Nanocellulose materials—Different cellulose, different functionality. Macromol. Symp. 2009, 280, 60–71. [Google Scholar] [CrossRef]
- Crowley, M.M.; Schroeder, B.; Fredersdorf, A.; Obara, S.; Talarico, M.; Kucera, S.; McGinity, J.W. Physicochemical properties and mechanism of drug release from ethyl cellulose matrix tablets prepared by direct compression and hot-melt extrusion. Int. J. Pharm. 2004, 269, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Entwistle, C.; Rowe, R. Plasticization of cellulose ethers used in the film coating of tablets. J. Pharm. Pharmacol. 1979, 31, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Grosswald, R.R.; Anderson, J.B.; Andrew, C.S. Method for the Manufacture of Pharmaceutical Cellulose Capsules. European Patent EP0587773A1, 30 July 1997. [Google Scholar]
- Vueba, M.; De Carvalho, L.B.; Veiga, F.; Sousa, J.; Pina, M. Influence of cellulose ether polymers on ketoprofen release from hydrophilic matrix tablets. Eur. J. Pharm. Biopharm. 2004, 58, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, N.; Dufresne, A. Nanocellulose in biomedicine: Current status and future prospect. Eur. Polym. J. 2014, 59, 302–325. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Yano, H. Formation of hydrogels from cellulose nanofibers. Carbohydr. Polym. 2011, 85, 733–737. [Google Scholar] [CrossRef]
- Fischer, F.; Rigacci, A.; Pirard, R.; Berthon-Fabry, S.; Achard, P. Cellulose-based aerogels. Polymer 2006, 47, 7636–7645. [Google Scholar] [CrossRef]
- Anderson, R.L.; Owens, J.W.; Timms, C.W. The toxicity of purified cellulose in studies with laboratory animals. Cancer Lett. 1992, 63, 83–92. [Google Scholar] [CrossRef]
- Cullen, R.T.; Miller, B.G.; Jones, A.D.; Davis, J.M.G. Toxicity of cellulose fibres. Ann. Occup. Hyg. 2002, 46, 81–84. [Google Scholar]
- Ilves, M.; Vilske, S.; Aimonen, K.; Lindberg, H.K.; Pesonen, S.; Wedin, I.; Nuopponen, M.; Vanhala, E.; Højgaard, C.; Winther, J.R.; et al. Nanofibrillated cellulose causes acute pulmonary inflammation that subsides within a month. Nanotoxicology 2018, 12, 729–746. [Google Scholar] [CrossRef]
- Catalán, J.; Rydman, E.; Aimonen, K.; Hannukainen, K.-S.; Suhonen, S.; Vanhala, E.; Moreno, C.; Meyer, V.; da Silva Perez, D.; Sneck, A.; et al. Genotoxic and inflammatory effects of nanofibrillated cellulose in murine lungs. Mutagenesis 2016, 32, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Ogonowski, M.; Edlund, U.; Gorokhova, E.; Linde, M.; Ek, K.; Liewenborg, B.; Könnecke, O.; Navarro, J.R.G.; Breitholtz, M. Multi-level toxicity assessment of engineered cellulose nanofibrils in Daphnia magna. Nanotoxicology 2018, 12, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, J.; Mishra, H.; Mishra, P.K.; Wimmer, R.W.; Ahmad, F.J.; Talegaonkar, S. Cellulose Nanofiber Aerogel as a Promising Biomaterial for Customized Oral Drug Delivery. Available online: https://www.dovepress.com/cellulose-nanofiber-aerogel-as-a-promising-biomaterial-for-customized--peer-reviewed-article-IJN (accessed on 27 March 2017).
