Simulating Long-Term Development of Greenhouse Gas Emissions, Plant Biomass, and Soil Moisture of a Temperate Grassland Ecosystem under Elevated Atmospheric CO2

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Implementation
2.3. Model Setup
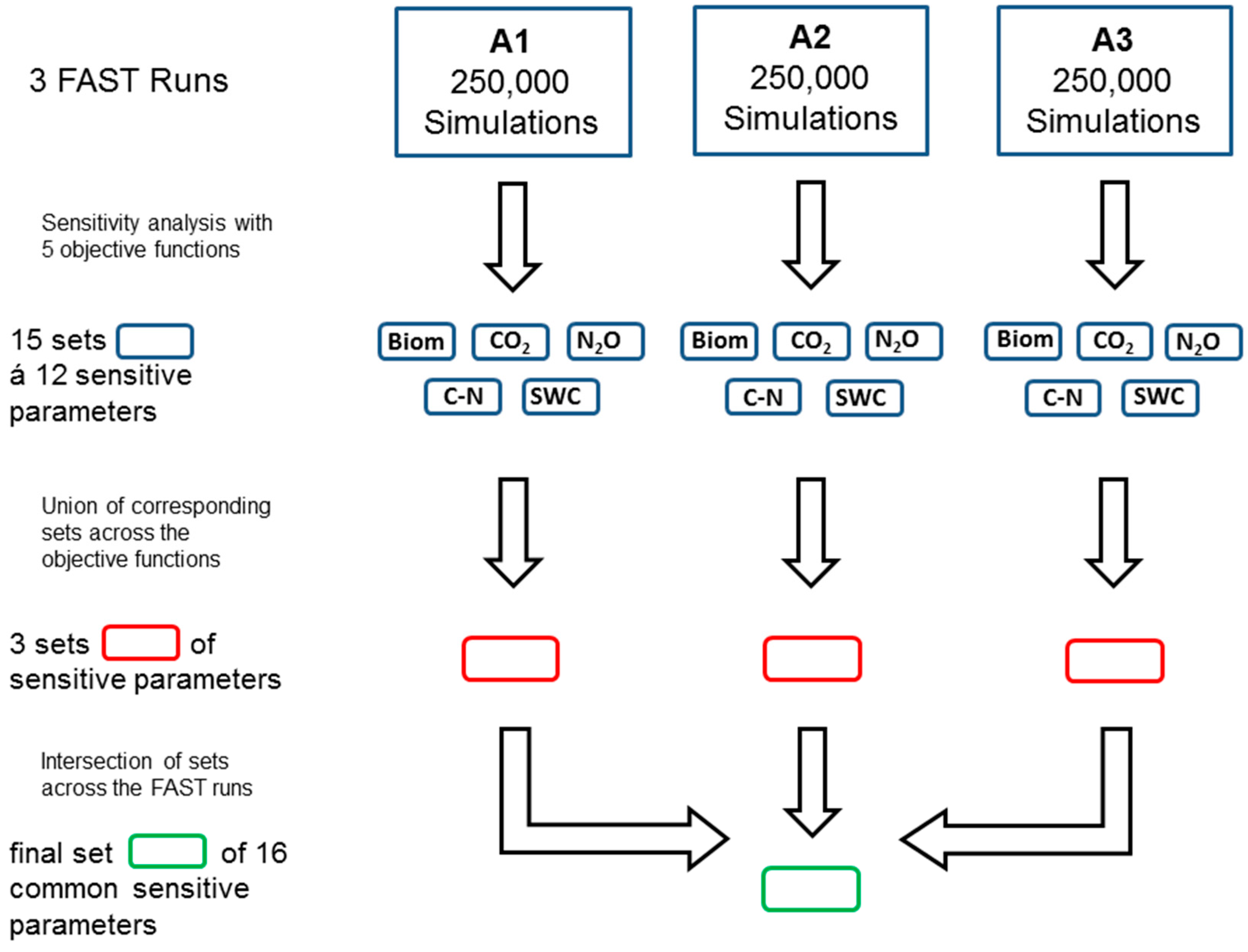
2.4. Sensitivity Analysis and Calibration
2.5. Evaluation
3. Results
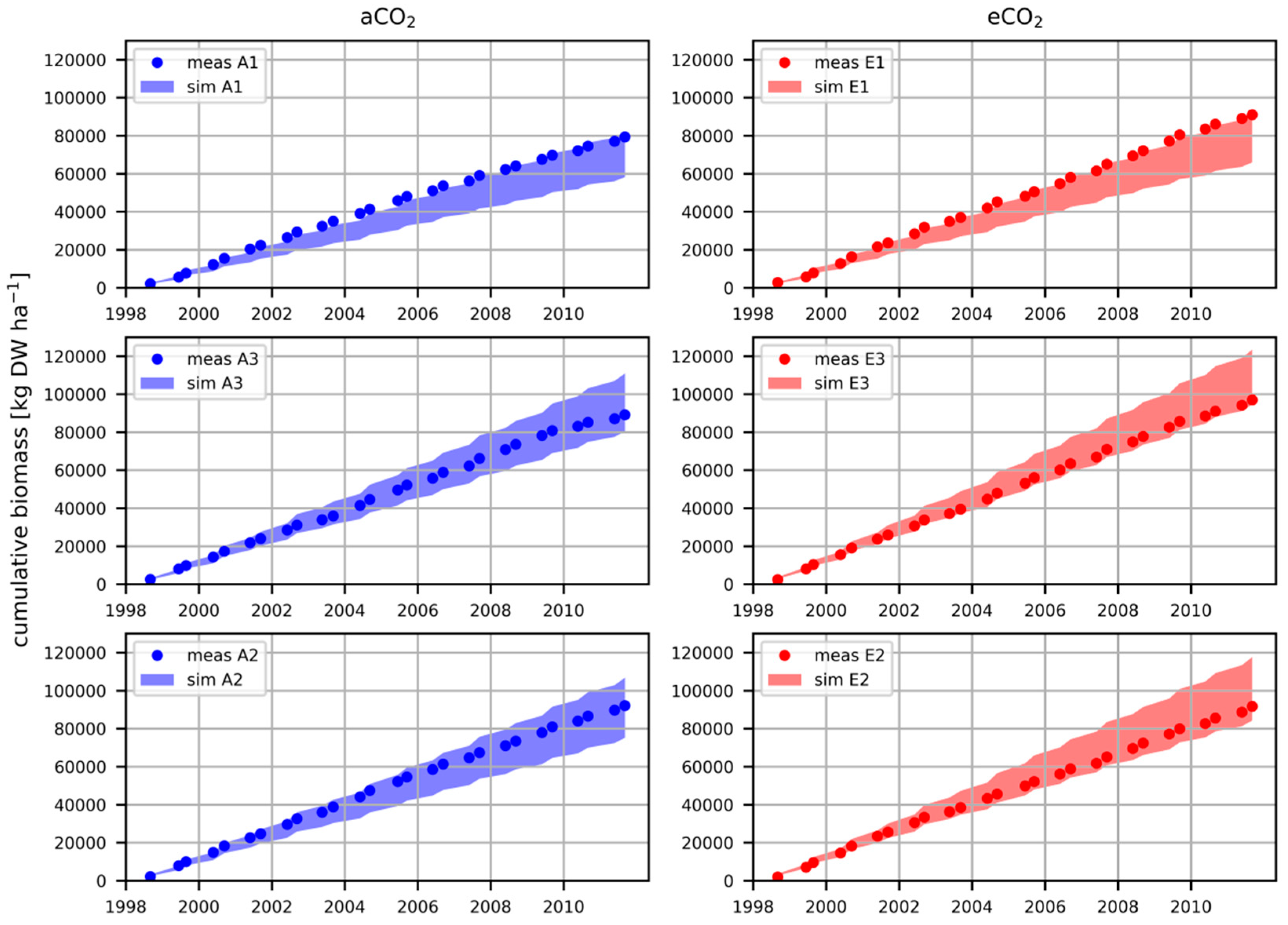
3.1. Cumulative Plant Biomass
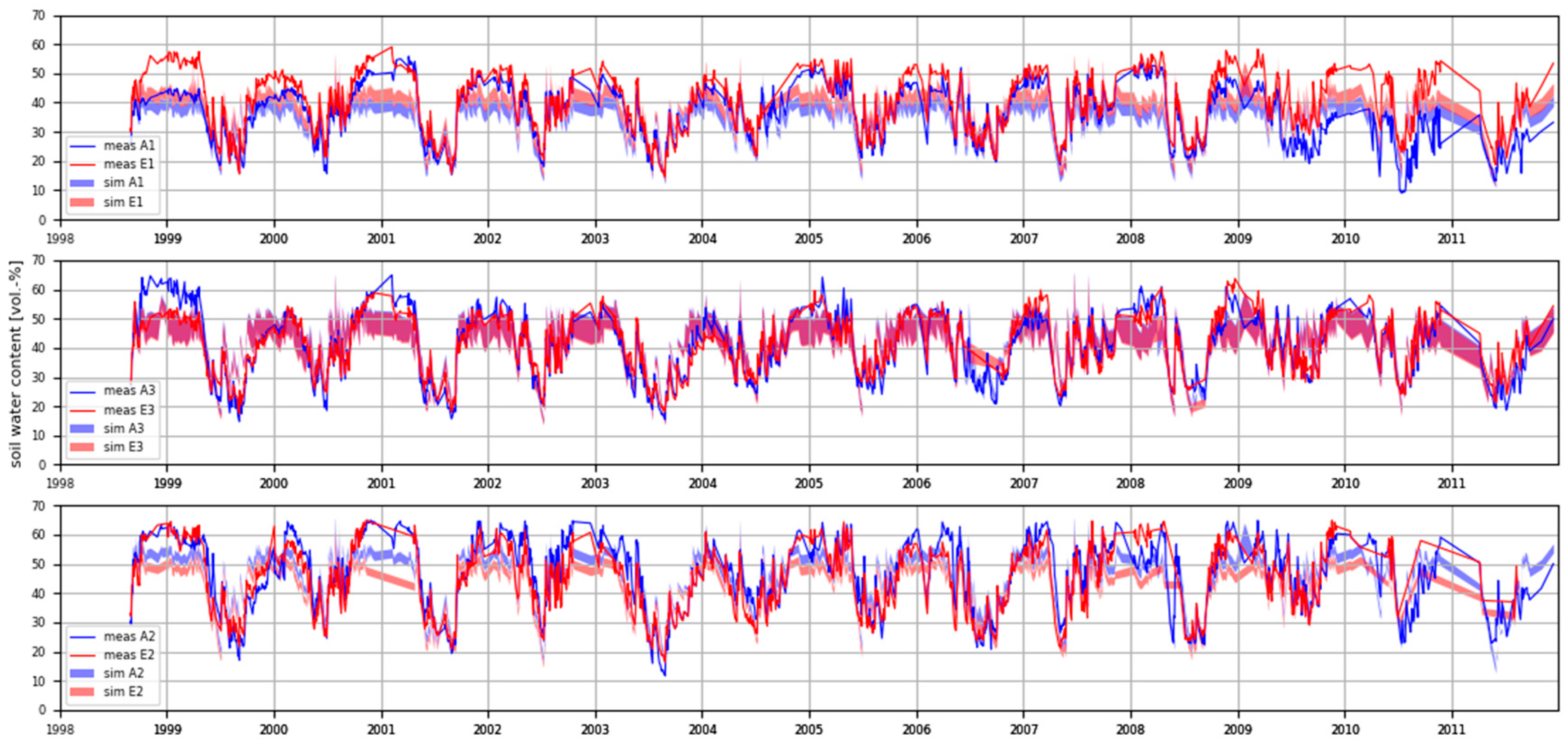
3.2. Soil Water Content
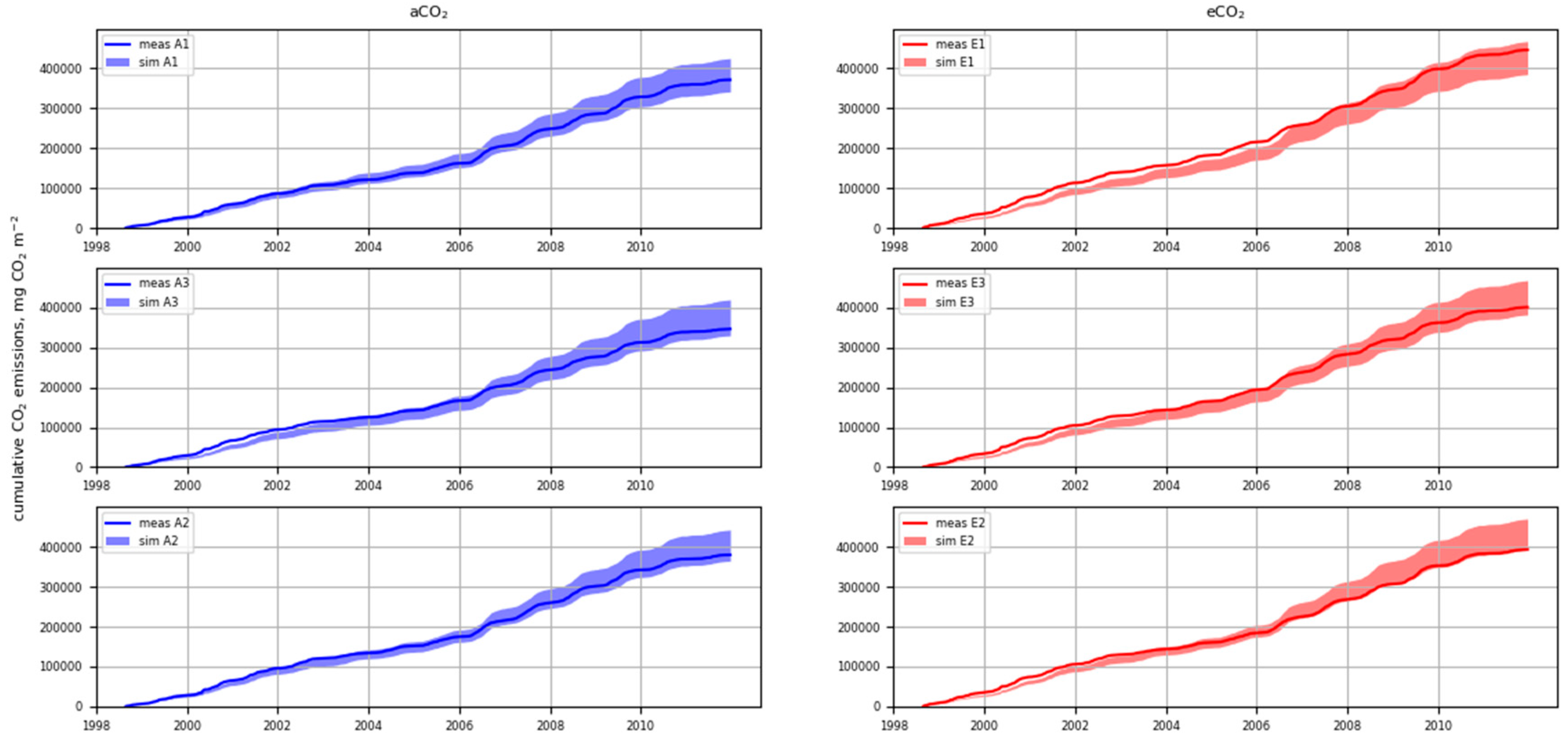
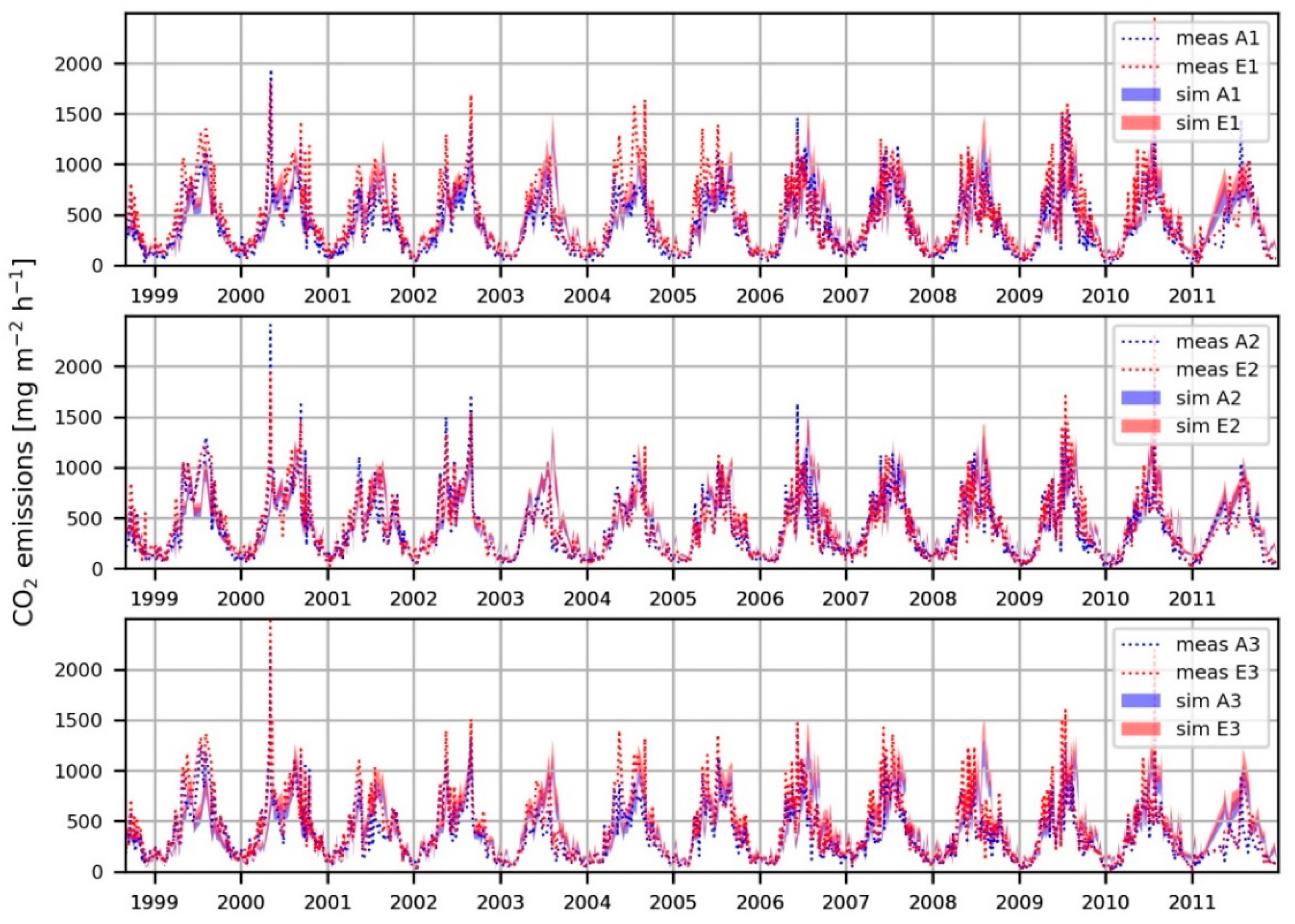
3.3. Cumulative CO2 Emissions
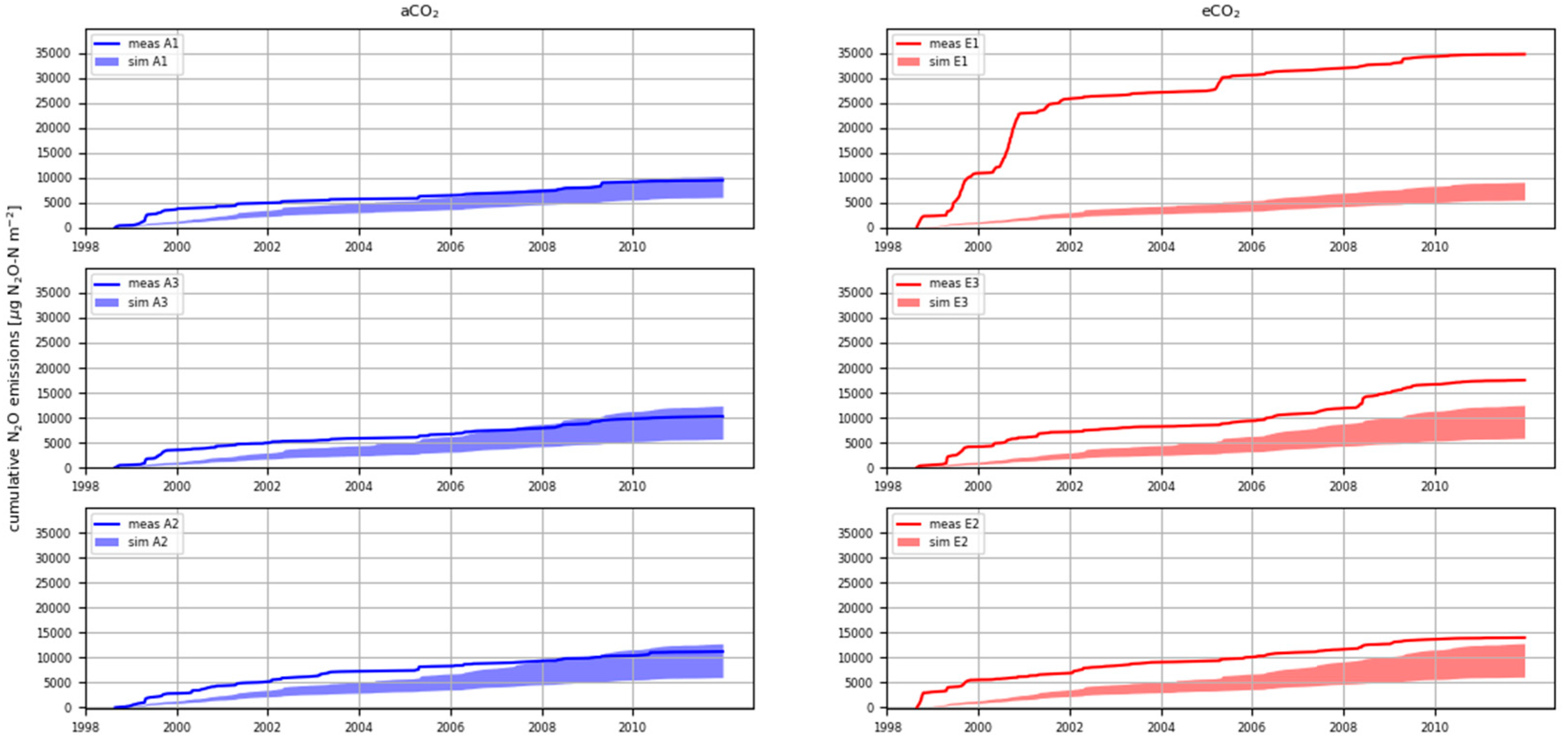
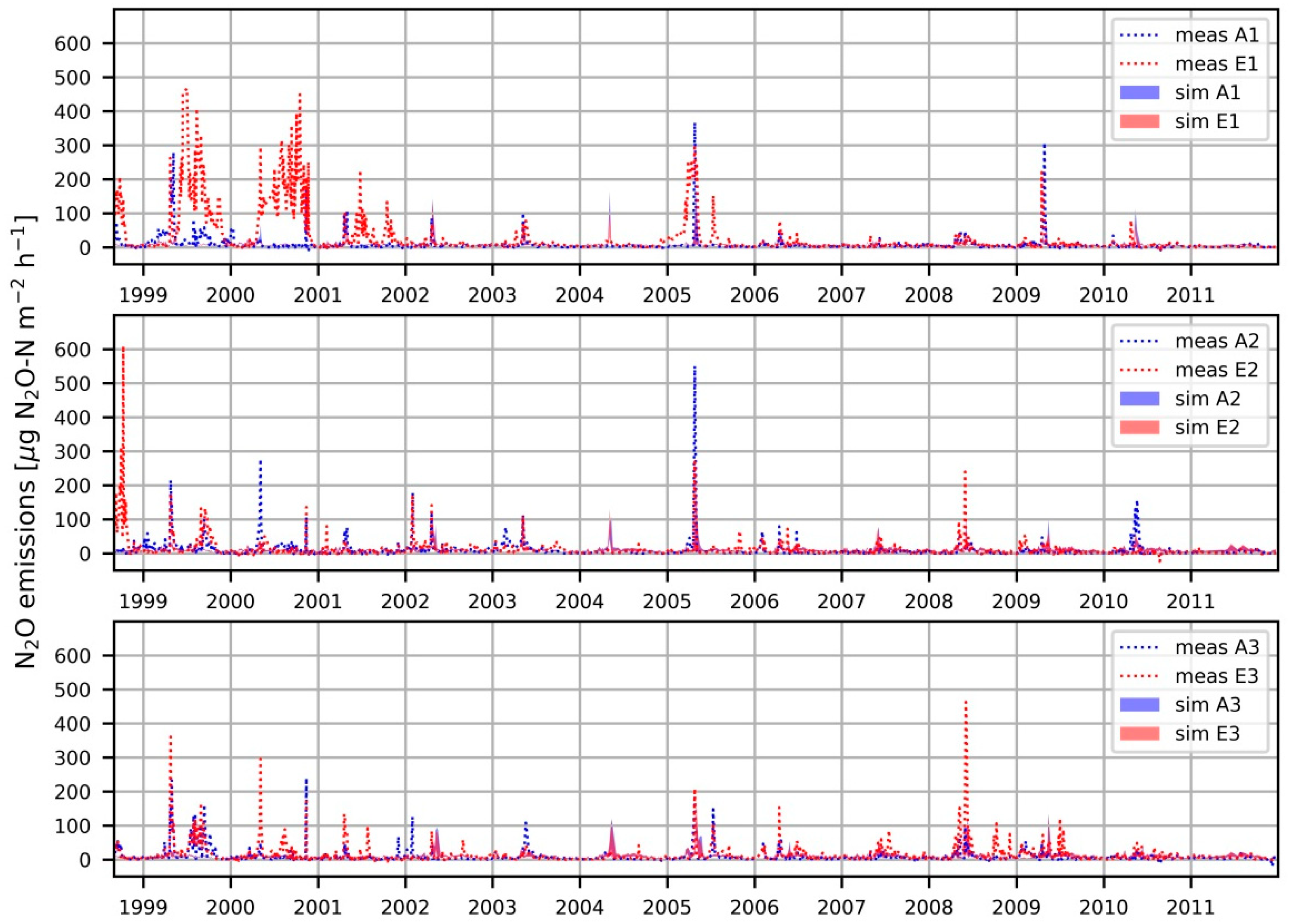
3.4. Cumulative N2O Emissions
4. Discussion
4.1. Biomass
4.2. SWC
4.3. Cumulative CO2 Emissions
4.4. Cumulative N2O Emissions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Value/Unit | Start/End | Temporal Resolution | Usage | Source |
|---|---|---|---|---|---|
| Air temperature (mean, min, max) | °C | 1995/2011 | daily | Driver data | WD |
| Global radiation | W m−2 | 1995/2011 | daily | Driver data | WD |
| Precipitation | mm day−1 | 1995/2011 | daily | Driver data | WD |
| Relative humidity | % | 1995/2011 | daily | Driver data | WD |
| Groundwater level | m | 1995/2011 | daily *1 | Driver data | FSM |
| Fertilizer application (ammonium nitrate) | 40 kg N ha−1 yr−1 | 1995/2011 | yearly | Driver data | [31] |
| N deposition | 14 kg N ha−1 yr−1 | 1993/1995 | mean | Driver data | [55] |
| Field capacity | mm m−1 | - | - | Calibrated parameter | [32] |
| Wilting point | mm m−1 | - | - | Calibrated parameter | [32] |
| Van Genuchten α | cm−1 | - | - | Calibrated parameter | [32] |
| Van Genuchten n | - | - | - | Calibrated parameter | [32] |
| Hydraulic Conductivity | cm min−1 | 2017 | - | Calibrated parameter | FSM |
| Fraction of soil org. N | 0.08–0.37% | 2001/2002 | - | Initialization | [29] |
| Fraction of soil org. C | 0.69–3.96% | 2001/2002 | - | Initialization | [29] |
| Soil pH | 5.4–6.0 | - | - | Fixed parameter | [29] |
| Cutting height | 4 cm | - | constant | Fixed parameter | FSM |
| Bulk density profile | 0.63–1.66 g cm−3 | - | - | Fixed parameter | [29,32] |
