 Mitchell H. Wright
Mitchell H. Wright Kati Geszvain
Kati Geszvain Véronique E. Oldham3,4
Véronique E. Oldham3,4 George W. Luther III
George W. Luther III Bradley M. Tebo
Bradley M. Tebo- 1Division of Environmental and Biomolecular Systems, Oregon Health & Science University, Portland, OR, United States
- 2Department of Biology, Lynchburg College, Lynchburg, VA, United States
- 3School of Marine Science and Policy, University of Delaware, Lewes, DE, United States
- 4Department of Marine Chemistry and Geochemistry, Woods Hole Oceanographic Institution, Woods Hole, MA, United States
The observation of significant concentrations of soluble Mn(III) complexes in oxic, suboxic, and some anoxic waters has triggered a re-evaluation of the previous Mn paradigm which focused on the cycling between soluble Mn(II) and insoluble Mn(III,IV) species as operationally defined by filtration. Though Mn(II) oxidation in aquatic environments is primarily bacterially-mediated, little is known about the effect of Mn(III)-binding ligands on Mn(II) oxidation nor on the formation and removal of Mn(III). Pseudomonas putida GB-1 is one of the most extensively investigated of all Mn(II) oxidizing bacteria, encoding genes for three Mn oxidases (McoA, MnxG, and MopA). P. putida GB-1 and associated Mn oxidase mutants were tested alongside environmental isolates Pseudomonas hunanensis GSL-007 and Pseudomonas sp. GSL-010 for their ability to both directly oxidize weakly and strongly bound Mn(III), and to form these complexes through the oxidation of Mn(II). Using Mn(III)-citrate (weak complex) and Mn(III)-DFOB (strong complex), it was observed that P. putida GB-1, P. hunanensis GSL-007 and Pseudomonas sp. GSL-010 and mutants expressing only MnxG and McoA were able to directly oxidize both species at varying levels; however, no oxidation was detected in cultures of a P. putida mutant expressing only MopA. During cultivation in the presence of Mn(II) and citrate or DFOB, P. putida GB-1, P. hunanensis GSL-007 and Pseudomonas sp. GSL-010 formed Mn(III) complexes transiently as an intermediate before forming Mn(III/IV) oxides with the overall rates and extents of Mn(III,IV) oxide formation being greater for Mn(III)-citrate than for Mn(III)-DFOB. These data highlight the role of bacteria in the oxidative portion of the Mn cycle and suggest that the oxidation of strong Mn(III) complexes can occur through enzymatic mechanisms involving multicopper oxidases. The results support the observations from field studies and further emphasize the complexity of the geochemical cycling of manganese.
Introduction
The geochemical cycling of manganese (Mn) in aquatic and terrestrial systems is largely governed by microbial oxidative and reductive processes, involving three oxidation states of Mn (II, III, and IV). Under anoxic conditions, bacterial reduction of insoluble Mn(III,IV) oxide minerals to soluble Mn(II) can occur as an energy generating process, involving the consumption of organic carbon coupled to oxides as terminal electron acceptors (Myers and Nealson, 1988). Conversely, in the presence of oxygen, bacteria are capable of oxidizing Mn(II) to Mn(III,IV) oxide though the purpose for this process is less understood (Tebo et al., 2004, 2005; Wright et al., 2016). While the processes that govern Mn(II) oxidation and solid Mn(III,IV) reduction have been well-studied, it has recently been observed that soluble Mn(III) bound to organic complexing ligands (Mn(III)-L) can dominate marine systems, comprising up to 100% of the total dissolved manganese (Oldham et al., 2017b). These complexes are defined as weak [Mn(III)-L(weak)] or strong [Mn(III)-L(strong)] as per their relative conditional stability constants (Oldham et al., 2015). This increased understanding of Mn(III) speciation has fundamentally changed the way we look at the role of environmental manganese. Mn(III)-L is capable of either donating or accepting electrons which subsequently significantly influences the redox chemistry of the surrounding environment (Luther et al., 2015). While research has highlighted the presence and significance of Mn(III)-L in these systems from a geochemical standpoint, little is known about the biological processes contributing to Mn(III)-L accumulation or removal.
In aquatic systems, the formation of Mn(III)-L is largely dependent on the surrounding redox conditions which are subject to change, not only through microbial influence, but also through various abiotic chemical processes. Environments containing high levels of hydrogen sulfide (H2S) can alter the relative prevalence of Mn(III,IV) oxide through redox transformations, producing reduced manganese and elemental sulfur (Herszage and Afonso, 2003). In iron rich systems, Fe2+ reduces Mn(III,IV) oxide at rates that can often outpace bacterial reduction (Siebecker et al., 2015). Furthermore, reactive superoxide () can drive the non-enzymatic oxidation of Mn(II) and is typically bacterially produced in aquatic systems (Learman et al., 2011; Diaz et al., 2013). Dissolved organic matter, such as humic substances, can also reduce solid phase manganese oxides, resulting in the formation of Mn(III)-L (Oldham et al., 2017a).
Abiotic oxidation of Mn(II) by O2 is slow, but it can be accelerated by surface reactions (Morgan, 2000). However, bacteria are considered the major driving force behind manganese oxide formation in most aquatic environments (Tebo et al., 2004, 2005). From an evolutionary standpoint, the process of manganese oxidation is both phylogenetically and enzymatically diverse and the bacteria that carry out these processes are ubiquitous in nature. Due to the significance of these microorganisms in the geochemical cycling of manganese in aquatic and terrestrial systems alike, numerous studies have been undertaken to further understand the mechanisms surrounding oxide formation across different phyla. Many of these studies have demonstrated enzymes to be involved in the oxidation of Mn(II), including: MofA in Leptothrix discophora SS-1 (Corstjens et al., 1997), MoxA in Pedomicrobium sp. ACM 3067 (Ridge et al., 2007) and MopA in Aurantimonas manganoxydans SI85-9A1 (Anderson et al., 2009). While these are illustrative of single-enzyme mechanisms, many bacteria oxidize manganese through more complex systems or use multiple enzymes. For example, Bacillus sp. strains PL-12 and SG-1 employ a multi-subunit enzyme which includes the MnxG multicopper oxidase (van Waasbergen et al., 1996; Dick et al., 2008; Butterfield et al., 2013).
Pseudomonas putida GB-1 is a prominent model Mn-oxidizing bacterium that has multiple Mn oxidase genes: mnxG, which encodes a multicopper oxidase similar (albeit with low sequence similarity) to that found in Bacillus sp. PL-12 and SG-1 (Dick et al., 2008; Geszvain et al., 2013); mcoA, encoding a unique multicopper oxidase (Geszvain et al., 2013); and mopA, an animal heme peroxidase similar to A. manganoxydans SI85-9A1 MopA and regulated by the flagellar synthesis gene fleQ (Geszvain et al., 2016). Many pseudomonads, including P. putida GB-1 (as well as another well-characterized strain, P. putida MnB1), produce pyoverdines (PVD), a class of fluorescent siderophores that chelate Fe(III) for uptake in environments with low soluble iron concentrations. Investigations analyzing PVD produced by P. putida MnB1 revealed that PVD binds Mn(III) with a strength equal to or greater than that with which it binds Fe(III) and that when present during Mn(II) oxidation, PVD binds to Mn(III) produced as an intermediate in the oxidation of Mn(II) to Mn(IV) (Parker et al., 2014). Although studies have demonstrated that Mn(III) occurs as an enzymatic intermediate in Mn(II) → Mn(IV) oxidation and that siderophores can bind Mn(III) with high affinity, no studies have investigated the effect of the presence of complexing ligands with different binding strengths on the rate and extent of Mn oxide formation during growth and Mn(II) oxidation by Mn(II)-oxidizing bacteria. Additionally, it is unknown whether bacteria can oxidize Mn(III) bound to heterologous siderophores, i.e., a siderophores produced by other organisms.
