Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings


1
Key Laboratory of Silviculture and Conservation, Ministry of Education, Beijing Forestry University, Beijing 100083, China
2
East China Inventory and Planning Institute of National Forestry and Grassland Administration, Hangzhou 310019, China
*
Author to whom correspondence should be addressed.
Forests 2019, 10(11), 1035; https://doi.org/10.3390/f10111035
Submission received: 16 August 2019
/
Revised: 8 October 2019
/
Accepted: 12 November 2019
/
Published: 15 November 2019
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:Plant growth depends on soil mineral elements, a lack of which results in reduced nutrient accumulation leading to poor growth and resistance in plants. Therefore, more information is needed about the response of Pistacia chinensis Bunge (P. chinensis) seedlings to nutrient deficiency. In this study, we investigated how soil nutrient availability affects the nutrient accumulation and root system of P. chinensis seedlings. Seedlings were cultivated under five different nutrient treatments (500 mg, 400 mg, 300 mg, 200 mg, and 100 mg N). Various indices, including seedling growth, nutrient accumulation and root morphology, were analyzed at the end of the growing season. Nutrient deficiency (300 mg, 200 mg, and 100 mg N) reduced the accumulation of nitrogen (N), phosphorus (P) and potassium (K) in roots and stems, while the nutrient proportion of N, P, and K stored in the roots and root to shoot ratio (R/S) was increased at the end of growing season. Root length, root surface area, and root volume of very fine roots (<0.5 mm in diameter) and coarse roots (>3.0 mm in diameter) of the three lower nutrient treatments were significantly lower than those of the two highest nutrient treatments, while no significant difference was detected in the fine roots (1.0–3.0 mm in diameter). Instead, foliar N and K contents in seedlings treated with the two highest treatments were significantly greater than those of the three lower nutrient treatments, resulting in a greater nutrient loss ratio. However, seedlings treated with 100 mg N had significantly higher foliar P content than those treated with 500 mg. Seedlings treated with 300 mg and 200 mg N did not have restricted root nutrient accumulation but did have reduced nutrient accumulation in the stems. The 100 mg N treatment significantly reduced the root nutrient accumulation of N and K. The 500 mg N treatment did not increase the accumulation of nutrients in the storage organs compared with the 400 mg N treatment, but did increase the loss of N and K due to defoliation in autumn. In conclusion, there is a threshold for nutrient accumulation in storage organs at the nursery stage under a specific environment. P. chinensis seedlings reduced the negative effects of nutrient deficiency by promoting root growth, particularly fine roots, and increasing N and K allocation in storage organs.
1. Introduction
Forest regeneration depends largely on soil nutrient availability. Mineral elements, such as plant nutrients, play essential roles in plant processes and life activities such as protein synthesis, cell turgor regulation, charge balance maintenance, and signal transduction. Deciduous plants accumulate large amounts of nutrients, including nitrogen (N), phosphorus (P), and potassium (K), which are stored in the roots and stems at the end of the growing season [1]. High nutrient accumulation can increase seedling stress resistance. For instance, K nutrient enrichment around the cell protoplast can improve the drought resistance of hibiscus [2]. When soil nutrients cannot meet plant nutrient demand, the stored nutrients are the primary nutrient source for new growth [3]. High nutrient storage in water-stressed Quercus variabilis seedlings had similar dry mass compared to lower nutrient storage but well-watered seedlings, which indicates that high nutrient storage partially counteracted the negative effects of moderate water stress on seedling growth [4]. Moreover, nutrition loading can promote bud burst and root growth in Norway spruce seedlings [5].
Root morphology is a complex system consisting of roots of different diameters and structures [6,7]. Generally, fine roots, defined as roots <2 mm in diameter, have strong water and nutrient absorption capacity and are considered as the most physiologically active underground organs [8,9,10]. Volder et al. [11] reported that the new fine roots of grapes had higher nitrate uptake capacities than older lateral roots. Moreover, the morphological characteristics of fine roots may affect the root respiratory function of Quercus serrata [12]. Coarse roots are also important as they increase the water and nutrient transport capacity to aboveground organs [13]. Coarse roots are central repositories for starch and sugar in the late season [14]. Therefore, the root system morphology reflects both the nutrient uptake capacity and growth potential of seedlings [15].
Root morphology varies based on external resources such as soil composition [16,17,18]. For example, plants distribute a high proportion of biomass to the roots in a nutrient-deficient state [19]. Despite that, nutrient deficiencies and toxicities can reduce overall root growth and alter root morphology [13,20,21,22,23]. Wan et al. [23] showed that high fertilization reduced 0.2–0.5 mm in diameter root lengths of larch seedlings. In Chinese pine seedlings, a reduction of available N in the soil increased the number and length of fine roots and decreased the stele diameter and tracheid diameter of coarse roots [13]. In addition, plant ectomycorrhizal roots exude organic anions to enhance the mobilization of Mg, P, and K from mineral particles under nutrient-deficient conditions, such as Pinus sylvestris Linn [24]. Chlorophyll concentration in the leaves decreased due to N deficiency [25,26], which led to a decrease in photosynthetic efficiency [27]. Shi et al. [28] reported that a lack of K would limit the flexibility of photosynthate transfer from aboveground to belowground parts during early growth of Pinus koraiensis Sieb. et Zucc. seedlings. The sensitivity of these physiological processes to nutrient deficiency may alter the root morphology, thus reducing nutrient accumulation and changing the nutrient allocation in plants.
Pistacia chinensis Bunge (P. chinensis), a broadleaf deciduous tree with ecological and energy-related applications, was selected for this study [29,30,31,32,33]. Trubat et al. reported that nitrogen deficiency resulted in a decrease of aboveground biomass accumulation but did not affect belowground biomass accumulation or root morphology [34]. Therefore, the subdivision of roots according to root diameter may provide more information for understanding the response of root morphology to nutrient deficiency. However, little is known about the effects of nutrient deficiency on root morphology and nutrient allocation in P. chinensis seedlings. We tested the following hypotheses: (1) nutrient deficiency promotes fine root growth and (2) increases nutrient relocation from leaves to stems or roots, resulting in high nutrient allocation in storage organs. To test these hypotheses, we cultivated seedlings under a variety of nutrient treatments and recorded various indices, including seedling growth, nutrition accumulation and root morphology, after one growing season.
