Antimicrobial Efficacy of Five Probiotic Strains Against Helicobacter pylori

 , , , , and
, , , , and
Abstract
:1. Introduction
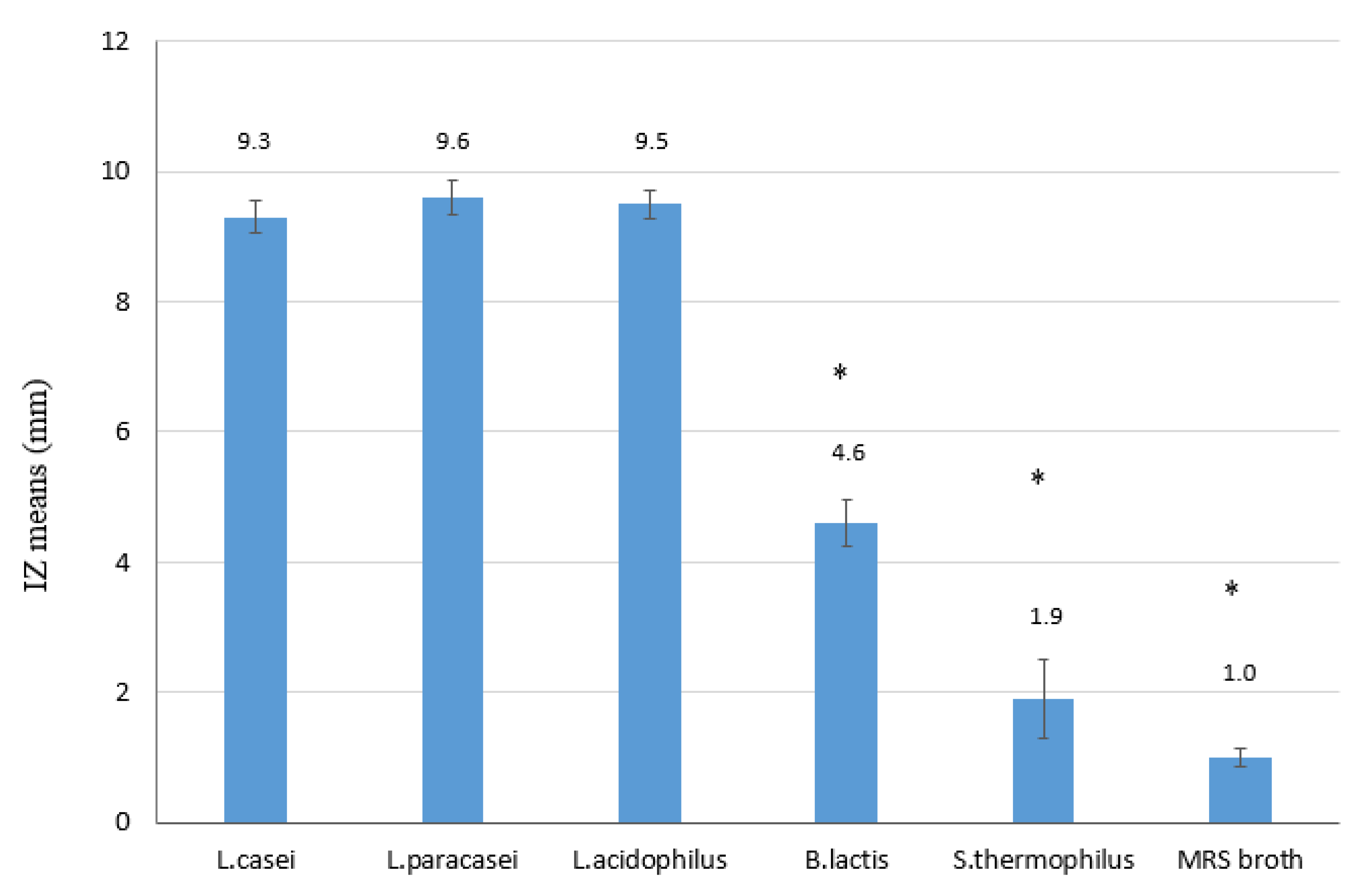
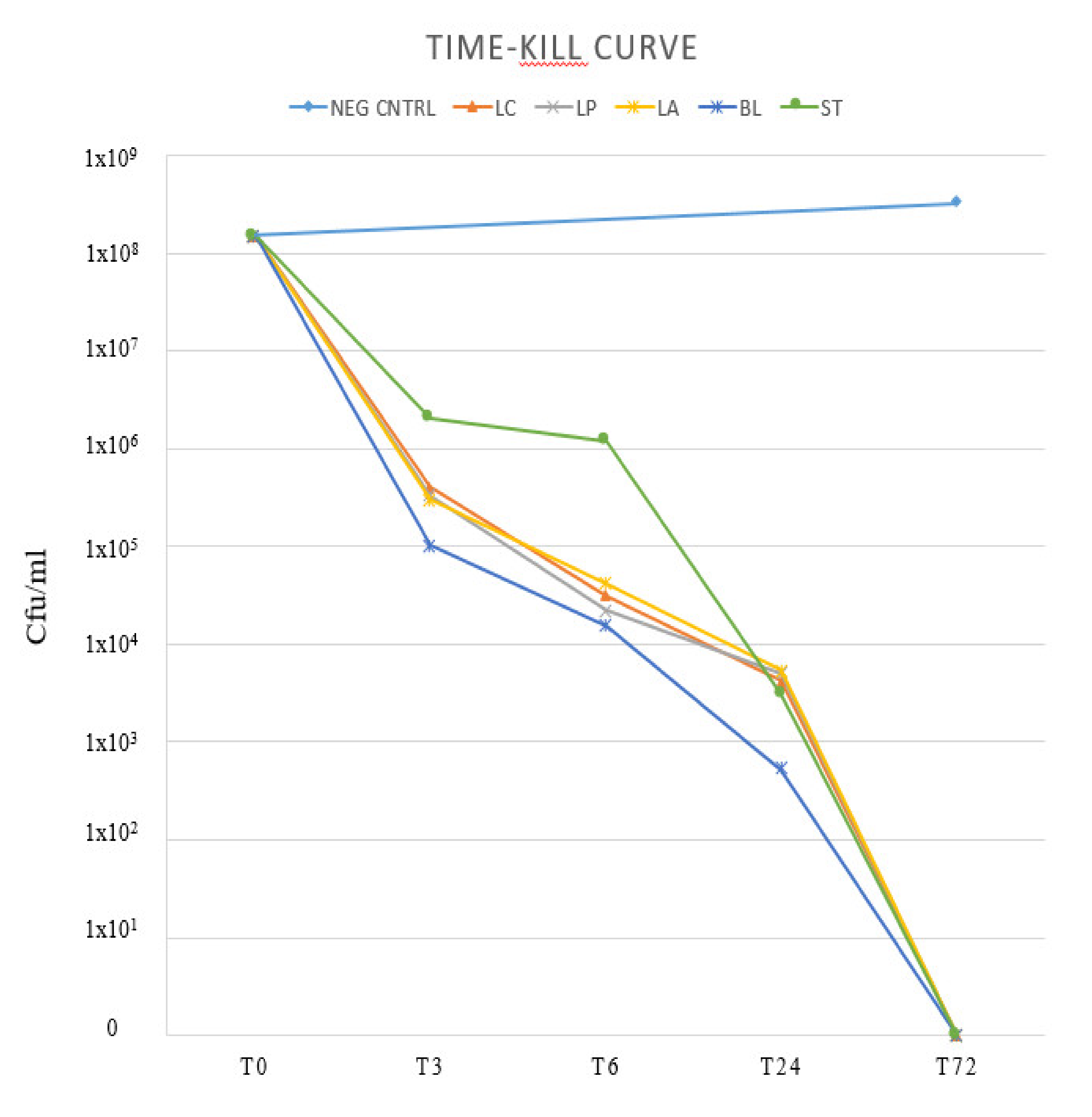
2. Results
3. Discussion
4. Materials and Methods
4.1. Helicobacter Pylori Culture and Susceptibility Test
4.2. Probiotic Strains
4.3. Overnight Broth Cultures
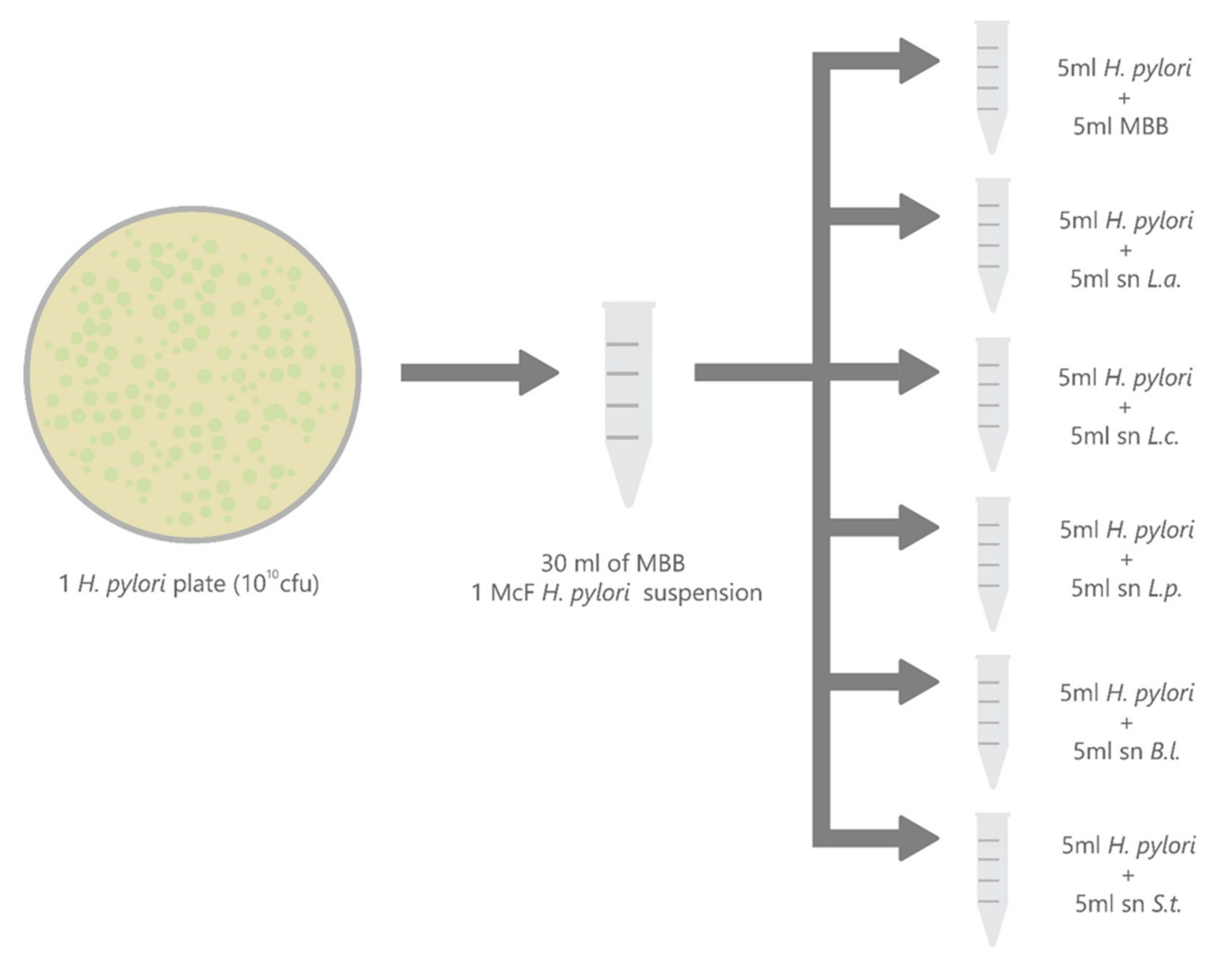
4.4. Supernatants
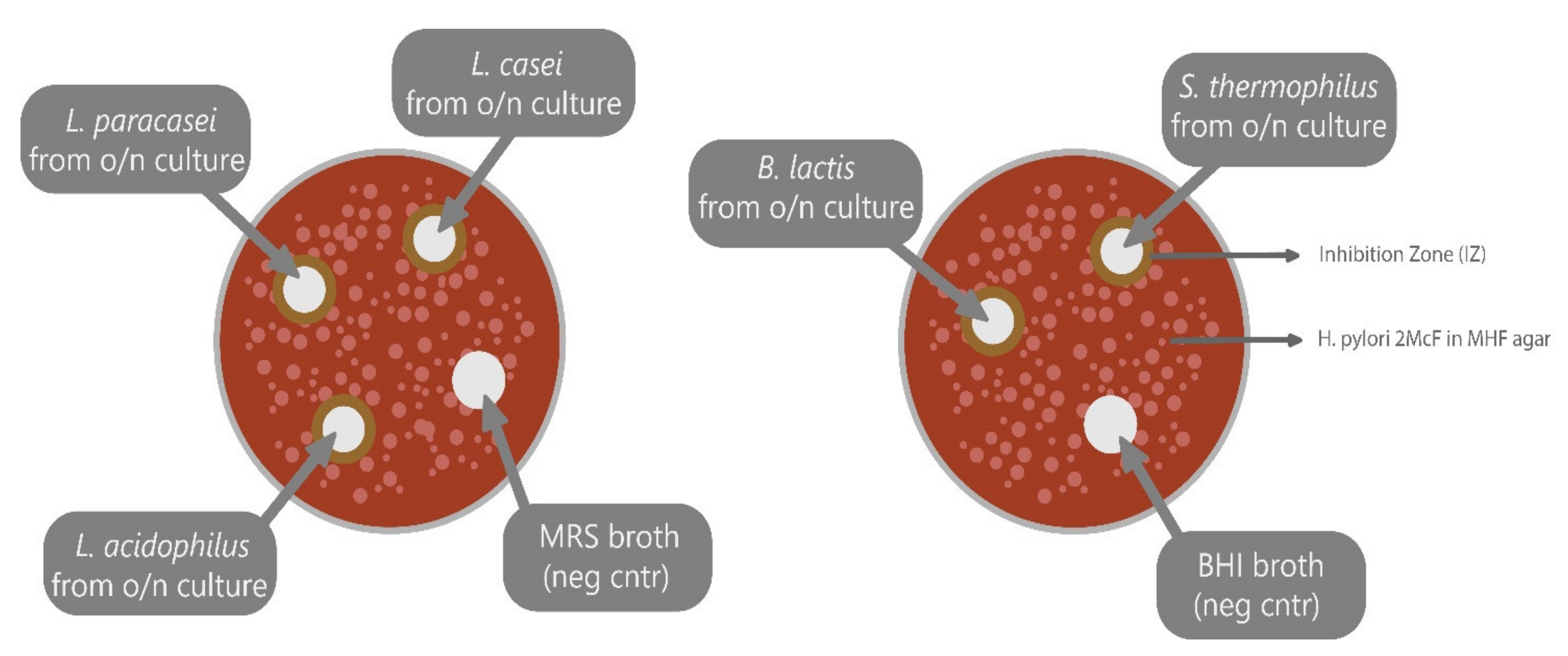
4.5. Agar Well Diffusion (Bacteriostatic Activity).
4.6. Time-Kill Curve (Bactericidal Activity)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holton, J.; Figura, N.; Vaira, D. Helicobacter pylori: Atlas of Investigation and Management; Clinical Publishing: Oxford, UK, 2012. [Google Scholar]
- Saracino, I.M.; Fiorini, G.; Zullo, A.; Pavoni, M.; Saccomanno, L.; Vaira, D. Trends in primary antibiotic resistance in H.pylori strains isolated in Italy between 2009 and 2019. Antibiotics 2020, 9, 26. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, P. Use of probiotics in the fight against Helicobacter pylori. World J. Gastrointest. Pathophys. 2014, 5, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Shah, N.; Prajapati, J.B. Clinical application of probiotics in the treatment of Helicobacter pylori infection. A brief review. J. Microbiol. Immunol. Infect. 2014, 47, 429–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bathia, S.J.; Kochar, N.; Abraham, P.; Nair, N.G.; Metha, A.P. Lactobacillus acidophilus Inhibits Growth of Campylobacter pylori In Vitro. J. Clin. Microbiol. 1989, 27, 2328–2330. [Google Scholar]