In this work, we show that Pseudomonas species are able to directly oxidize Mn(III)-L to Mn(IV). As relatively high levels of Mn(III)-L have been detected in aquatic environments, it is likely that under certain conditions, Mn(III)-L accumulates and therefore, Pseudomonas and other Mn-oxidizing bacteria would affect the formation and direct oxidation of Mn(III)-L (Oldham et al., 2017b). Through these data, we show that Pseudomonas spp. can not only directly oxidize Mn(III)-L, but that when Mn(II) is oxidized in the presence of excess ligand, it first accumulates as a Mn(III)-L complex before undergoing a second oxidation step to Mn(IV). Given their ubiquity in nature, we propose that manganese-oxidizing members of the genus Pseudomonas potentially play a role in both the accumulation and consumption of Mn(III)-L complexes in aquatic systems, and ultimately in the overall geochemical cycling of manganese.
Materials and Methods
Bacterial Strains
Pseudomonas hunanensis GSL-007 and Pseudomonas sp. GSL-010
Pseudomonas hunanensis GSL-007 and Pseudomonas sp. GSL-010 (Table 1) were isolated from water samples obtained from Station 23, St. Lawrence Estuary (48°42.032″N, 68°39.171″W). Briefly, sampled waters were diluted 1:1,000 using sterile 0.9% NaCl and streaked onto Minimal Media A (MMA) plates enriched with 100 μM Mn(III)-citrate. Individual colonies that exhibited Mn(III,IV) oxide formation (dark brown colonies) were subsequently selected, grown in MMA medium and the formation of Mn(III,IV) oxide reconfirmed. Phylogeny was determined using 16S rRNA sequencing and analysis as previously described (Farooqui et al., 2016). The 16S rRNA GenBank accession numbers for P. hunanensis GSL-007 and Pseudomonas sp. GSL-010 are KY471136 and KY471137 respectively. Pseudomonas sp. GSL-010 is currently under consideration for publication as a novel species and is deposited in the Japan Collection of Microorganisms (ID = 32154), the Korean Collection for Type Cultures (ID = 62392) and the NITE Biological Research Center (ID = 113027).
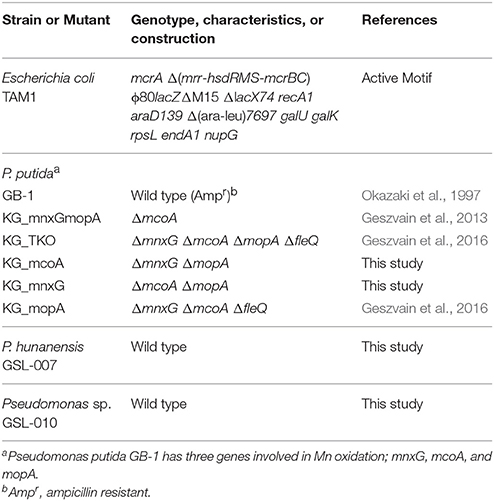
Table 1. Bacterial strains and mutants used in this study.
Pseudomonas putida GB-1
Pseudomonas putida GB-1 was originally isolated by the research group of Kenneth Nealson from Green Bay, Wisconsin, United States of America (Okazaki et al., 1997). Mn oxidation mutants of P. putida GB-1 were those as previously described (Geszvain et al., 2013, 2016) or constructed prior to use in this study. Briefly, deletion construct plasmids pKG168 or pKG170 (Table 2) were moved into the relevant mutant strains via conjugation. Counter-selection against the sacB locus on the gene replacement vector was used to isolate double recombinants; deletion of the target gene was confirmed by PCR across the deletion site (Table 3).
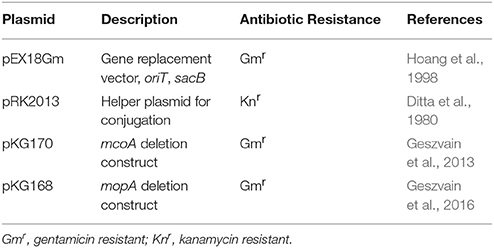
Table 2. Plasmids used in this study.
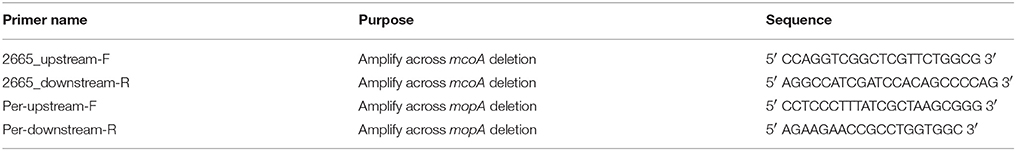
Table 3. Primers used in this study.
Culture Conditions
Strains used in this study are strictly aerobic mesophiles and were maintained in (Luria-Bertani Broth) LB medium, comprised of (per liter of diH2O): 10 g Bacto™ Tryptone (Becton, Dickinson and Company; MD, USA), 5 g yeast extract (Becton, Dickinson and Company; MD, USA) and 10 g NaCl with the pH adjusted to 7.5 using 1 M HCl/NaOH prior to autoclaving. For all quantitative analyses, Minimal Media A (MMA) was utilized and comprised of (per liter of Milli-Q H2O): 0.238 g (NH4)2SO4, 0.0602 g MgSO4, 0.0488 g CaCl2.2H2O, 0.0204 g KH2PO4 and 0.0284 g Na2HPO4 before autoclaving, with sterile 4.5 mL 20% glucose, 200 μL 5x Trace Elements Mix (Atlas, 2010), 1 mL 3.7 mM FeCl3.6H2O and 20 mL 1 M HEPES (pH 7.8) added after autoclaving with 100 μM Mn(II) (as MnCl2), Mn(III)-citrate, or Mn(III)-DFOB added as required. Autoclaving was completed under standard conditions (121°C for 15 min under 1.05 kg/cm2). All cultures were grown and maintained at 30°C and experiments were performed in triplicate. All glassware and other apparatus were acid washed prior to use.
Synthesis of Mn(III)-L Compounds
Mn(III)-Citrate
Mn(III)-citrate was used as a model weak Mn(III)-L complex. Stock solutions were prepared through the dissolution of Mn(III)-acetate in the presence of excess sodium citrate. Initially, 30 mM of sodium citrate (Fisher Scientific) was added to Milli-Q water and adjusted to pH 7 using trace metal clean HCl/NaOH. Ten millimolar Mn(III)-acetate was added, vigorously shaken, readjusted to pH 7 and allowed to sit in the absence of light for 24 h. Following this, the solution was 0.22 μm sterile filtered to remove any produced Mn(III,IV) oxide. Concentration of Mn(III)-citrate was determined spectrophotometrically at A428; ε = 310 L·mol−1·cm−1 (Duke, 1947).