2. Materials and Methods
2.1. Plant Material and Experimental Design
Mature P. chinensis seeds were collected from one open-pollinated mother tree in early September 2016 in Linzhou, Henan, China (36°14′N, 113°57′E). The seeds were manually stripped of their pericarps, placed in a cotton fabric bag, and stored at 2 °C and 60% humidity until the following April.
After six months of storage, seeds of similar size were selected for the study. The average seed length, width, and height were 5.52 ± 0.23 mm, 5.12 ± 0.22 mm, and 4.13 ± 0.18 mm, respectively. Seeds were germinated on a moist sand bed covered with gauze until about 30% of the seed coats had broken on 23 April 2017. Then, one seed per container was sown in cylindrical hard plastic 983 mL containers (D60, Stuewe & Sons, Inc., Tangent, Oregon, USA; diameter and depth of 6.4 cm and 36.0 cm, respectively) at a depth of 0.5–1 cm and grown in a greenhouse in the Beijing Dadongliu Nursery (40°10′N, 116°28′E). The container was filled with a mix of 70% peat (Pindstrup seeding, Ryomgaard, Denmark; pH = 6.0; screening, 0–6 mm) and 30% perlite (5 mm diameter; Xinyang Jinhualan Mining Co., Henan, China).
Osmocote NO.5 (14N-5.7P-10.8K; Scotts®, Marysville, OH, USA) 5–6 month controlled-release fertilizer (CRF) was incorporated into the growing media at one of five rates: 500 mg, 400 mg, 300 mg, 200 mg and 100 mg N per seedling (Table 1). A total of 300 germinating seeds were completely randomized into 30 trays (10 containers per tray) and randomly distributed into five nutrient treatments. Containers were rotated positions once a week to eliminate edge effects. The day/night temperatures in the greenhouse were approximately 28/16 °C. All of the seedlings were irrigated from overhead when the gravimetric water content (GWC) reached 70% of field capacity. In the middle of October, seedlings were moved outside to accelerate hardening.
2.2. Measurement of Seedling Morphology and Tissue Nutrient Concentration
Seedlings were covered with white nylon mesh bags to collect fallen leaves on 4 November 2017. Nine seedlings per treatment were randomly sampled for morphology and nutrient analyses once all leaves had abscised (29 November). Roots were washed gently to remove the growth medium, and each plant was excised at the root collar and separated into roots and stems. Seedling height was measured as the length of the stem (from the cotyledon insertion point to the tip of the apical bud), and root-collar diameter (RCD) was measured slightly above (2 mm) the cotyledon insertion point. Root system images were acquired by carefully scanning with an Epson Twain Pro high-quality scanner at 400 dpi (Epson (China) Co., Ltd., Beijing, China) and the root systems based on the images were divided into six classes according to root diameter: 0–0.2 mm (class 1), 0.2–0.5 mm (class 2), 0.5–1.0 mm (class 3), 1.0–2.0 mm (class 4), 2.0–3.0 mm (class 5), and >3.0 mm (class 6). Scanned images of the roots were used to determine root length, root surface area, and root volume using WinRHIZO software (Régent Instruments Inc., Quebec, Canada). Then, the stems, leaves, and roots were oven-dried at 70 °C for 48 h to determine their dry mass. Each of the individual organ samples were subsequently ground, sieved through a 0.25 mm screen, and wet-digested using the H2SO4–H2O2 method [35]. An AutoAnalyzer III (Bran Luebbe GmbH, Germany) was used to measure total N. P was determined with a UV-visible spectrophotometer (Agilent 8453, Palo Alto, USA) and K was quantified using atomic emission photometry (SpectrAA 220 Atomic Absorption Spectrometer, VARIN, Palo Alto, USA).
2.3. Data Calculations
The nutrient (N, P, or K) content increment (Ri) between two nutrient treatments (i) was calculated using Equation (1):
where Xi is the nutrient content of the roots, stems, or leaves at nutrient treatment i. The nutrient content proportion of roots (PRj) and stems (PSj) to storage organs was calculated using Equations (2) and (3), respectively:
where X(root)j and X(stem)j are the mean nutrient content of the roots and stems at nutrient treatment j, respectively. The proportion of nutrient storage (PSSj) and loss (PLLj) to plant uptake was calculated using Equations (4) and (5), respectively:
where X(storage)j is the mean of the sum of the root and stem nutrient contents at nutrient treatment j, and X(leaf)j is the mean of the nutrient content of the leaves at nutrient treatment j.
Ri = X(i+100) − Xi (mg)
i = 400, 300, 200, 100
j = 500, 400, 300, 200, 100
2.4. Statistical Analyses
Single-factor analysis of variance (ANOVA) was used to analyze the effects of nutrient treatment using SPSS 22.0 for Windows (SPSS Statistics, Inc., Chicago, IL, USA). Multiple comparisons of means were analyzed using Duncan’s multiple range test at α = 0.05. Normality and variance homogeneity requirements were met, and no data transformation was necessary. Graphs were produced using SigmaPlot 14.0 (Systat Software, Inc., San Jose, CA, USA).