| Texture (clay, silt, sand) | - | - | constant | Fixed parameter | [32] |
| CO2 concentration | ppm | 1998/2011 | daily | Fixed parameter | FSM |
| Groundwater NO3− concentration | 0.05–23.32 mg L−1 | 2016 | daily | Fixed parameter | FSM |
| Plant C-N ratio | - | 1998/2011 | 2 cuts/year | Calibration data | FSM |
| Biomass | kg ha−1 yr−1 | 1998/2011 | 2 cuts/year | Calibration data | FSM |
| Soil water content | vol.-% | 1998/2011 | variable *2 | Calibration data | FSM |
| CO2 emissions | mg CO2 m−2 h−1 | 1998/2011 | variable | Calibration data | FSM |
| N2O emissions | µg N m−2 h−1 | 1998/2011 | variable | Calibration data | FSM |
| Module | Name | Int | Min | Max | Description |
|---|---|---|---|---|---|
| PLAMOX | AEJM | 46,270 | 37,000 | 86,900 | Activation energy for electron transport (J mol−1) |
| PLAMOX | AEVO | 37,530 | 37,530 | 60,110 | Activation energy for RubP oxygenation (J mol−1) |
| PLAMOX | GSMIN | 21.9 | 5.0 | 60.0 | Minimum stomata conductivity (mmol H2O m−2 s−1) |
| PLAMOX | H2OREF_A | 0.5 | 0.2 | 1.0 | Relative available soil water content at which stomata conductance is affected |
| PLAMOX | H2OREF_GS | 1.0 | 0.2 | 1.0 | Relative available soil water content at which stomata are fully closed |
| PLAMOX | NFIX_RATE | 2.0 | 0.01 | 5.0 | Potential nitrogen fixation rate per plant dry matter tissue and day (kg N kg−1 DM d−1) |
| PLAMOX | N_DEF_FACTOR | 1.0 | 0.5 | 3.0 | Factor defines nitrogen deficiency |
| PLAMOX | ROOT | 0.45 | 0.3 | 0.65 | Plant root fraction |
| PLAMOX | SLAMAX | 15.0 | 13.0 | 25.0 | Specific leaf area in the shade (m2 kg−1) |
| PLAMOX | SLAMIN | 15.0 | 10.0 | 25.0 | Specific leaf area in under full light (m2 kg−1) |
| PLAMOX | SLOPE_GSA | 10.4 | 4.0 | 12.0 | Slope of foliage conductivity in response to assimilation in BERRY-BALL model |
| site | sks_upper | 1.0 | 0.357 | 3.57 | Saturated hydraulic conductivity for the uppermost layer |
| site | vangenuchten_n_upper | 1.1 | 1.1 | 1.2 | VanGenuchten parameter n (uppermost layer) |
| METRX | METRX_F_DECOMP_T_EXP_1 | 2 | 0.5 | 5 | Factor for temperature dependency of decomposition |
| METRX | METRX_KF_NIT_N2O | 0.003 | 0.001 | 0.2 | Maximum fraction of nitrified NH4 that goes to N2O |