- Shi, X.; Zhang, J.; Mo, L.; Shi, J.; Qin, M.; Huang, X. Efficacy and safety of probiotics in eradicating Helicobacter pylori A network meta-analysis. Medicine 2019, 98, 7. [Google Scholar]
- Fuhler, G.M.; Hecht, G.A.; Foley, S.E.; Lebron, A.C.; Hazleton, K. 2019 Gut Microbiota for Health World Summit. Gastroenterology 2020, 158, 28–31. [Google Scholar] [CrossRef] [Green Version]
- Nardone, G.; Compare, D. The human gastric microbiota: Is it time to rethink the pathogenesis of stomach diseases? UEG J. 2015, 3, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Brawner, K.M.; Morrow, C.D.; Smith, P.D. Gastric Microbiome and Gastric Cancer. Cancer J. 2014, 3, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Yu, J. Effect of Helicobacter pylori Infection on the Composition of Gastric Microbiota in the Development of Gastric Cancer. Gastrointest. Tumors 2015, 2, 14–25. [Google Scholar] [CrossRef]
- Perdigon, G.; Medina, M.; Vintiñi, E.; Valdéz, J.C. Intestinal pathway of internalization of lactic acid bacteria and gut mucosal immunostimulation. Int. J. Immunopathol. Pharmacol. 2000, 13, 141–150. [Google Scholar]
- Yang, Y.J.; Chuang, C.C.; Yang, H.B.; Lu, C.C.; Sheu, B.S. Lactobacillus acidophilus ameliorates, H. pylori induced gastric inflammation by inactivating the Smad7 and NFkB pathways. BMC Microbiol. 2012, 12, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murosaki, S.; Muroyama, K.; Yamamoto, Y.; Yoshikai, Y. Antitumor effect of heat-killed Lactobacillus plantarum L-137 through restoration of impaired interleukin-12 production in tumorbearing mice. Cancer Immunol. Immunother. 