3. Results
3.1. Seedling Morphology
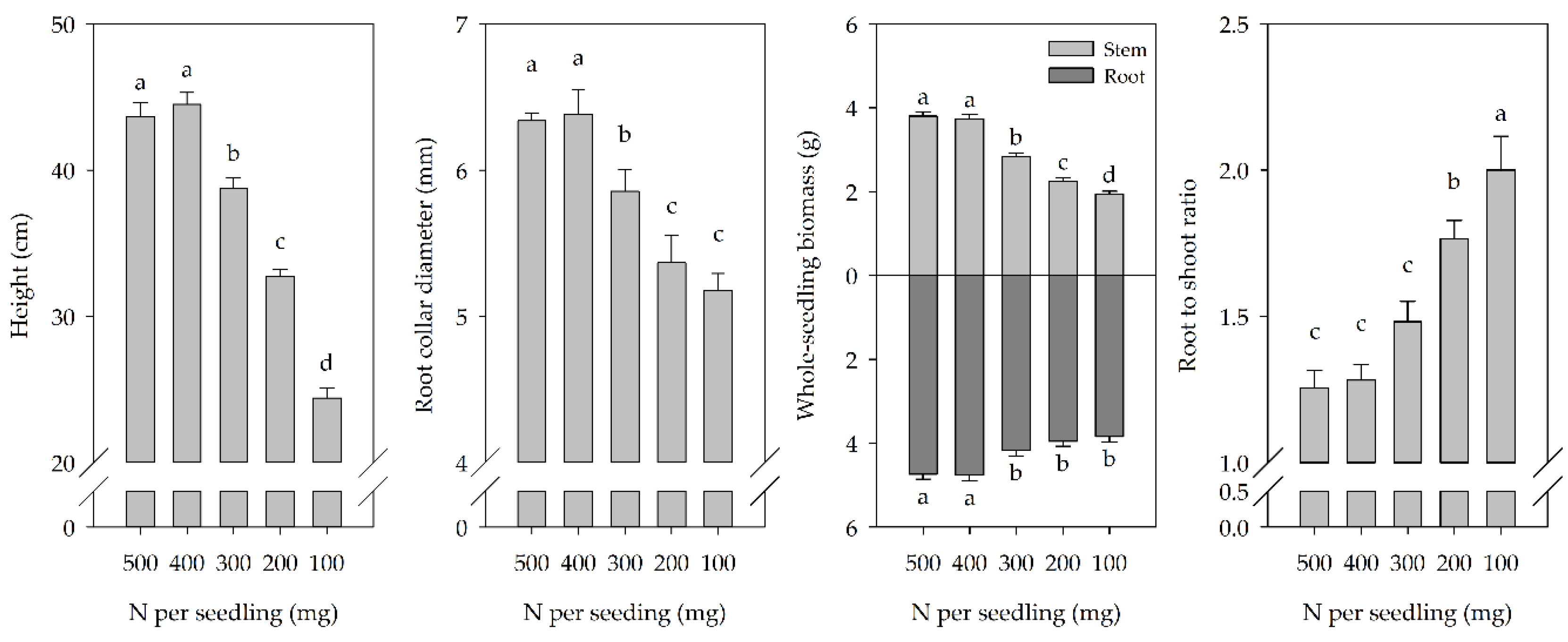
Seedling height, RCD, and whole-seedling biomass decreased, but R/S increased with decreasing nutrient treatment (Figure 1), though there was no difference between the 500 mg and the 400 mg treatments. The height, RCD, and whole-seedling biomass of the 100 mg N treatment were 45.1%, 18.9%, and 32.1% lower relative to the 400 mg N treatment, respectively. However, the R/S treated with 100 mg N treatment was significantly higher than that of other nutrient treatments.
3.2. Nutrient Concentration of the Storage Organs
There was no significant difference for the N, P, and K concentrations of roots and stems among the 500 mg, 400 mg, 300 mg, and 200 mg N treatments, while the 100 mg N treatment had significantly lower nitrogen concentration of roots and stems than other treatments. Seedlings treated with 100 mg N had significantly lower P concentration of roots than seedlings treated with 500 mg, 400 mg, and 300 mg N treatments. The K concentration of stems in the 100 mg N treatment was significantly lower than that in other treatments (Table 2).
3.3. Nutrient Accumulation of the Storage Organs
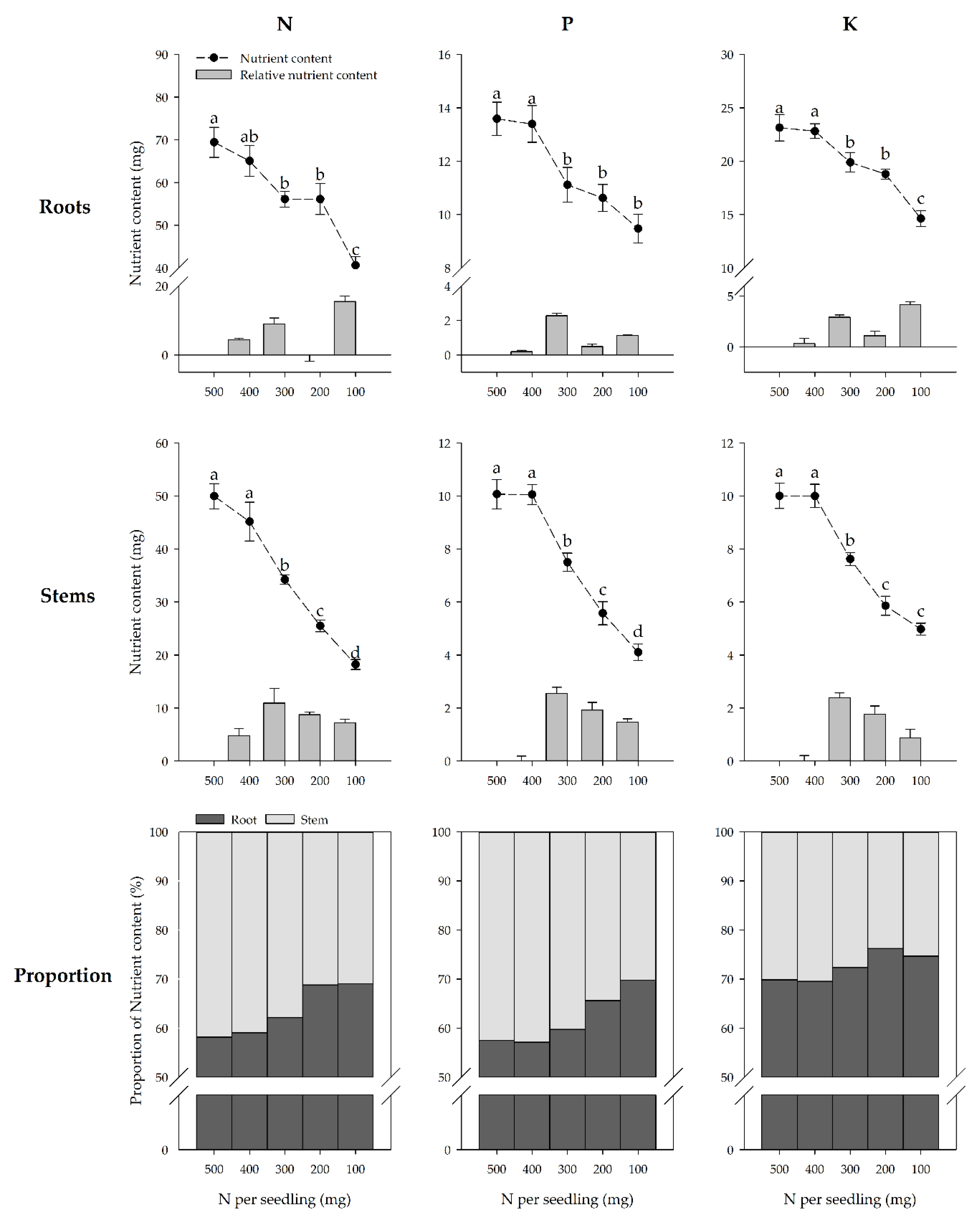
The nutrient content of the roots and stems decreased with decreasing nutrient treatments (Figure 2). No impact on N, P, and K content in roots and stems was observed for the seedlings in the two highest nutrient treatments. However, the reduction in N, P, and K content of roots was accelerated in 300–100 mg treatment groups. In contrast to the roots, the decrease for N, P, and K contents of stems slowed down with the decrease of nutrient treatments.