| METRX | METRX_MIC_EFF | 0.848 | 0.1 | 2 | Microbial carbon use efficiency |
| Target Value | A1 | E1 | A2 | E2 | A3 | E3 |
|---|---|---|---|---|---|---|
| Biomass (kg DW ha−1) | 1056 | 1242 | 1017 | 1076 | 1114 | 1010 |
| CO2 (mg CO2 m−2 h−1) | 199.3 | 238.9 | 206.0 | 208.8 | 212.22 | 231.6 |
| N2O (µg N2O-N m−2 h−1) | 23.85 | 69.12 | 25.71 | 32.83 | 21.71 | 34.00 |
| SWC (%) | 6.90 | 6.65 | 7.61 | 6.76 | 6.83 | 6.26 |
| Target Value | A1/E1 | A2/E2 | A3/E3 |
|---|---|---|---|
| Biomass | 0.0433 | 0.431 | 0.110 |
| CO2 emissions | 3.992 × 10−9 | 0.0692 | 0.000293 |
| N2O emissions | 8.946 × 10−48 | 0.00240 | 1.536 × 10−22 |
| SWC | 1.255 × 10−32 | 2.671 × 10−8 | 0.00138 |
| Target Value | A1 | E1 | A2 | E2 | A3 | E3 | Data Points |
|---|---|---|---|---|---|---|---|
| Biomass (kg DW ha−1) | 2940 | 3373 | 3411 | 3400 | 3302 | 3593 | 27 |
| CO2 (mg CO2 m−2 h−1) | 384.8 | 462.1 | 393.5 | 407.3 | 358.8 | 415.1 | 966 |
| N2O (µg N2O-N m−2 h−1) | 8.80 | 32.25 | 10.42 | 13.02 | 9.48 | 16.25 | 1077 |
| SWC (%) | 34.8 | 38.4 | 44.4 | 42.1 | 38.4 | 39.5 | 2034 |



References
- World Meteorological Organization. The State of Greenhouse Gases in the Atmosphere Based on Global Observations through 2017; WMO: Geneva, Switzerland, 2018. [Google Scholar]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Weigel, H.-J.; Manderscheid, R. Crop growth responses to free air CO2 enrichment and nitrogen fertilization: Rotating barley, ryegrass, sugar beet and wheat. Eur. J. Agron. 2012, 43, 97–107. [Google Scholar] [CrossRef]
- Bindi, M.; Fibbi, L.; Miglietta, F. Free Air CO2 Enrichment (FACE) of grapevine (Vitis vinifera L.): II. Growth and quality of grape and wine in response to elevated CO2 concentrations. Eur. J. Agron. 2001, 14, 145–155. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; Finzi, A.C.; Norby, R.J.; Schlesinger, W.H. Soil Nitrogen Cycling Under Elevated Co2: A Synthesis of Forest Face Experiments. Ecol. Appl. 2003, 13, 1508–1514. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Su, B.; Currie, W.S.; Dukes, J.S.; Finzi, A.C.; Hartwig, U.; Hungate, B.; McMurtrie, R.E.; Oren, R.; Parton, W.J.; et al. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience 2004, 54, 731–739. [Google Scholar] [CrossRef] [Green Version]