Mn(III)-Desferrioxamine B [Mn(III)-DFOB]
Mn(III)-DFOB was used as a model strong Mn(III)-L complex. Stock solutions were prepared through air oxidation of Mn(II) in the presence of DFOB mesylate salt (Sigma Aldrich) using a method adapted from Duckworth and Sposito (2005). Initially, 2 mM DFOB mesylate salt was added to Milli-Q water and adjusted to pH 9 using trace metal clean HCl/NaOH. 1.1 mM MnCl2 was then added and the solution oxygenated for 24 h using filtered air. The solution was then readjusted to pH 8 and 0.22 μM sterile filtered to remove any produced Mn(III,IV) oxide. Final Mn(III)-DFOB concentration was determined spectrophotometrically at A310; ε = 2055 L·mol−1·cm−1. Mn(III)-DFOB recovery was ≥97% and produced Mn(III)-DFOB complex is stable through pH 7–11.3 and was used within 30 days of synthesis.
Chemical Analyses
Leucoberbelin Blue (LBB) Assay
Quantification of formed Mn(III)-L(weak) and Mn(III,IV) oxide was performed colorimetrically using an adapted LBB assay (Krumbein and Altman, 1973; Tebo et al., 2007). Stock solutions of 4% LBB were prepared using Milli-Q H2O, 24 mM NaOH and stored in the absence of light at 4°C. For use in assaying, LBB stock was diluted 1:100 in 1% glacial acetic acid (v/v) (storage life ~1 week). Standard curves of 10–100 μM were generated using known concentrations of fresh (≤6 h) KMnO4. Samples were visualized spectrophotometrically at A623 using a SpectraMax M2e (Molecular Devices LLC; CA, USA). LBB measures the oxidizing equivalents of Mn with an oxidation state > +II. Thus, one mole of Mn(VII) (KMnO4) will oxidize 5 moles of LBB. When calculating Mn(III,IV) oxide concentrations, it was assumed that all particulate Mn was MnO2.
Growth Consistency and Enumeration of Cells
For Mn(III)-L oxidation and production experiments, growth rates were synchronized to ensure consistency of results. Initially, frozen glycerol culture stocks (−80°C, 2:1 80% glycerol/log phase culture) were streaked onto LB agar plates. Individual colonies were then streaked to MMA agar plates and transferred to MMA after 24 h. For consistency of results, throughout all analytical experiments relative growth between replicates and between strains was monitored spectrophotometrically at OD600 (data not shown) to ensure the replicates for each given strain grew similarly. All experiments were performed in the absence of light unless otherwise specified.
Mn(III)-L Oxidation Experiments
For Mn(III,IV) oxide formation experiments, the Pseudomonas strains were grown in 5 mL aliquots of MMA media-containing 100 μM of either Mn(II), Mn(III)-citrate or Mn(III)-DFOB. Cultures were vacuum filtered through a 0.45 μm filter disk (Durapore), both the tube and disk were washed with 5 mL MilliQ H2O and the filter disk was returned to the original tube for analysis. Mn-oxides were observed on filter disks and were colorimetrically determined using 0.04% LBB. Sampling occurred at h 0, 8, 16, 24, 48, 96 for Mn(II)/Mn(III)-citrate and h 0, 24, 48, 72, 120, 168 for Mn(III)-DFOB. The maximum rate of Mn(III,IV) oxide formation was determined from the line between the two points that showed the steepest slope.
Mn(III)-L Formation Experiments
Our experimental design for Mn(III)-L formation experiments was formulated to both prevent iron limitation and siderophore production by the Pseudomonas isolates, as well as to avoid abiotic Mn(III)-DFOB production before biotic oxidation could proceed. To achieve this, the Pseudomonas strains or mutants were grown for 16 h in 300 mL of MMA media and monitored to ensure no siderophores were produced (fluorescence of pyoverdines). The growth consistency was confirmed and then each culture was amended with 100 μM Mn(II), as well as 2 mM Na-citrate, or 150 μM DFOB mesylate salt. These cultures were then dispensed into 5 mL aliquots. At various times, cultures were vacuum filtered through a 0.45 μm filter disk (Durapore), both tube and disk washed with 5 mL Milli-Q H2O and the filter disk was returned to the original tube for analysis as described above. Mn-oxides were observed on filter disks and were colorimetrically quantified using 0.04% LBB. Mn(III)-citrate present in the filtrate was determined using 0.04% LBB. Mn(III)-DFOB present in the filtrate was determined spectrophotometrically at A310; ε = 2,055 L·mol−1·cm−1. Sampling occurred at h 0, 8, 16, 24, 48, 96 for Mn(II)/Mn(III)-citrate and h 0, 24, 48, 72, 120, 168 for Mn(III)-DFOB. Unlike removal experiments, the Mn source and ligand were added after growth reached log phase to avoid chelation with free iron in the medium (see section Discussion).
Results
Mn(III)-L Oxidation by St. Lawrence Estuary Isolates
The ability of P. hunanensis GSL-007 (Figure 1A) and Pseudomonas sp. GSL-010 (Figure 1B) to directly oxidize Mn(III)-L over time was assessed using Mn(III)-citrate (weakly complexed ligand), Mn(III)-DFOB (strongly complexed ligand), with Mn(II) as a positive control. P. putida TKO (Figure 1G) was used as a negative control and did not exhibit any Mn oxidation. Both environmental isolates were able to oxidize all manganese species tested, with the rate of production and final yield from all three being higher for P. hunanensis GSL-007 than Pseudomonas sp. GSL-010 (Figures 1A,B; quantitative data shown in Table 4). Mn(II) and Mn(III)-citrate oxidation were oxidized at the same rate, with Mn(III,IV) oxide initially detected after 8–16 h and maximum oxidation detected after 48–96 h. However, oxidation of Mn(III)-DFOB was significantly slower, with initial oxidation evident after 48–120 h and maximum oxidation observed after 168 h. There were also significant differences observed in the final quantity of Mn(III,IV) oxide formed depending on the initial substrate, with Mn(II) and Mn(III)-citrate producing equal amounts of Mn(III,IV) oxide and Mn(III)-DFOB producing significantly less (Table 4).