The proportion of N, P, and K in the roots exceeded 50% under all nutrient treatments at the end of the growing season (Figure 2). The plants in the two highest nutrient treatments did not differ in their proportion of root N, P, and K content to total nutrient content. The highest proportion of N, P, and K in the roots occurred in the lowest nutrient treatments.
3.4. Nutrient Loss of the Seedlings
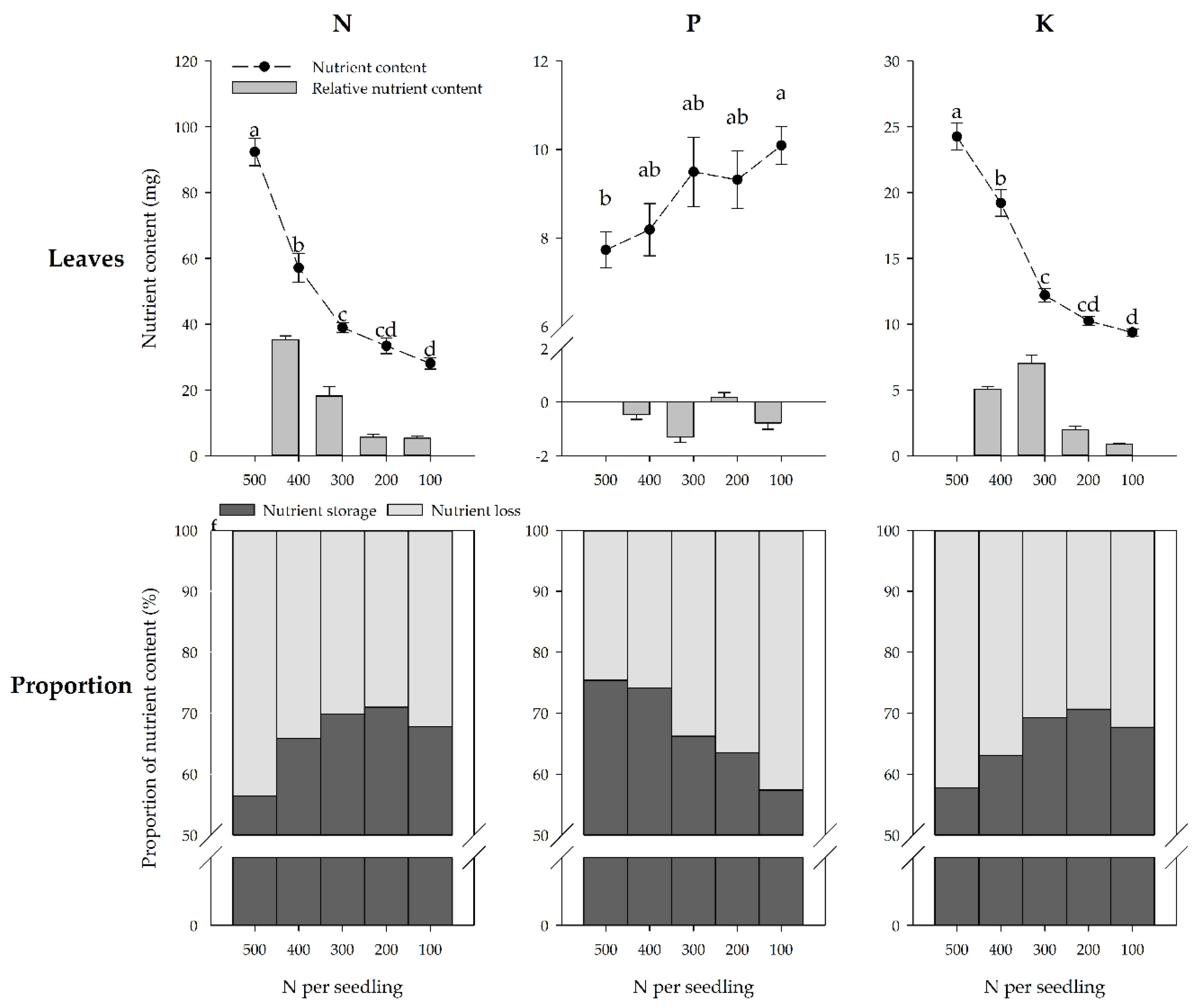
The roots and stems are the nutrient storage organs in deciduous plants, as the nutrients in the leaves are lost as they fall off in autumn. Leaf N and K contents decreased, but foliar P content increased with the decrease of nutrient treatments (Figure 3). The foliar N and K content were 61.7% and 26.3% higher respectively for the seedlings in the 500 mg treatment than that in the 400 mg treatment. Afterward, the N and K content decreased slowly in the 300–100 mg treatment groups. The seedlings in the 100 mg treatment group had significantly higher leaf P contents than those in the 500 mg treatment group.