2000, 49, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Kabir, A.M.; Aiba, Y.; Takagi, A.; Kamiya, S.; Miwa, T.; Koga, Y. Prevention of Helicobacter pylori infection by lactobacilli in a gnotobiotic murine model. Gut 1997, 41, 49–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal epithelial cells in vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Mattar, A.F.; Teitelbaum, D.H.; Drongowski, R.A.; Yongyi, F.; Harmon, C.M.; Coran, A.G. Probiotics up-regulate MUC-2 mucin gene expression in a Caco-2 cell-culture model. Pediatr. Surg. Int. 2002, 18, 586–590. [Google Scholar] [PubMed]
- Sanz, Y. Antimicrobial peptides are among the antagonistic metabolites produced by Bifidobacterium against Helicobacter pylori. Int. J. Antimicrob. Agents 2005, 25, 385–389. [Google Scholar]
- Saracino, I.M.; Pavoni, M.; Zullo, A.; Fiorini, G.; Saccomanno, L.; Lazzarotto, T.; Cavallo, R.; Antonelli, G.; Vaira, D. Antibiotic resistance and therapy outcome in H.pylori eradication failure patients. Antiobiotics 2020, 9, 121. [Google Scholar]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Atherton, J.; Axon, A.T.; Bazzoli, F.; El-Omar, E.M. Management of Helicobacter pylori Infection—The Maastricht IV/Florence Consensus Report. Gut 2012, 61, 646–664. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-H.; Gao, Q.-Y.; Fang, J.-Y. Meta-Analysis of the Efficacy and Safety of Lactobacillus-containing and Bifidobacterium-containing Probiotic Compound Preparation in Helicobacter pylori Eradication Therapy. J. Clin. Gastroenterol. 2013, 47, 25–32. [Google Scholar] [CrossRef]