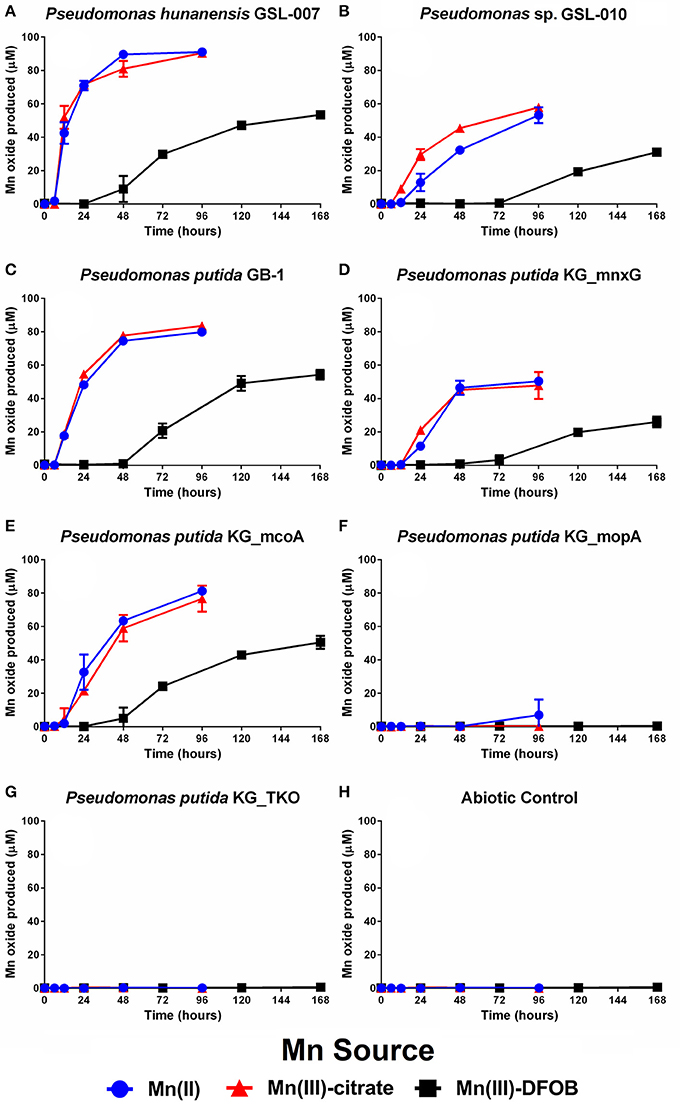
Figure 1. Mn(II), Mn(III)-citrate, and Mn(III)-DFOB oxidation over time by (A) P. hunanensis GSL-007, (B) Pseudomonas sp. GSL-010, (C) P. putida GB-1 alongside associated mutants—(D) P. putida KG_mnxG (producing only MnxG), (E) P. putida KG_mcoA (producing only McoA) and (F) P. putida KG_mopA (producing only MopA)—as measured by Mn(III,IV) oxide formation. Mn(II) was used as a positive control with (G) sterile medium and (H) P. putida TKO (devoid of any Mn oxidase genes) serving as negative controls. Grown in minimal media at 30°C containing 100 μM of relevant Mn compound. Relative growth between bacterial isolates was normalized by OD600 (results not shown).
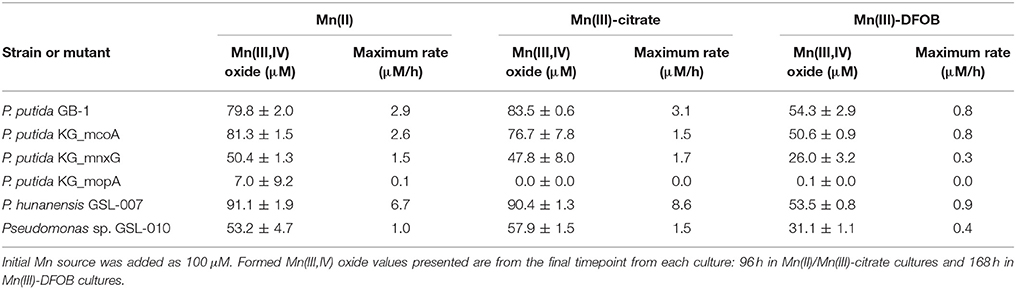
Table 4. Final yields of Mn(III,IV) oxide formed through bacterial Mn(II/III) oxidation (see Figure 1) and maximum rates.
Mn(III)-L Oxidation Between Pseudomonas putida GB-1 and Mn Oxidase Mutants
The specificity of P. putida enzymes MnxG, McoA, and MopA toward Mn(III) was tested using the wildtype strain GB-1, alongside mutants P. putida KG_mnxG (only produces MnxG), P. putida KG_mcoA (only produces McoA) and P. putida KG_mopA (only produces MopA) (Figures 1C–F; Table 1). Mn(II) and Mn(III) oxidation were observed with P. putida GB-1, P. putida KG_mnxG and P. putida KG_mcoA; however, P. putida KG_mopA failed to oxidize Mn(III) and showed only limited Mn(II) oxidation after 96 h (Figure 1F; Table 4). Because P. putida KG_mopA showed little to no activity under these conditions, the remainder of the results describe just the wildtype, P. putida KG_mnxG and P. putida KG_mcoA strains. As was seen with the St. Lawrence estuary isolates, the P. putida GB-1 strains oxidized Mn(II) and Mn(III)-citrate at similar rates, while oxidation of Mn(III)-DFOB was much slower (Figures 1C–E; Table 4). For each of the three tested Mn compounds, the strain P. putida KG_mnxG produced lower quantities of Mn(III,IV) oxide than wildtype, P. putida KG_mcoA and P. hunanensis GSL-007 but roughly similar amounts as Pseudomonas sp. GSL-010 (Table 4).
Mn(III)-Citrate Accumulation
Mn(II) oxidation by P. putida GB-1, P. hunanensis GSL-007 and Pseudomonas sp. GSL-010, in the presence of excess sodium citrate, was tested to detect accumulation of intermediary, weakly-bound Mn(III)-L (Figure 2; Table 5). There was no Mn(III,IV) formation observed in the triple knockout mutant (P. putida TKO). Mn(III)-citrate formation was detected at 8–16 h in all oxidizing cultures and remained at low concentrations during Mn(III,IV) oxide formation; however, the Mn(III)/Mn(IV) ratios observed were different for the tested isolates. For P. putida GB-1, Mn(III,IV) oxide was the primary product detected (40.6 ± 4.0 μM) after 16 h, compared to Mn(III)-citrate (19.0 ± 3.8 μM). The opposite trend was observed in Pseudomonas sp. GSL-010, with Mn(III)-citrate (27.1 ± 1.5 μM) present at greater concentrations than Mn(III,IV) oxide (7.9 ± 1.4 μM). Relatively similar levels of Mn(III)-citrate (31.5 ± 0.8 μM) and Mn(III,IV) oxide (45.5 ± 0.3 μM) were detected in P. hunanensis GSL-007 at initial formation after 8 h. After 48 h, Mn(III,IV) oxide formation was observed (69.6–85.6 μM) under all conditions and almost no Mn(III)-citrate (0.2–3.4 μM) was detected.