- Gärdenäs, A.I.; Ågren, G.I.; Bird, J.A.; Clarholm, M.; Hallin, S.; Ineson, P.; Kätterer, T.; Knicker, H.; Nilsson, S.I.; Näsholm, T.; et al. Knowledge gaps in soil carbon and nitrogen interactions—From molecular to global scale. Soil Biol. Biochem. 2011, 43, 702–717. [Google Scholar] [CrossRef]
- Feng, Z.; Rütting, T.; Pleijel, H.; Wallin, G.; Reich, P.B.; Kammann, C.I.; Newton, P.C.D.; Kobayashi, K.; Luo, Y.; Uddling, J. Constraints to nitrogen acquisition of terrestrial plants under elevated CO2. Glob. Chang. Biol. 2015, 21, 3152–3168. [Google Scholar] [CrossRef]
- Van Groenigen, K.J.; Qi, X.; Osenberg, C.W.; Luo, Y.; Hungate, B.A. Faster Decomposition Under Increased Atmospheric CO2 Limits Soil Carbon Storage. Science 2014, 344, 508–509. [Google Scholar] [CrossRef]
- Cheng, W.; Johnson, D.W. Elevated CO2, rhizosphere processes, and soil organic matter decomposition. Plant Soil 1998, 202, 167–174. [Google Scholar] [CrossRef]
- Phillips, D.A.; Fox, T.C.; Six, J. Root exudation (net efflux of amino acids) may increase rhizodeposition under elevated CO2. Glob. Chang. Biol. 2006, 12, 561–567. [Google Scholar] [CrossRef]
- Terrer, C.; Vicca, S.; Hungate, B.A.; Phillips, R.P.; Prentice, I.C. Mycorrhizal association as a primary control of the CO2 fertilization effect. Science 2016, 353, 72–74. [Google Scholar] [CrossRef] [Green Version]
- Müller, C.; Sherlock, R.R.; Williams, P.H. Mechanistic model for nitrous oxide emission via nitrification and denitrification. Biol. Fertil. Soils 1997, 24, 231–238. [Google Scholar] [CrossRef]
- Ryan, M.; Müller, C.; Di, H.J.; Cameron, K.C. The use of artificial neural networks (ANNs) to simulate N2O emissions from a temperate grassland ecosystem. Ecol. Model. 2004, 175, 189–194. [Google Scholar] [CrossRef]