- Pandey, U.B.; Nichols, C.D. Human Disease Models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Yoshida, H. Drosophila as a model organism. Adv. Exp. Med. Biol. 2018, 1076, 1–10. [Google Scholar] [PubMed]
- Panacek, A.; Prucek, R.; Safarova, D.; Dittrich, M.; Richtrova, J.; Benickova, K.; Zboril, R.; Kvitek, L. Acute and chronic toxicity effects of silver nanoparticles (NPs) on Drosophila melanogaster. Environ. Sci. Technol. 2011, 45, 4974–4979. [Google Scholar] [CrossRef] [PubMed]
- Carmona, E.R.; Escobar, B.; Vales, G.; Marcos, R. Genotoxic testing of titanium dioxide anatase nanoparticles using the wing-spot test and the comet assay in Drosophila. Mutat. Res. Toxicol. Environ. Mutagen. 2015, 778, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Krebs, R.A.; Roberts, S.P.; Bettencourt, B.R.; Feder, M.E. Changes in thermotolerance and Hsp70 expression with domestication in Drosophila melanogaster. J. Evol. Biol. 2001, 14, 75–82. [Google Scholar] [CrossRef]
- Jakubowski, B.R.; Longoria, R.A.; Shubeita, G.T. A high throughput and sensitive method correlates neuronal disorder genotypes to Drosophila larvae crawling phenotypes. Fly (Austin) 2012, 6, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Nichols, C.D.; Becnel, J.; Pandey, U.B. Methods to assay Drosophila behavior. JoVE J. Vis. Exp. 2012, e3795. [Google Scholar] [CrossRef]
- Mishra, M.; Barik, B.K. Behavioral teratogenesis in Drosophila melanogaster. Methods Mol. Biol. 2018, 1797, 277–298. [Google Scholar]
- Sabat, D.; Patnaik, A.; Ekka, B.; Dash, P.; Mishra, M. Investigation of titania nanoparticles on behaviour and mechanosensory organ of Drosophila melanogaster. Physiol. Behav. 2016, 167, 76–85. [Google Scholar] [CrossRef]
- Priyadarsini, S.; Sahoo, S.K.; Sahu, S.; Mukherjee, S.; Hota, G.; Mishra, M. Oral administration of graphene oxide nano-sheets induces oxidative stress, genotoxicity, and behavioral teratogenicity in Drosophila melanogaster. Environ. Sci. Pollut. Res. 2019, 26, 19560–19574. [Google Scholar] [CrossRef] [PubMed]
- Pappus, S.A.; Ekka, B.; Sahu, S.; Sabat, D.; Dash, P.; Mishra, M. A toxicity assessment of hydroxyapatite nanoparticles on development and behaviour of Drosophila melanogaster. J. Nanopart. Res. 2017, 19, 136. [Google Scholar] [CrossRef]
- Anwar, M.; Akhter, S.; Mallick, N.; Mohapatra, S.; Zafar, S.; Rizvi, M.M.A.; Ali, A.; Ahmad, F.J. Enhanced anti-tumor efficacy of paclitaxel with PEGylated lipidic nanocapsules in presence of curcumin and poloxamer: In vitro and in vivo studies. Pharmacol. Res. 2016, 113, 146–165. [Google Scholar] [CrossRef] [PubMed]
- Pappus, S.A.; Mishra, M. A Drosophila model to decipher the toxicity of nanoparticles taken through oral routes. Adv. Exp. Med. Biol. 2018, 1048, 311–322. [Google Scholar] [PubMed]
- Kitamoto, T. Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitiveshibire allele in defined neurons. J. Neurobiol. 2001, 47, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Pulver, S.R.; Pashkovski, S.L.; Hornstein, N.J.; Garrity, P.A.; Griffith, L.C. Temporal dynamics of neuronal activation by channelrhodopsin-2 and TRPA1 determine behavioral output in Drosophila larvae. J. Neurophysiol. 2009, 101, 3075–3088. [Google Scholar] [CrossRef]
- Ariano, P.; Zamburlin, P.; Gilardino, A.; Mortera, R.; Onida, B.; Tomatis, M.; Ghiazza, M.; Fubini, B.; Lovisolo, D. Interaction of spherical silica nanoparticles with neuronal cells: Size-dependent toxicity and perturbation of calcium homeostasis. Small 2011, 7, 766–774. [Google Scholar] [CrossRef]
- Barik, B.K.; Mishra, M. Nanoparticles as a potential teratogen: A lesson learnt from fruit fly. Nanotoxicology 2018, 13, 258–284. [Google Scholar] [CrossRef]
- Posgai, R.; Cipolla-McCulloch, C.B.; Murphy, K.R.; Hussain, S.M.; Rowe, J.J.; Nielsen, M.G. Differential toxicity of silver and titanium dioxide nanoparticles on Drosophila melanogaster development, reproductive effort, and viability: Size, coatings and antioxidants matter. Chemosphere 2011, 85, 34–42. [Google Scholar] [CrossRef]