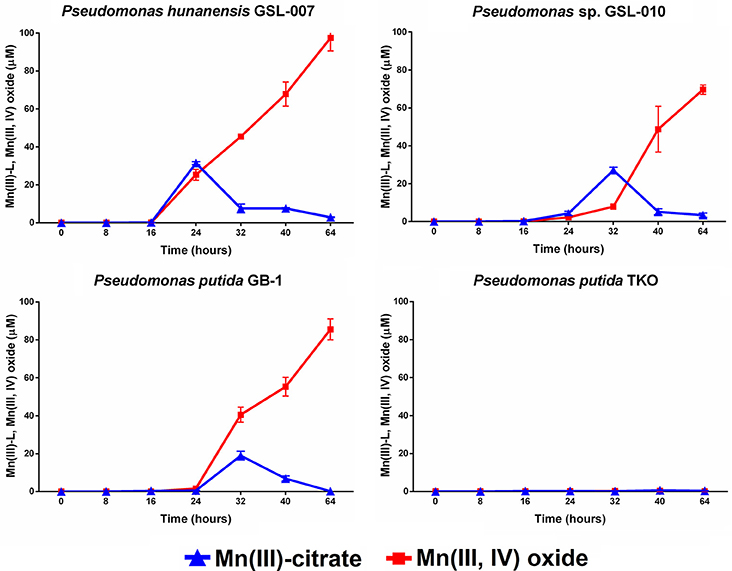
Figure 2. Mn(III)-citrate and Mn(III,IV) oxide accumulation during Mn(II) oxidation from Pseudomonas putida GB-1, P. hunanensis GSL-007 and Pseudomonas sp. GSL-010. P. putida TKO served as a negative control. Cultures were grown for 16 h in MMA at 30°C before addition of 100 μM MnCl2 and 2 mM sodium citrate. Relative growth between bacterial isolates was normalized by OD600 (results not shown).
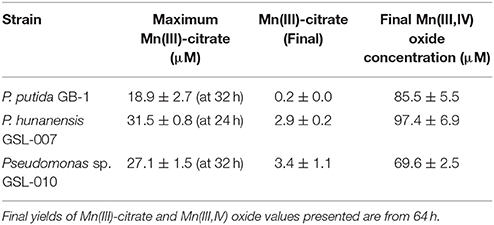
Table 5. Maximum Mn(III)-citrate observed along with final yields of Mn(III)-citrate and Mn(III,IV) oxide formed through bacterial Mn(II) oxidation (see Figure 2).
Mn(III)-DFOB Accumulation
Mn(II) oxidation by P. putida GB-1, P. hunanensis GSL-007 and Pseudomonas sp. GSL-010 in the presence of excess DFOB was tested to assess accumulation of intermediary, strongly-bound Mn(III)-L (Figure 3; Table 6). Substantial accumulation of Mn(III)-DFOB was observed for both the abiotic and P. putida TKO negative controls. DFOB is known to promote the oxidation of Mn(II) by oxygen resulting in the formation of Mn(III)-DFOB (see section Materials and Methods; Duckworth and Sposito, 2005) However, bacterial oxidation proceeded at greater rates than the abiotic control, with all isolates showing complete (≥97.5 μM) Mn(III)-DFOB formation after 32 h. Comparatively, at the same timepoint abiotic Mn(III)-DFOB formation was observed at levels of 36.0 ± 3.0 μM. Similar trends were observed between all bacterial isolates, with initial Mn(III)-DFOB formation observed followed by Mn(III,IV) oxide formation 40–48 h after initial inoculation. Mn(III,IV) oxide formation was not observed in the abiotic control but was observed in all oxidizing cultures (4.7–13.4 μM) and occurred as Mn(III)-DFOB concentrations decreased, indicative of a secondary oxidation step. There was no significant increase in Mn(III)-DFOB formation in the triple knockout mutant (P. putida TKO) relative to the abiotic control.
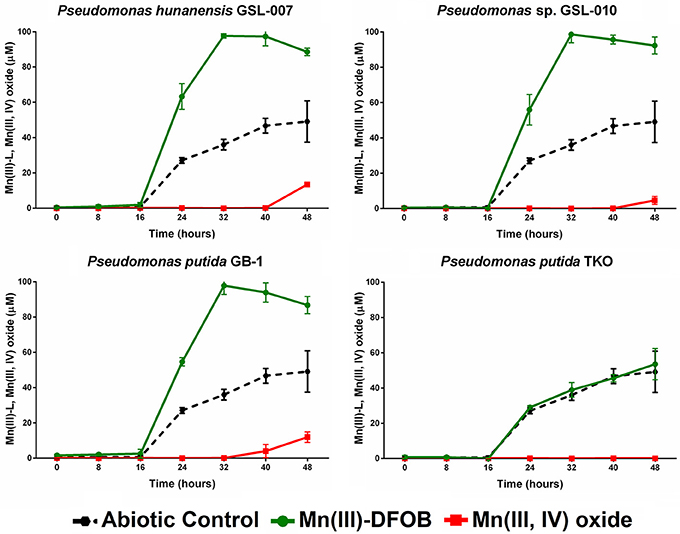
Figure 3. Mn(III)-DFOB and Mn(III,IV) oxide accumulation during Mn(II) oxidation from Pseudomonas putida GB-1, P. hunanensis GSL-007 and Pseudomonas sp. GSL-010. P. putida TKO was used as a negative control and abiotic oxidation was measured over time in culture-free medium. Cultures were grown for 16 h in MMA at 30°C before being inoculated with 100 μM MnCl2 and 150 μM DFOB. Relative growth between bacterial isolates was normalized by OD600 (results not shown).
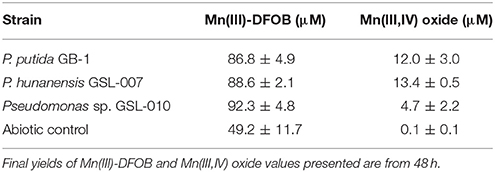
Table 6. Final yields of Mn(III)-DFOB and Mn(III,IV) oxide formed through bacterial Mn(II) oxidation (see Figure 3).
Discussion
Research into bacterial Mn(II)-oxidizing enzymes has highlighted that these oxidases produce Mn(III) as an intermediate during the formation of particulate Mn(III,IV) oxide and are also capable of directly oxidizing Mn(III). The multicopper oxidase (MCO) in Bacillus sp. PL-12 (containing a MnxG subunit with low levels of homology to P. putida GB-1) catalyzes two single one-electron oxidation steps, from Mn(II) to Mn(III) and then Mn(III) to Mn(IV) (Soldatova et al., 2017a,b). Similarly, animal heme peroxidases produced by A. manganoxydans SI85-9A1, Erythrobacter sp. SD-21 and Roseobacter sp. Azwk-3b oxidized Mn(II) to Mn(III), which subsequently disproportionated or was further oxidized to Mn(IV) (Anderson et al., 2009; Andeer et al., 2015). Though illustrative of Mn(III) oxidation/complexation, these studies used purified protein or the exosporium, whereas this study examines these processes when carried out by bacterial cultures. Given the frequent occurrence of Pseudomonas in different environments, members of the genus were selected as model organisms to further understand the bacterial influence on Mn(III)-L formation and removal in aquatic systems. This included species isolated from the St. Lawrence Estuary (P. hunanensis GSL-007 and Pseudomonas sp. GSL-010) as well as P. putida GB-1, an Mn(II)-oxidizer long studied as a laboratory model. P. putida GB-1 encodes two multicopper oxidases (MnxG and McoA) and an animal heme peroxidase (MopA) implicated in Mn oxidation (Geszvain et al., 2013, 2016). Mn(III) oxidation in P. putida GB-1 was examined with mutants expressing only one of these genes in order to determine whether a specific oxidase, or a group of enzymes, interacts with Mn(III).