The proportion of nutrient loss corresponded with nutrient treatments and varied with the different nutrients (Figure 3). The N and K loss proportion decreased, but the P loss proportion increased with the decrease in nutrient treatments. Remarkably, the three lower nutrient treatment seedlings had lower N and K losses but higher P losses compared to the highest nutrient treatment.
3.5. Root Morphology
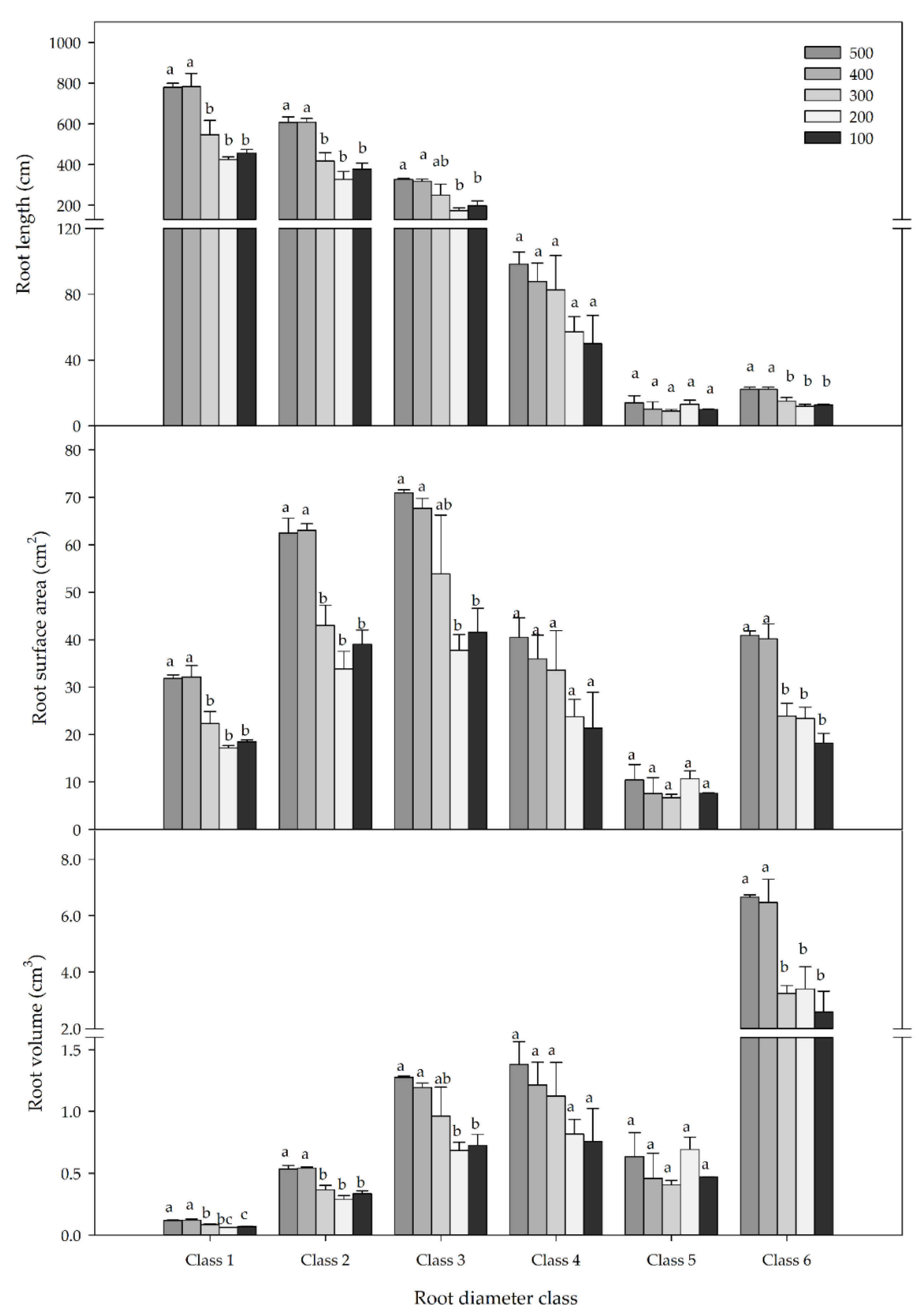
The root morphology of the P. chinensis seedlings was significantly affected by nutrient treatment, root diameter, and their interaction (Table 3). The root length, root surface area, and root volume were obviously higher at specific root diameter classes (Figure 4). The root lengths of classes 1 and 2 (accounting for 42.1 and 32.9% of the total root length, respectively) were higher than the rest of the root diameter classes. Roots in diameter classes 1, 2, and 3 had higher root surface areas (accounting for 12.5%, 24.6%, and 27.7% of the total root surface area, respectively) compared to the remainder of the diameter class. Furthermore, the root volume of class 6 accounted for 59.3% of the total root volume, which exceeded the total root volume of the other diameter classes.
The two highest nutrient treatments had a significantly higher root length, root surface area, and root volume in classes 1, 2 and 6 compared to the other three treatments. Nevertheless, there was no significant difference in root length, root surface area, or root volume of the seedlings under different nutrient treatments for classes 4 and 5. In addition, there was no significant difference in root morphology among the three lower nutrient treatments (Figure 4).