- Zhang, X.; Niu, G.-Y.; Elshall, A.S.; Ye, M.; Barron-Gafford, G.A.; Pavao-Zuckerman, M. Assessing five evolving microbial enzyme models against field measurements from a semiarid savannah—What are the mechanisms of soil respiration pulses? Geophys. Res. Lett. 2014, 41, 6428–6434. [Google Scholar] [CrossRef]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Chang. Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- Xu-Ri; Prentice, I.C.; Spahni, R.; Niu, H.S. Modelling terrestrial nitrous oxide emissions and implications for climate feedback. New Phytol. 2012, 196, 472–488. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Zhang, L.; Wang, Y.-P.; Yu, Q.; Eamus, D. Quantifying the effects of elevated CO2 on water budgets by combining FACE data with an ecohydrological model. Ecohydrology 2014, 7, 1574–1588. [Google Scholar] [CrossRef]
- Li, F.Y.; Newton, P.C.D.; Lieffering, M. Testing simulations of intra- and inter-annual variation in the plant production response to elevated CO2 against measurements from an 11-year FACE experiment on grazed pasture. Glob. Chang. Biol. 2014, 20, 228–239. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Medlyn, B.E.; Zaehle, S.; Walker, A.P.; Dietze, M.C.; Wang, Y.-P.; Luo, Y.; Jain, A.K.; El-Masri, B.; Hickler, T.; et al. Where does the carbon go? A model—Data intercomparison of vegetation carbon allocation and turnover processes at two temperate forest free-air CO2 enrichment sites. New Phytol. 2014, 203, 883–899. [Google Scholar] [CrossRef] [Green Version]
- Zaehle, S.; Medlyn, B.E.; De Kauwe, M.G.; Walker, A.P.; Dietze, M.C.; Hickler, T.; Luo, Y.; Wang, Y.-P.; El-Masri, B.; Thornton, P.; et al. Evaluation of 11 terrestrial carbon–nitrogen cycle models against observations from two temperate Free-Air CO2 Enrichment studies. New Phytol. 2014, 202, 803–822. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.P.; Hanson, P.J.; De Kauwe, M.G.; Medlyn, B.E.; Zaehle, S.; Asao, S.; Dietze, M.; Hickler, T.; Huntingford, C.; Iversen, C.M.; et al. Comprehensive ecosystem model-data synthesis using multiple data sets at two temperate forest free-air CO2 enrichment experiments: Model performance at ambient CO2 concentration. J. Geophys. Res. Biogeosci. 2014, 119, 2013JG002553. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.P.; Zaehle, S.; Medlyn, B.E.; De Kauwe, M.G.; Asao, S.; Hickler, T.; Parton, W.; Ricciuto, D.M.; Wang, Y.-P.; Wårlind, D.; et al. Predicting long-term carbon sequestration in response to CO2 enrichment: How and why do current ecosystem models differ? Glob. Biogeochem. Cycles 2015, 29, 2014GB004995. [Google Scholar] [CrossRef]