- Tomás, N.-R.; Fernando, M.S.; Ricardo, C.B.; Rejane, M.J.; Prado, M.-F.; de Oliveira, M.N.; Bogsan, C.S.; Chinzon, D.; Eisig, J.N. Association of a probiotic to a Helicobacter pylori eradication regimen does not increase efficacy or decreases the adverse effects of the treatment: A prospective, randomized, double-blind, placebo-controlled study. BMC Gastroenterol. 2013, 13, 56. [Google Scholar]
- Lau, C.S.M.; Ward, A.; Chamberlain, R.S. Probiotics improve the efficacy of standard triple therapy in the eradication of Helicobacter pylori: A meta-analysis. Infect. Drug Resist. 2016, 9, 275–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakya, S.; Zhang, G.Q.; Hu, H.J.; Li, Z.Y. Lactobacillus GG Supplementation on Anti- Helicobacter pylori Therapy-Related Side Effects and Eradication Rates: A Meta-Analysis. Open J. Internal Med. 2015, 5, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Enany, S.; Abdalla, S. In vitro antagonistic activity of Lactobacillus casei against Helicobacter pylori. Brazil. J. Microbiol. 2015, 46, 1201–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyanova, L.; Stephanova-Kondratenko, M.; Mitov, I. Anti-Helicobacter pylori activity of Lactobacillus delbrueckii subsp. bulgaricus strains: Preliminary report. Soc. Appl. Microbiol. Lett. Appl. Microbiol. 2009, 48, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Medouakh, A.; Abdeslam, A.; Belsotane, A. Antagonistic activity of Lactobacillus sp. against Helicobacter pylori. Int. J. Microbiol. Res. 2010, 1, 80–86. [Google Scholar]