- Montell, C. Drosophila TRP channels. Pflügers Arch. Eur. J. Physiol. 2005, 451, 19–28. [Google Scholar] [CrossRef]
- Neely, G.G.; Keene, A.C.; Duchek, P.; Chang, E.C.; Wang, Q.-P.; Aksoy, Y.A.; Rosenzweig, M.; Costigan, M.; Woolf, C.J.; Garrity, P.A.; et al. TrpA1 regulates thermal nociception in Drosophila. PLoS ONE 2011, 6, e24343. [Google Scholar] [CrossRef] [PubMed]
- Tracey, W.D.; Wilson, R.I.; Laurent, G.; Benzer, S. Painless, a Drosophila gene essential for nociception. Cell 2003, 113, 261–273. [Google Scholar] [CrossRef]
- Fowler, M.A.; Montell, C. Drosophila TRP channels and animal behavior. Life Sci. 2013, 92, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Afonso, B.; Vonner, A.J.; Hernandez-Nunez, L.; Berck, M.; Tabone, C.J.; Kane, E.A.; Pieribone, V.A.; Nitabach, M.N.; Cardona, A.; et al. Sensory determinants of behavioral dynamics in Drosophila thermotaxis. Proc. Natl. Acad. Sci. USA 2014, 112, E220–E229. [Google Scholar] [CrossRef] [PubMed]
- Mohammadpour, R.; Yazdimamaghani, M.; Reilly, C.; Ghandehari, H. Transient receptor potential (TRP) ion channel—Dependent toxicity of silica nanoparticles and poly(amido amine) (PAMAM) dendrimers. J. Pharmacol. Exp. Ther. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Liu, J.; Guo, C.; Zhou, Y.; Teng, Y.; Liu, L. Two pairs of neurons in the central brain control Drosophila innate light preference. Science 2010, 330, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Keene, A.C.; Mazzoni, E.O.; Zhen, J.; Younger, M.A.; Yamaguchi, S.; Blau, J.; Desplan, C.; Sprecher, S.G. Distinct visual pathways mediate Drosophila larval light avoidance and circadian clock entrainment. J. Neurosci. 2011, 31, 6527–6534. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, N.; Romero, N.M.; Martin, F.A.; Rewitz, K.F.; Sun, M.; O’Connor, M.B.; Leopold, P. Neuroendocrine control of Drosophila larval light preference. Science 2013, 341, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Key, S.C.S.; Reaves, D.; Turner, F.; Bang, J.J. Impacts of silver nanoparticle ingestion on pigmentation and developmental progression in Drosophila. Atlas J. Biol. 2011, 1, 52–61. [Google Scholar]
- Raj, A.; Shah, P.; Agrawal, N. Ingestion of gold nanoparticles (AuNPs) affects survival in Drosophila in a dose-dependent manner. Int. J. Sci. Res. 2016, 5. [Google Scholar] [CrossRef]
- Berthold, H.K.; Unverdorben, S.; Degenhardt, R.; Unverdorben, M.; Gouni-Berthold, I. Effect of a cellulose-containing weight-loss supplement on gastric emptying and sensory functions. Obesity 2008, 16, 2272–2280. [Google Scholar] [CrossRef] [PubMed]
- Papathanasopoulos, A.; Camilleri, M. Dietary fiber supplements: Effects in obesity and metabolic syndrome and relationship to gastrointestinal functions. Gastroenterology 2010, 138, 65–72. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, P.K.; Ekielski, A.; Mukherjee, S.; Sahu, S.; Chowdhury, S.; Mishra, M.; Talegaonkar, S.; Siddiqui, L.; Mishra, H. Wood-Based Cellulose Nanofibrils: Haemocompatibility and Impact on the Development and Behaviour of Drosophila melanogaster. Biomolecules 2019, 9, 363. https://doi.org/10.3390/biom9080363
Mishra PK, Ekielski A, Mukherjee S, Sahu S, Chowdhury S, Mishra M, Talegaonkar S, Siddiqui L, Mishra H. Wood-Based Cellulose Nanofibrils: Haemocompatibility and Impact on the Development and Behaviour of Drosophila melanogaster. Biomolecules. 2019; 9(8):363. https://doi.org/10.3390/biom9080363
Chicago/Turabian StyleMishra, Pawan Kumar, Adam Ekielski, Sumit Mukherjee, Swetapadma Sahu, Saptarshi Chowdhury, Monalisa Mishra, Sushama Talegaonkar, Lubna Siddiqui, and Harshita Mishra. 2019. "Wood-Based Cellulose Nanofibrils: Haemocompatibility and Impact on the Development and Behaviour of Drosophila melanogaster" Biomolecules 9, no. 8: 363. https://doi.org/10.3390/biom9080363