- Houska, T.; Kraft, P.; Liebermann, R.; Klatt, S.; Kraus, D.; Haas, E.; Santabarbara, I.; Kiese, R.; Butterbach-Bahl, K.; Müller, C.; et al. Rejecting hydro-biogeochemical model structures by multi-criteria evaluation. Environ. Model. Softw. 2017, 93, 1–12. [Google Scholar] [CrossRef]
- Kellner, J.; Multsch, S.; Houska, T.; Kraft, P.; Müller, C.; Breuer, L. A coupled hydrological-plant growth model for simulating the effect of elevated CO2 on a temperate grassland. Agric. For. Meteorol. 2017, 246, 42–50. [Google Scholar] [CrossRef]
- Liebermann, R.; Breuer, L.; Houska, T.; Klatt, S.; Kraus, D.; Haas, E.; Müller, C.; Kraft, P. Closing the N-Budget: How Simulated Groundwater-Borne Nitrate Supply Affects Plant Growth and Greenhouse Gas Emissions on Temperate Grassland. Atmosphere 2018, 9, 407. [Google Scholar] [CrossRef] [Green Version]
- Haas, E.; Klatt, S.; Fröhlich, A.; Kraft, P.; Werner, C.; Kiese, R.; Grote, R.; Breuer, L.; Butterbach-Bahl, K. LandscapeDNDC: A process model for simulation of biosphere–atmosphere–hydrosphere exchange processes at site and regional scale. Landsc. Ecol. 2013, 28, 615–636. [Google Scholar] [CrossRef]
- Grünhage, L.; Schmitt, J.; Hertstein, U.; Janze, S.; Peter, M.; Jäger, H.-J., III. Beschreibung der Versuchsfläche. In Auswirkungen Dynamischer Veränderungen der Luftzusammensetzung und des Klimas auf Terrestrische Ökosysteme in Hessen-II-Umweltbeobachtungs- und Klimafolgenforschungsstation Linden, Jahresbericht 1995; Schriftenreihe der Hessischen Landesanstalt für Umwelt 220; Umweltplanung, Arbeits- und Umweltschutz: Wiesbaden, Germany, 1996; ISBN 3-89026-236-8. [Google Scholar]
- Jäger, H.-J.; Schmidt, S.W.; Kammann, C.; Grünhage, L.; Müller, C.; Hanewald, K. The University of Giessen Free-Air Carbon dioxide Enrichment study: Description of the experimental site and of a new enrichment system. J. Appl. Bot. 2003, 77, 117–127. [Google Scholar]
- Patterson, D.E.; Smith, M.W. The use of time domain reflectometry for the measurement of unfrozen water content in frozen soils. Cold Reg. Sci. Technol. 1980, 3, 205–210. [Google Scholar] [CrossRef]