DFOB, a siderophore produced by some bacteria and fungi, was selected as a model strong complexing ligand because siderophores are produced by a wide range of bacteria and fungi under low iron stress in environments (such as aquatic systems) where iron is not always readily available and are environmentally significant (Neilands, 1995). Indeed, both P. putida GB-1 and the closely related strain, P. putida MnB1, produce siderophores under iron-starved conditions; these siderophores were found to have a high binding affinity for Mn(III) and inhibit Mn(IV) formation (Parker et al., 2004, 2014). Given that siderophores (including DFOB and the P. putida-produced siderophores) have a natural affinity to bind to iron, one consideration in our experimental design was the necessity to avoid chelation between the DFOB and free iron in the culture media during growth phase. A second consideration was that abiotic Mn(III)-DFOB synthesis can occur through the air oxidation of Mn(II) in the presence of DFOB (Figure 3; Duckworth and Sposito, 2005). We saw no evidence of siderophore production (fluorescence of pyoverdines known to be produced by P. putida GB-1) during the growth phase of our incubations and although abiotic oxidation of Mn(II) was observed in the DFOB experiments, Mn-oxidizing bacteria formed Mn(III)-DFOB at a faster rate. The low levels of Mn(III,IV) oxide production observed after 48 h relative to the Mn(II) and Mn(III)–citrate experiments is likely due to the reduction of Mn(III,IV) oxide by DFOB (Oldham et al., 2015) resulting in an apparent decrease in the rate of formation due to a decrease in Mn(III,IV) oxide accumulation.
Direct Mn(III)-L oxidation was observed in wild type cultures of P. hunanensis GSL-007, Pseudomonas sp. GSL-010, and P. putida GB-1 as well as by mutants P. putida KG_mcoA and P. putida KG_mnxG (Figure 1). Conversely, no Mn(III)-L oxidation was detected in P. putida KG_mopA cultures and very little particulate Mn(III,IV) oxide formation was observed in the Mn(II) control. These results contrast previous investigations of MopA, which after 72 h showed significant Mn(III,IV) oxide formation (Geszvain et al., 2016). This difference can be explained by the effect of light. Our experiments were performed in the dark, while those reported by Geszvain et al. (2016) were exposed to ambient room light. A separate experiment with P. putida KG_mopA cell-free extracts showed that MopA produces very little Mn(III,IV) oxide in the dark compared to relatively high levels in the light (Data Sheet 1). A similar effect of light has been demonstrated in Roseobacter sp. AzwK-3b, another species that utilizes a MopA homolog in Mn oxidation (Hansel and Francis, 2006). However, by conducting our experiments in the dark, we intended to better replicate the environmental conditions from which our isolates were obtained and to avoid any potential photoreactions with the HEPES or photoreduction of Mn in our growth media.
One longstanding question surrounding P. putida GB-1 is the rationale behind the presence of multiple Mn oxidase enzymes, with one hypothesis being that they are activated under different environmental conditions (Geszvain et al., 2016). Given the dependence on light, MopA may be expressed predominately in the upper regions of the oxic zone of aquatic environments where light is abundant, whereas in the lower regions where light is absent, it may not be expressed. However, this hypothesis assumes that MopA-driven oxidation is beneficial in the euphotic zone and not at deeper levels. Even though the Mn(II)-oxidizing peroxidases from both P. putida GB-1 and Roseobacter sp. AzwK-3b are thought to perform single, one electron oxidation reactions [Mn(II) to Mn(III)], both appear to produce Mn(III,IV) oxide. Similarly, Mn(III/IV) oxide formation was observed in our MopA light/dark experiments through the enzymatic oxidation of Mn(II). How peroxidases catalyze formation of solid Mn(III,IV) oxide requires further investigation.
Citrate was selected as a model weak complexing ligand because P. hunanensis GSL-007 and Pseudomonas sp. GSL-010 were originally isolated by their ability to form Mn(III,IV) oxide from Mn(III)-citrate. We observed that in the presence of excess citrate, oxidation of Mn(II) would lead to complexation of Mn(III) to form Mn(III)-citrate before a later secondary oxidation step to form Mn(III,IV) oxide. Though our results highlight the first instance of direct bacterial oxidation causing Mn(III)-L accumulation, trapping of an Mn(III)-intermediate using the ligand pyrophosphate (PP) and subsequent oxidation to Mn(IV) has been demonstrated using exosporium containing the intact Mnx complex of Bacillus sp. SG-1 (Webb et al., 2005; Soldatova et al., 2012) and the purified Mnx complex from Bacillus sp. PL-12 (Soldatova et al., 2017a,b). Given the weak complexation of Mn(III) to PP, we predicted that Mn(III)-citrate would behave in a similar manner. Because the HEPES we used in our media can abiotically reduce Mn(III)-pyrophosphate (Kostka and Luther, 1994), we used Mn(III)-citrate as the model weak complexing ligand. When we substituted a phosphate buffer for HEPES and tested for pyrophosphate, all isolates were capable of Mn(III,IV) oxide formation using Mn(III)-pyrophosphate (results not shown). There was no observed abiotic reduction of Mn(III)-citrate by HEPES under the experimental conditions utilized in this study.
While our findings prompt a reevaluation of the process of bacterial Mn(II, III) oxidation in aquatic environments, it is important to note that recent investigations into reductive processes have also fundamentally changed our understanding of these systems. Recently, it was learned that Mn(IV) reduction requires an initial solubilization step and that Mn(IV) reduction is in fact a series of two consecutive one-electron transfer reactions, with soluble Mn(III) produced as intermediary species (Lin et al., 2012). Subsequent investigations of a bacterial isolate, Shewanella oneidensis, highlighted the process of direct Mn(III) reduction and provided insight into the associated mechanisms (Szeinbaum et al., 2014). Given that reduction also involves formation of an Mn(III) intermediate, complexation to surrounding ligands is likely to occur as is observed in manganese oxidation. Previous work from members of our group, Oldham et al. (2015, 2017a), highlighted that one of the potential Mn(III)-L formation pathways is through the bacterial reduction of Mn(IV) in the presence of suitable ligands, resulting in complexation and formation of Mn(III)-L. Thus, while our results highlight the potential significance of Mn oxidizers in the formation and removal of Mn(III) in aquatic systems, any reassessment of the current cycling model should also incorporate Mn-reducing bacteria.
From an aquatic sciences standpoint, traditional research has focused almost exclusively on the reduction of Mn(IV) and the oxidation of Mn(II) based on operationally defined soluble, reduced Mn(II) (which passes through a 0.2 μm or 0.45 μm filter) and oxidized particulate Mn(III,IV) oxide trapped on the filters. However, an increased understanding into the significance of Mn(III) has prompted a paradigm shift in how different manganese species are cycled within environmental systems (Trouwborst et al., 2006; Madison et al., 2011; Luther et al., 2015; Oldham et al., 2017a,b). Though it has previously been suggested that bacterial oxidative processes likely drive the presence of Mn(III)-L in aquatic systems (Webb et al., 2005), this paper shows the first instance of direct bacterial Mn(III)-L formation and removal. Evidence of Mn(III)-L oxidation by MnxG and McoA, but not MopA suggests that this step may be predominately MCO driven, at least in the absence of light. While this research focused solely on Pseudomonas spp., given the ubiquity of manganese-oxidizing bacteria in aquatic environments and the influence they have on Mn(II), Pseudomonas and many other genera are also likely some of the driving forces behind Mn(III)-L formation and removal in the environment. Investigating whether these findings apply to other Mn(II) oxidizers may provide further insight into their influence on the cycling of Mn(III) in aquatic systems.