4. Discussion
In this study, the morphological index and nutrient content of seedlings treated with the two highest nutrient treatments were significantly higher than those treated with the three lower nutrient treatments. At the two highest nutrient treatments, fertility did not significantly increase the accumulation of N, P, and K contents in the roots and stems. Therefore, we considered that the seedlings were nutrient-sufficient when exposed to 500 mg and 400 mg N treatments, but were nutrient-deficient when exposed to 300 mg, 200 mg, and 100 mg N treatments. Additionally, the proportions of N, P, and K contents in the roots or stems to storage organs did not differ significantly at the two highest nutrient treatments, which indicated that higher nutrient treatment did not alter the nutrient allocation in the storage organs under sufficient nutrient conditions. As noted in previous studies in Picea mariana, nutrient uptake in seedlings increased with the increase of soil fertility under specific environmental conditions, but excessive nutrient supply would not continue to promote seedling growth [36]. This indicates that there is a threshold for the nutrient accumulation of storage organs during the growth period of seedlings. In our study, foliar N and K contents increased significantly when fertilizer application increased under sufficient nutrient conditions. This result is correlated to the increase of transpiration by fertilization [37], which may enhance the mass-flow of nutrients to leaves. The accumulation of foliar N and K decreased with defoliation, resulting in high N and K loss rates under higher nutrient treatment. A similar result was reported in Salix gordejevii Chang [38].
Almost all of the mineral nutrients in seedlings are obtained from the soil via the root system [39,40]. In this study, nutrient treatment and root diameter and their interaction influenced the root length, root surface area, and root volume of the seedlings. Seventy-five percent of the total root length was very fine roots (<0.5 mm in diameter, classes 1 and 2), which is considered as a better indicator of root response to soil nutrients [13]. Roots with a diameter of less than 1 mm (classes 1–3) accounted for 64.8% of the total root surface area. Coarse roots (>3.0 mm in diameter, class 6) accounted for 59.3% of the total root volume. Previous studies reported that fine roots affect water and nutrient uptake, while coarse roots determine water and nutrient transport capacity [8,13]. The very fine root length, surface area with a root diameter less than 1 mm, and coarse root volume at the two highest treatments were significantly higher than those at the three lower nutrient treatment, suggesting root growth directly promoted the nutrient uptake and transport of seedlings treated with the higher nutrient treatment. In contrast, the R/S treated with the 200 mg and 100 mg N treatment was significantly higher than that of the two highest nutrient treatments, indicating that nutrient deficiency promoted root growth.
Mineral nutrients in many deciduous species withdraw from the leaves to the branches and roots before autumn defoliation to reduce nutrient loss, as observed in Salix dasyclados Wimm [41], Populus nigra Linn. [42], birch, and larch [43]. For the current study, the proportion of N and K lost by defoliation was lower, and the proportion of N, P, and K stored in the roots was higher in the seedlings treated with the three lower nutrient treatments than in the seedlings treated with the two highest nutrient treatments, indicating that nutrient deficiency promoted nutrient transfer from the leaves and stems to the roots. In addition, there were no significant differences in root length, root surface area, and root volume between root diameters larger than 1 mm and less than 3 mm (classes 4 and 5) treated with the different nutrient treatment, indicating that nutrient deficiency encouraged the seedlings to use more nutrients for the growth of fine roots (1.0–3.0 mm in diameter). Keyes and Grier [44] reported that for Douglas fir aboveground productivity, the seedlings might need greater investment in fine roots on harsher sites. Previous studies have shown that N is transported in the phloem as a mixture of amino acids and amides [45], eventually reaching the root or stem phloem cells where it is stored in cytoplasmic vacuoles in the form of storage proteins in deciduous trees [46,47]. Therefore, fine roots are not only a link between very fine roots and coarse roots, but are also an important component of nutrient storage in the roots.
In current study, the root morphology of P. chinensis seedlings among the three lower nutrient treatments did not differ significantly, which is consistent with the result that root N and K contents between the 300 mg and 200 mg N treatments did not differ significantly. Conversely, N and K contents in roots of seedlings treated with the 100 mg N treatment were significantly lower than those treated with the 200 mg N treatment. Meanwhile, the proportion of N and K defoliation lost under 100 mg N treatment was not less than that under the 200 mg N treatment, and the proportion of N and K stored in the roots under 100 mg N treatment was not higher than that under the 200 mg N treatment, which suggested that the nutrient allocation in stems and roots is limited by the degree of nutrient deficiency. Two reasons may explain the above results. First, root growth counteracted some of the negative effects of nutrient deficiency, and the factor limiting nutrient uptake in the seedlings under higher nutrient deficiency was no longer root absorptivity, but rather soil ion concentration [39]. Second, the total photosynthate of the leaves decreased continuously with the increase in nutrient deficiency [48,49]. The nutrients transported from the aboveground to the belowground organs decrease when the proportion of nutrients transferred from the leaves to the roots or stems reaches the threshold value [28,50].
Surprisingly, the variation in foliar P accumulation with nutrient treatment differed from that of N and K, and this phenomenon was not observed in the roots and stems. The accumulation of foliar P increased slowly with the decrease in nutrient treatment, which indicated that nutrient deficiency might limit P transport in the leaves. This result is inconsistent with previous studies where the foliar P concentration of the high-fertility plants was higher than that of the low-fertility plants before abscission in S. dasyclados [41]. P is stored in organic and inorganic forms in plants [51,52,53]. Chapin et al. [43] reported that the quantities of nucleic acids and phospholipids that were hydrolyzed, which is an energy-consuming process, in autumn were equivalent to 40–47% and 26–38%, respectively, of the total P re-translocated from the leaves of deciduous species before abscission. P deficiency increases the fraction of nucleotides and phospholipids to total phosphorus in tobacco leaves [54]. Nucleotides and phospholipids of leaves in seedlings treated with nutrient deficiency might not be fully hydrolyzed, resulting in reduced P transport before defoliation. However, this inference should be further validated by the determination of nucleotide and phospholipid content in leaves.