- Kammann, C.; Müller, C.; Grünhage, L.; Jäger, H.-J. Elevated CO2 stimulates N2O emissions in permanent grassland. Soil Biol. Biochem. 2008, 40, 2194–2205. [Google Scholar] [CrossRef]
- Kammann, C.; Grünhage, L.; Jäger, H.-J., II. N2O- und CH4-Flüsse in der bodennahen Atmosphäre eines extensiv genutzten Grünlandökosystems. In Auswirkungen Dynamischer Veränderungen der Luftzusammensetzung und des Klimas auf Terrestrische Ökosysteme in Hessen-III-Umweltbeobachtungs- und Klimafolgenforschungsstation Linden, Berichtszeitraum 1996–1999; Schriftenreihe der Hessischen Landesanstalt für Umwelt; Umweltplanung, Arbeits- und Umweltschutz: Wiesbaden, Germany, 2000; Volume 274, ISBN 3-89026-311-9. [Google Scholar]
- Grote, R.; Lehmann, E.; Brümmer, C.; Brüggemann, N.; Szarzynski, J.; Kunstmann, H. Modelling and observation of biosphere–atmosphere interactions in natural savannah in Burkina Faso, West Africa. Phys. Chem. Earth Parts ABC 2009, 34, 251–260. [Google Scholar] [CrossRef]
- Grote, R.; Lavoir, A.-V.; Rambal, S.; Staudt, M.; Zimmer, I.; Schnitzler, J.-P. Modelling the drought impact on monoterpene fluxes from an evergreen Mediterranean forest canopy. Oecologia 2009, 160, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Kraus, D.; Weller, S.; Klatt, S.; Santabárbara, I.; Haas, E.; Wassmann, R.; Werner, C.; Kiese, R.; Butterbach-Bahl, K. How well can we assess impacts of agricultural land management changes on the total greenhouse gas balance (CO2, CH4 and N2O) of tropical rice-cropping systems with a biogeochemical model? Agric. Ecosyst. Environ. 2016, 224, 104–115. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, D.; Weller, S.; Klatt, S.; Haas, E.; Wassmann, R.; Kiese, R.; Butterbach-Bahl, K. A new LandscapeDNDC biogeochemical module to predict CH4 and N2O emissions from lowland rice and upland cropping systems. Plant Soil 2014, 386, 125–149. [Google Scholar] [CrossRef]
- Dirnböck, T.; Kraus, D.; Grote, R.; Klatt, S.; Kobler, J.; Schindlbacher, A.; Seidl, R.; Thom, D.; Kiese, R. Substantial understory contribution to the C sink of a European temperate mountain forest landscape. Landsc. Ecol. under review.