- Sgouras, D.; Maragkoudakis, P.; Petraki, K.; Martinez-Gonzalez, B.; Eriotou, E.; Michopoulos, S.; Kalantzopoulos, G.; Tsakalidou, A.; Mentis, E. In Vitro and In Vivo Inhibition of Helicobacter pylori by Lactobacillus casei Strain Shirota. Appl. Environ. Microbiol. 2004, 70, 518–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cats, A.; Kuipers, E.J.; Bosschaert, M.A.; Pot, R.G.J.; Vandenbrouckergrauls, C.M.J.E.; Kusters, J.G. Effect of frequent consumption of a Lactobacillus casei-containing milk drink in Helicobacter pylori-colonized subjects. Aliment. Pharmacol. Ther. 2003, 17, 429–435. [Google Scholar] [CrossRef]
- Boke, H.; Aslim, B.; Alp, G. Rore of resistance to bile salts and acid tolerance of exopolysaccarides (EPSS) produced by yogurt starter bacteria. Arch. Biol. Sci. Belgrade 2014, 62, 323–328. [Google Scholar] [CrossRef]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbial. 2013, 4, 1–8. [Google Scholar]
- Fuochi, V.; Petronio, G.; Lissandrello, E.; Furneri, P.M. Evaluation of resistance to low pH and bile salts of human Lactobacillus spp. Isolates. Int. J. Immunopathol. Pharmacol. 2015, 28, 426–433. [Google Scholar] [CrossRef] [Green Version]
- De Francesco, V.; Zullo, A.; Hassan, C.; Giorgio, F.; Rosania, R.; Ierardi, E. Mechanisms of Helicobacter pylori antibiotic resistance: An updated appraisal. World J. Gastrointest. Pathophysiol. 2011, 2, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Clinical Breakpoints—Breakpoints and Guidance. Available online: http://www.eucast.org/clinical_breakpoints (accessed on 7 January 2020).
- Murray, P.R. What Is New in Clinical Microbiology—Microbial Identification by MALDI-TOF Mass Spectrometry. J. Mol. Diagn. 2012, 14, 419–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probiotic Strain | N° of Cells/mL (Range) | Means | SD | pH Range |