The results of the present study have ecological and practical implications for forest restoration plantations. Stems and roots act as nutrient sink organs at the end of seedling growth in deciduous species, thus determining the growth status of seedlings in the next growing season [4]. As the nutrient content of P. chinensis seeds is low, the nutrients needed for seed germination are highly dependent on external nutrients. Seedlings have developed two strategies to reduce the inhibition of nutrient deficiency. First, higher proportions of nutrients and photosynthates are used for root growth to increase nutrient uptake in growing seedlings. Second, seedlings increase nutrient accumulation by increasing transport efficiency from the leaves to the roots or stems at the end of the growing season. However, the efficiency of these two strategies would decrease with the increase in nutrient deficiency. Our data also show that increasing fertilizer under nutrient sufficiency did not promote nutrient accumulation in the storage organs, and most of the nutrients were lost with defoliation in autumn. Hence, our findings could help to cultivate high-quality seedlings with well-developed roots by reducing the negative effects of nutrient deficiency on seedling growth and also avoiding excessive fertilization when seedlings reach nutrient sufficiency.
5. Conclusions
Compared with the two highest nutrient treatments, the three lower nutrient treatments had higher R/S and the proportion of N and K in storage organs, and lower proportions of N and K lost by defoliation. This indicated that nutrient deficiency promoted root growth and increased N and K allocation in storage organs. There were no significant differences in root length, root surface area, or root volume of fine roots treated with the different nutrient treatments, indicating that nutrient deficiency mainly promoted the growth of fine roots. The 100 mg N treatment increased the proportion of N and K lost by defoliation compared with the 200 mg N treatment, which suggested that the ability of nutrient deficiency to promote nutrient relocation from leaves to stems and roots was affected by the degree of nutrient deficiency. Also, improving fertilizer under sufficient nutrient conditions did not promote nutrient accumulation in the storage organs, and most of the nutrients were lost with defoliation.
Author Contributions
Conceptualization and methodology, Y.L. and X.S.; software, X.S., X.C. and J.Z.; formal analysis, X.S.; investigation, X.S., F.W., X.C., J.Z. and M.S.; writing—original draft, X.S.; writing—review and editing, Y.L. and F.W.; funding acquisition, Y.L.
Funding
This research was funded by the Project of Beijing Gardening and Greening Bureau, grant number CEG-2015-01.
Acknowledgments
We gratefully acknowledge the editors and reviewers and wish to thank the managers and workers of Beijing Dadongliu nursery for their valuable assistance with this project. We thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Taiz, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates: Sunderland, UK, 2006. [Google Scholar]
- Egilla, J.N.; Davies, F.T.; Drew, M.C. Effect of potassium on drought resistance of Hibiscus rosa-sinensis cv. Leprechaun: Plant growth, leaf macro- and micronutrient content and root longevity. Plant Soil 2001, 229, 213–224. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Uscola, M.; Jacobs, D.F. The role of stored carbohydrates and nitrogen in the growth and stress tolerance of planted forest trees. New For. 2015, 46, 813–839. [Google Scholar] [CrossRef]
- Wang, J.; Villar-Salvador, P.; Li, G.; Liu, Y. Moderate water stress does not inhibit nitrogen remobilization, allowing fast growth in high nitrogen content Quercus variabilis seedlings under dry conditions. Tree Physiol. 2018, 39, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Luoranen, J.; Rikala, R. Nutrient loading of Norway spruce seedlings hastens bud burst and enhances root growth after outplanting. Silva Fenn. 2011, 45, 319–329. [Google Scholar] [CrossRef]
- Adams, T.S.; Mccormack, M.L.; Eissenstat, D.M. Foraging strategies in trees of different root morphology: The role of root lifespan. Tree Physiol. 2013, 33, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Goebel, M.; Hobbie, S.E.; Bulaj, B.; Zadworny, M.; Archibald, D.D.; Oleksyn, J.; Reich, P.B.; Eissenstat, D.M. Decomposition of the finest root branching orders: Linking belowground dynamics to fine-root function and structure. Ecol. Monogr. 2011, 81, 89–102. [Google Scholar] [CrossRef]
- Doi, R.; Tanikawa, T.; Miyatani, K.; Hirano, Y. Intraspecific variation in morphological traits of root branch orders in Chamaecyparis obtusa. Plant Soil 2017, 416, 503–513. [Google Scholar] [CrossRef]
- Hishi, T.; Tateno, R.; Takeda, H. Anatomical characteristics of individual roots within the fine-root architecture of Chamaecyparis obtusa (Sieb. & Zucc.) in organic and mineral soil layers. Ecol. Res. 2006, 21, 754–758. [Google Scholar]
- Makita, N.; Hirano, Y.; Mizoguchi, T.; Kominami, Y.; Dannoura, M.; Ishii, H.; Finér, L.; Kanazawa, Y. Very fine roots respond to soil depth: Biomass allocation, morphology, and physiology in a broad-leaved temperate forest. Ecol. Res. 2011, 26, 95–104. [Google Scholar] [CrossRef]
- Volder, A.; Smart, D.R.; Bloom, A.J.; Eissenstat, D.M. Rapid decline in nitrate uptake and respiration with age in fine lateral roots of grape: Implications for root efficiency and competitive effectiveness. New Phytol. 2005, 165, 493–502. [Google Scholar] [CrossRef]
- Makita, N.; Hirano, Y.; Dannoura, M.; Kominami, Y.; Mizoguchi, T.; Ishii, H.; Kanazawa, Y. Fine root morphological traits determine variation in root respiration of Quercus serrata. Tree Physiol. 2009, 29, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, F.; Xue, S. Nitrogen addition enhanced water uptake by affecting fine root morphology and coarse root anatomy of Chinese pine seedlings. Plant Soil 2017, 418, 177–189. [Google Scholar] [CrossRef]