- Cukier, R.I.; Fortuin, C.M.; Shuler, K.E.; Petschek, A.G.; Schaibly, J.H. Study of the sensitivity of coupled reaction systems to uncertainties in rate coefficients. I Theory. J. Chem. Phys. 1973, 59, 3873–3878. [Google Scholar] [CrossRef]
- Saltelli, A.; Tarantola, S.; Chan, K.P.-S. A Quantitative Model-Independent Method for Global Sensitivity Analysis of Model Output. Technometrics 1999, 41, 39–56. [Google Scholar] [CrossRef]
- McKay, M.D.; Beckman, R.J.; Conover, W.J. A Comparison of Three Methods for Selecting Values of Input Variables in the Analysis of Output from a Computer Code. Technometrics 1979, 21, 239–245. [Google Scholar]
- Beven, K.; Freer, J. Equifinality, data assimilation, and uncertainty estimation in mechanistic modelling of complex environmental systems using the GLUE methodology. J. Hydrol. 2001, 249, 11–29. [Google Scholar] [CrossRef]
- Houska, T.; Kraft, P.; Chamorro-Chavez, A.; Breuer, L. SPOTting Model Parameters Using a Ready-Made Python Package. PLoS ONE 2015, 10, e0145180. [Google Scholar] [CrossRef]
- Andresen, L.C.; Yuan, N.; Seibert, R.; Moser, G.; Kammann, C.I.; Luterbacher, J.; Erbs, M.; Müller, C. Biomass responses in a temperate European grassland through 17 years of elevated CO2. Glob. Chang. Biol. 2017, 24, 3875–3885. [Google Scholar] [CrossRef] [PubMed]
- Niklaus, P.A.; Spinnler, D.; Körner, C. Soil moisture dynamics of calcareous grassland under elevated CO2. Oecologia 1998, 117, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Morgan, J.A.; McMaster, G.S.; Ahuja, L.R.; Derner, J.D. Simulating Carbon Dioxide Effects on Range Plant Growth and Water Use with GPFARM-Range Model. Rangel. Ecol. Manag. 2015, 68, 423–431. [Google Scholar] [CrossRef]
- Kammann, C. Die Auswirkung Steigender Atmosphärischer CO2-Konzentrationen auf die Flüsse der Klimaspurengase N2O und CH4 in Einem Grünland-Ökosystem. Available online: http://geb.uni-giessen.de/geb/volltexte/2001/491/ (accessed on 7 March 2019).
- Xiang, S.-R.; Doyle, A.; Holden, P.A.; Schimel, J.P. Drying and rewetting effects on C and N mineralization and microbial activity in surface and subsurface California grassland soils. Soil Biol. Biochem. 2008, 40, 2281–2289. [Google Scholar] [CrossRef]
- Müller, C.; Stevens, R.J.; Laughlin, R.J.; Jäger, H.-J. Microbial processes and the site of N2O production in a temperate grassland soil. Soil Biol. Biochem. 2004, 36, 453–461. [Google Scholar] [CrossRef]
- Moser, G.; Gorenflo, A.; Brenzinger, K.; Keidel, L.; Braker, G.; Marhan, S.; Clough, T.J.; Müller, C. Explaining the doubling of N2O emissions under elevated CO2 in the Giessen FACE via in-field 15N tracing. Glob. Chang. Biol. 2018, 24, 3897–3910. [Google Scholar] [CrossRef]
- Kammann, C.; Grünhage, L.; Grüters, U.; Janze, S.; Jäger, H.-J. Response of aboveground grassland biomass and soil moisture to moderate long-term CO2 enrichment. Basic Appl. Ecol. 2005, 6, 351–365. [Google Scholar] [CrossRef]
- Denef, K.; Bubenheim, H.; Lenhart, K.; Vermeulen, J.; Van Cleemput, O.; Boeckx, P.; Müller, C. Community shifts and carbon translocation within metabolically-active rhizosphere microorganisms in grasslands under elevated CO2. Biogeosciences 2007, 4, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Guenet, B.; Lenhart, K.; Leloup, J.; Giusti-Miller, S.; Pouteau, V.; Mora, P.; Nunan, N.; Abbadie, L. The impact of long-term CO2 enrichment and moisture levels on soil microbial community structure and enzyme activities. Geoderma 2012, 170, 331–336. [Google Scholar] [CrossRef]
- Dämmgen, U.; Grünhage, L.; Schaaf, S. The precision and spatial variability of some meteorological parameters needed to determine vertical fluxes of air constituents. Landbauforschung Volkenrode 2005, 55, 29–37. [Google Scholar]
- Scholz-Seidel, C.D. UV 2 Messungen der Bulk-Depositionen sedimentierender anorganischer Spezies (September 1993 bis Dezember 1995). In Auswirkungen Dynamischer Veränderungen der Luftzusammensetzung und des Klimas auf Terrestrische Ökosysteme in Hessen-II-Umweltbeobachtungs- und Klimafolgenforschungsstation Linden, Jahresbericht 1995; Schriftenreihe der Hessischen Landesanstalt für Umwelt 220; Umweltplanung, Arbeits- und Umweltschutz: Wiesbaden, Germany, 1996. [Google Scholar]




| Target Variables | Threshold | Unit | Evaluated for Plots |
|---|---|---|---|
| Plant Biomass | 1300 | kg DW ha−1 | A1, A2, A3 |
| C–N ratio | 4.10 | - | |
| CO2 emissions | 200 | mg CO2 m−2 h−1 | |
| N2O emissions | 26.0 | µg N2O-N m−2 h−1 | |
| Soil Water Content (SWC) | 9.0 | vol.-% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liebermann, R.; Breuer, L.; Houska, T.; Kraus, D.; Moser, G.; Kraft, P. Simulating Long-Term Development of Greenhouse Gas Emissions, Plant Biomass, and Soil Moisture of a Temperate Grassland Ecosystem under Elevated Atmospheric CO2. Agronomy 2020, 10, 50. https://doi.org/10.3390/agronomy10010050
Liebermann R, Breuer L, Houska T, Kraus D, Moser G, Kraft P. Simulating Long-Term Development of Greenhouse Gas Emissions, Plant Biomass, and Soil Moisture of a Temperate Grassland Ecosystem under Elevated Atmospheric CO2. Agronomy. 2020; 10(1):50. https://doi.org/10.3390/agronomy10010050
Chicago/Turabian StyleLiebermann, Ralf, Lutz Breuer, Tobias Houska, David Kraus, Gerald Moser, and Philipp Kraft. 2020. "Simulating Long-Term Development of Greenhouse Gas Emissions, Plant Biomass, and Soil Moisture of a Temperate Grassland Ecosystem under Elevated Atmospheric CO2" Agronomy 10, no. 1: 50. https://doi.org/10.3390/agronomy10010050