Author Contributions
The concepts being tested in this manuscript resulted from the collaboration between MW, BT, VO, and GL on Mn(III) cycling in the environment. MW and BT designed and performed the Mn oxidation experiments. KG generated the mutants and completed the light/dark experiments. VO and GL provided guidance on the Mn(III) analyses. All authors participated in the interpretation of the results and preparation of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors acknowledge the critical comments of Dr. Christine Romano and Mr. John Hanna throughout the manuscript preparation and are greatly appreciative for their input. This work was funded by grants from the Chemical Oceanography program of the National Science Foundation (OCE-1558738 and OCE-1558692).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00560/full#supplementary-material
References
Andeer, P. F., Learman, D. R., McIlvin, M., Dunn, J. A., and Hansel, C. M. (2015). Extracellular haem peroxidases mediate Mn(II) oxidation in a marine Roseobacter bacterium via superoxide production. Environ. Microbiol. 17, 3925–3936. doi: 10.1111/1462-2920.12893
Anderson, C. R., Johnson, H. A., Caputo, N., Davis, R. E., Torpey, J. W., and Tebo, B. M. (2009). Mn(II) Oxidation is catalyzed by heme peroxidases in “Aurantimonas manganoxydans”strain SI85-9A1 and Erythrobacter sp. strain SD-21. Appl. Environ. Microbiol. 75, 4130–4138. doi: 10.1128/AEM.02890-08
Butterfield, C., Soldatova, A. V., Lee, S. W., Spiro, T. G., and Tebo, B. M. (2013). Mn(II,III) oxidation and MnO2 mineralization by an expressed bacterial multicopper oxidase. Proc. Natl. Acad. Sci. U.S.A. 110, 11731–11735. doi: 10.1073/pnas.1303677110
Corstjens, P. L. A. M., de Vrind, J. P. M., Goosen, T., and de Vrind-de Jong, E. W. (1997). Identification and molecular analysis of the Leptothrix discophora SS-1 mofA gene, a gene putatively encoding a manganese-oxidizing protein with copper domains. Geomicrobiol. J. 14, 91–108. doi: 10.1080/01490459709378037
Diaz, J. M., Hansel, C. M., Voelker, B. M., Mendes, C. M., Andeer, P. F., and Zhang, T. (2013). Widespread production of extracellular superoxide by heterotrophic bacteria. Science 340, 1223–1226. doi: 10.1126/science.1237331
Dick, G. J., Torpey, J. W., Beveridge, T. J., and Tebo, B. M. (2008). Direct identification of a bacterial manganese(II) oxidase, the multicopper oxidase MnxG, from spores of several different marine Bacillus species. Appl. Environ. Microbiol. 74, 1527–1534. doi: 10.1128/AEM.01240-07
Ditta, G., Stanfield, S., Corbin, D., and Helinski, D. R. (1980). Broad host range DNA cloning system for gram-negative bacteria: construction of a gene bank of Rhizobium meliloti. Proc. Natl. Acad. Sci. U.S.A. 77, 7347–7351. doi: 10.1073/pnas.77.12.7347
Duckworth, O. W., and Sposito, G. (2005). Siderophore-manganese(III) interactions. I. air-oxidation of manganese(III) promoted by desferrioxamine, B. Envrion. Sci. Tech. 39, 6037–6044. doi: 10.1021/es050275k
Duke, F. R. (1947). The theory and kinetics of specific oxidation. I. The trivalent manganese–oxalate reaction. J. Am. Chem. Soc. 69, 2885–2888. doi: 10.1021/ja01203a073
Farooqui, S. M., Wright, M. H., and Greene, A. C. (2016). Aliidiomarina minuta sp. nov., a haloalkaliphilic bacterium that forms ultra-small cells under non-optimal conditions. Antonie van Leeuwenhoek 109, 83–93. doi: 10.1007/s10482-015-0611-3
Geszvain, K., McCarthy, J. K., and Tebo, B. M. (2013). Elimination of manganese(II,III) oxidation in Pseudomonas putida GB-1 by a double knockout of two putative multicopper oxidase genes. Appl. Environ. Microbiol. 79, 357–366. doi: 10.1128/AEM.01850-12
Geszvain, K., Smesrud, L., and Tebo, B. M. (2016). Identification of a third Mn(II) oxidase enzyme in Pseudomonas putida GB-1. Appl. Environ. Microbiol. 82, 3774–3782. doi: 10.1128/AEM.00046-16
Hansel, C. M., and Francis, C. A. (2006). Coupled photochemical and enzymatic Mn(II) oxidation pathways of a planktonic Roseobacter-like bacterium. Appl. Environ. Microbiol. 72, 3543–3549. doi: 10.1128/AEM.72.5.3543-3549.2006
Herszage, J., and Afonso, M. (2003). Mechanism of hydrogen sulfide oxidation by manganese(IV) oxide in aqueous solutions. Langmuir 19, 9684–9692. doi: 10.1021/la034016p
Hoang, T. T., Karkhoff-Schweizer, R. R., Kutchma, A. J., and Schweizer, H. P. (1998). A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 212, 77–86. doi: 10.1016/S0378-1119(98)00130-9
Kostka, J. E., Luther, G. W. III., and Nealson, K. H. (1994). Chemical and biological reduction of Mn (III) -pyrophosphate complexes: potential importance of dissolved Mn (III) as an environmental oxidant. Geochim. Cosmochim. Acta 59, 885–894.
Krumbein, W. E., and Altman, H. J. (1973). A new method for detection and enumeration of manganese oxidizing and reducing microorganisms. Helgol. Meeresunters. 25, 347–356. doi: 10.1007/BF01611203
Learman, D. R., Voelker, D. M., Vazquez-Rodriguez, A. I., and Hansel, C. M. (2011). Formation of manganese oxides by bacterially generated superoxide. Nat. Geosci. 4, 95–98. doi: 10.1038/ngeo1055
Lin, H., Szeinbaum, N. H., DiChristina, T. J., and Taillefert, M. (2012). Microbial Mn(IV) reduction requires an initial one-electron reductive solubilization step. Geochim. Cosmochim. Acta. 99, 179–192. doi: 10.1016/j.gca.2012.09.020
Luther, G. W. III., Madison, A. S., Mucci, A., Sundby, B., and Oldham, V. E. (2015). A kinetic approach to assess the strengths of ligands bound to soluble Mn(III). Mar. Chem. 173, 93–99. doi: 10.1016/j.marchem.2014.09.006
Madison, A. S., Muchi, A., Sundby, B., and Oldham, V. E. (2011). Simultaneous determination of soluble manganese(III), manganese(II) and total manganese in natural (pore)waters. Talanta 84, 374–381. doi: 10.1016/j.talanta.2011.01.025
Morgan, J. J. (2000). “Manganese in natural waters and earth's crust: Its availability to organisms,” in Metal Ions in Biological Systems Manganese and Its Role in Biological Processes, Vol. 37, eds A. Sigel and H. Sigel (New York, NY: Marcel Dekker), 1–33.