|---|---|---|---|---|
| L. casei | 9 × 109–1 × 1010 | 9.4 × 109 | 3.1 × 108 | 4 to 4.5 |
| L. paracasei | 6.3 × 109–1 × 1010 | 7.7 × 109 | 1 × 109 | 4 to 4.5 |
| L. acidophilus | 6.6 × 109–8 × 109 | 7.1 × 109 | 4.5 × 108 | 4 to 4.5 |
| B. lactis | 3.6 × 109–7.5 × 109 | 6.1 × 109 | 1.8 × 109 | 4 to 4.5 |
| S. thermophilus | 3 × 109–6.9 × 109 | 3.7 × 109 | 1.4 × 109 | 5 to 6 |
| Resistance Patterns | Number of Strains |
|---|---|
| ClaS, MzS, LS | 9 |
| ClaR, MzR, LR | 10 |
| ClaR, MzS, LS | 9 |
| ClaR, MzR, LS | 10 |
| ClaS, MzR, LS | 4 |
| ClaS, MzS, LR | 3 |
| ClaS, MzR, LR | 5 |
| ClaR, MzS, LR | 7 |
| Total | 57 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saracino, I.M.; Pavoni, M.; Saccomanno, L.; Fiorini, G.; Pesci, V.; Foschi, C.; Piccirilli, G.; Bernardini, G.; Holton, J.; Figura, N.; et al. Antimicrobial Efficacy of Five Probiotic Strains Against Helicobacter pylori. Antibiotics 2020, 9, 244. https://doi.org/10.3390/antibiotics9050244
Saracino IM, Pavoni M, Saccomanno L, Fiorini G, Pesci V, Foschi C, Piccirilli G, Bernardini G, Holton J, Figura N, et al. Antimicrobial Efficacy of Five Probiotic Strains Against Helicobacter pylori. Antibiotics. 2020; 9(5):244. https://doi.org/10.3390/antibiotics9050244
Chicago/Turabian StyleSaracino, Ilaria Maria, Matteo Pavoni, Laura Saccomanno, Giulia Fiorini, Valeria Pesci, Claudio Foschi, Giulia Piccirilli, Giulia Bernardini, John Holton, Natale Figura, and et al. 2020. "Antimicrobial Efficacy of Five Probiotic Strains Against Helicobacter pylori" Antibiotics 9, no. 5: 244. https://doi.org/10.3390/antibiotics9050244