Myers, C. R., and Nealson, K. H. (1988). Bacterial manganese reduction and growth with manganese oxide as the sole electron acceptor. Science 240, 1319–1321. doi: 10.1126/science.240.4857.1319
Neilands, J. B. (1995). Siderophores: structure and function of microbial iron transport compounds. J. Biol. Chem. 270, 26723–26726. doi: 10.1074/jbc.270.45.26723
Okazaki, M., Sugita, T., Shimizu, M., Ohode, Y., Iwamoto, K., de Vrind-de Jong, E. W., et al. (1997). Partial purification and characterization of manganese-oxidizing factors of Pseudomonas fluorescens GB-1. Appl. Environ. Microbiol. 63, 4793–4799.
Oldham, V. E., Jones, M. R., Tebo, B. M., and Luther, G. W. (2017a). Oxidative and reductive processes contributing to manganese cycling at oxic-anoxic interfaces. Mar. Chem. 195, 122–128. doi: 10.1016/j.marchem.2017.06.002
Oldham, V. E., Mucci, A., Tebo, B. M., and Luther, G. W. (2017b). Soluble Mn(III)-L complexes are abundant in oxygenated waters and stabilized by humic ligands. Geochim. Cosmochim. Acta 199, 238–246. doi: 10.1016/j.gca.2016.11.043
Oldham, V. E., Owings, S. M., Jones, M. R., Tebo, B. M., and Luther, G. W. (2015). Evidence for the presence of strong Mn(III)-binding ligands in the water column of the Chesapeake Bay. Mar. Chem. 171, 58–66. doi: 10.1016/j.marchem.2015.02.008
Parker, D. L., Lee, S.-W., Geszvain, K., Davis, R. E., Gruffaz, C., Meyer, J.-M., et al. (2014). Pyoverdine synthesis by the Mn(II)-oxidizing bacterium Pseudomonas putida GB-1. Front. Microbiol. 5:202. doi: 10.3389/fmicb.2014.00202
Parker, D. L., Sposito, G., and Tebo, B. M. (2004). Manganese(III) binding to a pyoverdine siderophore produced by a manganese(II)-oxidizing bacterium. Geochim. Cosmochim. Acta 68, 4809–4820. doi: 10.1016/j.gca.2004.05.038
Ridge, J. P., Lin, M., Larsen, E. I., Fegan, M., McEwan, A. G., and Sly, L. I. (2007). A multicopper oxidase is essential for manganese oxidation and laccase-like activity in Pedomicrobium sp. ACM 3067. Environ. Microbiol. 9, 944–953. doi: 10.1111/j.1462-2920.2006.01216.x
Siebecker, M., Madison, A. S., and Luther, G. W. (2015). Reduction kinetics of polymeric (soluble) manganese (iv) oxide (MnO2) by ferrous iron (Fe2+). Aquat. Geochem. 21, 143–158. doi: 10.1007/s10498-015-9257-z
Soldatova, A. V., Butterfield, C., Oyerinde, O. F., Tebo, B. M., and Spiro, T. G. (2012). Multicopper oxidase involvement in both Mn(II) and Mn(III) oxidation during bacterial formation of MnO2. J. Biol. Inorg. Chem. 17, 1151–1158. doi: 10.1007/s00775-012-0928-6
Soldatova, A. V., L. Tao, C. A., Romano, T. A., Stich, W. H., Casey, R. D., Britt, B. M., et al. (2017a). Mn(II) oxidation by the multicopper oxidase complex Mnx: a binuclear activation mechanism. J. Am. Chem. Soc. 139, 11369–11380. doi: 10.1021/jacs.7b02771
Soldatova, A. V., Romano, C. A., Tao, L., Stich, T. A., Casey, W. H., Britt, R. D., et al. (2017b). Mn(II) oxidation by the multicopper oxidase complex Mnx: a coordinated two-stage Mn(II)/(III) and Mn(III)/(IV) Mechanism. J. Am. Chem. Soc. 139, 11381–11391. doi: 10.1021/jacs.7b02772
Szeinbaum, N., Burns, J. L., and DiChristina, T. J. (2014). Electron transport and protein secretion pathways involved in Mn(III) reduction by Shewanella oneidensis. Environ. Microbiol. Rep. 6, 490–500. doi: 10.1111/1758-2229.12173
Tebo, B. M., Bargar, J. R., Clement, B. G., Dick, G. J., Murray, K. J., Parker, D., et al. (2004). Biogenic manganese oxides: properties and mechanisms of formation. Annu. Rev. Earth Planet. Sci. 32, 287–328. doi: 10.1146/annurev.earth.32.101802.120213
Tebo, B. M., Clement, B. G., and Dick, G. J. (2007). “Biotransformations of manganese,” in Manual of Environmental Microbiology, 3rd Edn, eds, C. J. Hurst, R. L. Crawford, J. L. Garland, D. A. Lipson, A. L. Mills, and L. D. Stetzenbach (Washington, DC: ASM Press).
Tebo, B. M., Johnson, H. A., McCarthy, J. K., and Templeton, A. S. (2005). Geomicrobiology of manganese(II) oxidation. Trends Microbiol. 13, 421–428. doi: 10.1016/j.tim.2005.07.009
Trouwborst, R. E., Clement, B. G., Tebo, B. M., Glazer, B. T., and Luther, G. W. (2006). Soluble Mn(III) in suboxic zones. Science 313, 1955–1957. doi: 10.1126/science.1132876
van Waasbergen, L. G., Hildebrand, M., and Tebo, B. M. (1996). Identification and characterization of a gene cluster involved in manganese oxidation by spores of the marine Bacillus sp. strain SG-1. J. Bacteriol. 178, 3517–3530. doi: 10.1128/jb.178.12.3517-3530.1996
Webb, S. M., Dick, G. J., Bargar, J. R., and Tebo, B. M. (2005). Evidence for the presence of Mn(III) intermediates in the bacterial oxidation of Mn(II). Proc. Natl. Acad. Sci. U.S.A. 102, 5558–5563. doi: 10.1073/pnas.0409119102
Keywords: Manganese(III), Mn(III)-DFOB, Mn(III)-citrate, Mn(III)-L, Pseudomonas, bacterial manganese oxidation
Citation: Wright MH, Geszvain K, Oldham VE, Luther GW III and Tebo BM (2018) Oxidative Formation and Removal of Complexed Mn(III) by Pseudomonas Species. Front. Microbiol. 9:560. doi: 10.3389/fmicb.2018.00560
Received: 18 January 2018; Accepted: 12 March 2018;
Published: 12 April 2018.
Edited by:
Christopher Rensing, Fujian Agriculture and Forestry University, ChinaReviewed by:
Dimitry Y. Sorokin, Winogradsky Institute of Microbiology (RAS), RussiaEric D. van Hullebusch, IHE Delft Institute for Water Education, Netherlands
Copyright © 2018 Wright, Geszvain, Oldham, Luther and Tebo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mitchell H. Wright, wrigmit@ohsu.edu
Bradley M. Tebo, tebob@ohsu.edu