- Nguyen, P.V.; Dickmann, D.I.; Pregitzer, K.S.; Hendrick, R. Late-season changes in allocation of starch and sugar to shoots, coarse roots, and fine roots in two hybrid poplar clones. Tree Physiol. 1990, 7, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.S.; Jacobs, D.F. Quantifying root system quality of nursery seedlings and relationship to outplanting performance. New For. 2005, 30, 295–311. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L. Seedling root morphology and biomass allocation of 62 tropical tree species in relation to drought- and shade-tolerance. J. Ecol. 2009, 97, 311–325. [Google Scholar] [CrossRef]
- Mcinenly, L.E.; Merrill, E.; Cahill, J.F.; Juma, N.G. Festuca campestris alters root morphology and growth in response to simulated grazing and nitrogen form: Defoliation, N-form and fescue roots. Funct. Ecol. 2010, 24, 283–292. [Google Scholar] [CrossRef]
- Zangaro, W.; Nishidate, F.R.; Camargo, F.R.S.; Romagnoli, G.G.; Vandressen, J. Relationships among Arbuscular Mycorrhizas, Root morphology and seedling growth of tropical native woody species in southern Brazil. J. Trop. Ecol. 2005, 21, 529–540. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Cao, X.; Chen, C.; Zhang, D.; Shu, B.; Xiao, J.; Xia, R. Influence of nutrient deficiency on root architecture and root hair morphology of trifoliate orange (Poncirus trifoliata L. Raf.) seedlings under sand culture. Sci. Hortic. 2013, 162, 100–105. [Google Scholar] [CrossRef]
- Gao, Y.; Li, X.; Tian, Q.; Wang, B.; Zhang, B. Sulfur deficiency had different effects on Medicago truncatula ecotypes A17 and R108 in terms of growth, root morphology and nutrient contents. J. Plant Nutr. 2016, 39, 301–314. [Google Scholar] [CrossRef]
- Mollier, A.; Pellerin, S. Maize root system growth and development as influenced by phosphorus deficiency. J. Exp. Bot. 1999, 50, 487–497. [Google Scholar] [CrossRef]
- Wan, F.; Ross-Davis, A.L.; Shi, W.; Weston, C.; Song, X.; Chang, X.; Davis, A.S.; Liu, Y.; Teng, F. Subirrigation effects on larch seedling Growth, root morphology, and media chemistry. Forests 2019, 10, 38. [Google Scholar] [CrossRef]
- Schöll, L.V.; Hoffland, E.; Breemen, N.V. Organic acid exudation by ectomycorrhizal fungi and Pinus sylvestris in response to nutrient deficiencies. New Phytol. 2006, 170, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.W.; Dale, J.E. Nitrogen deficiency and fertilization effects on needle growth and photosynthesis in Sitka spruce (Picea sitchensis). Tree Physiol. 1995, 15, 813–817. [Google Scholar] [CrossRef]
- Ciompi, S.; Gentili, E.; Guidi, L.; Soldatini, G.F. The effect of nitrogen deficiency on leaf gas exchange and chlorophyll fluorescence parameters in sunflower. Plant Sci. 1996, 118, 177–184. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Shi, Z.; Balogh-Brunstad, Z.; Grant, M.; Harsh, J.; Gill, R.; Thomashow, L.; Dohnalkova, A.; Stacks, D.; Letourneau, M.; Keller, C.K. Cation uptake and allocation by red pine seedlings under cation-nutrient stress in a column growth experiment. Plant Soil 2014, 378, 83–98. [Google Scholar] [CrossRef]
- Lu, L.; Jiang, D.; Fu, J.; Zhuang, D.; Huang, Y.; Hao, M. Evaluating energy benefit of Pistacia chinensis based biodiesel in China. Renew. Sustain. Energy Rev. 2014, 35, 258–264. [Google Scholar] [CrossRef]
- Zhu, B.; Wang, Q.; Roge, E.F.; Nan, P.; Liu, Z.; Zhong, Y. Chemical variation in leaf oils of Pistacia chinensis from five locations in China. Chem. Nat. Compd. 2006, 42, 422–425. [Google Scholar] [CrossRef]
- Li, X.; He, X.; Li, Z.; Wang, Y.; Wang, C.; Shi, H.; Wang, F. Enzymatic production of biodiesel from Pistacia chinensis bge seed oil using immobilized lipase. Fuel 2012, 92, 89–93. [Google Scholar] [CrossRef]
- Noureen, F.; Khan, M.R.; Shah, N.A.; Khan, R.A.; Naz, K.; Sattar, S. Pistacia chinensis: Strong antioxidant and potent testicular toxicity amelioration agent. Asian Pac. J. Trop. Med. 2017, 10, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Yayeh, T.; Hong, M.; Jia, Q.; Lee, Y.C.; Kim, H.J.; Hyun, K.T.; Rhee, M.H. Pistacia chinensis inhibits NO production and upregulates HO-1 induction via PI-3K/Akt pathway in LPS stimulated macrophage cells. Am. J. Chin. Med. 2012, 40, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Plant morphology and root hydraulics are altered by nutrient deficiency in Pistacia lentiscus (L.). Trees 2006, 20, 334–339. [Google Scholar] [CrossRef]
- Lowther, J.R. Use of a single sulphuric acid-hydrogen peroxide digest for the analysis of Pinus radiata needles. Commun. Soil Sci. Plant Anal. 1980, 11, 175–188. [Google Scholar] [CrossRef]
- Salifu, K.F.; Timmer, V.R. Optimizing nitrogen loading of Picea mariana seedlings during nursery culture. Can. J. For. Res. 2003, 33, 1287–1294. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Peñuelas, J.L.; Jacobs, D.F. Nitrogen nutrition and drought hardening exert opposite effects on the stress tolerance of Pinus pinea L. seedlings. Tree Physiol. 2013, 33, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.Y.; Li, L.H.; Han, X.G.; Huang, J.H.; Wan, S.Q. Foliar nitrogen dynamics and nitrogen resorption of a sandy shrub Salix gordejevii in northern China. Plant Soil 2005, 278, 183–193. [Google Scholar] [CrossRef]
- Pallardy, S.G. Physiology of Woody Plants, 3rd ed.; Academic Press: Burlington, NJ, USA, 2008. [Google Scholar]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Fircks, Y.V.; Ericsson, T.; Sennerby-Forsse, L. Seasonal variation of macronutrients in leaves, stems and roots of Salix dasyclados Wimm. grown at two nutrient levels. Biomass Bioenergy 2001, 21, 321–334. [Google Scholar] [CrossRef]
- Kurdali, F. Seasonal nitrogen changes in Alnus orientalis and Populus nigra and N2 fixation by exotic alder species in Syria. Commun. Soil Sci. Plant Anal. 2000, 31, 2509–2522. [Google Scholar] [CrossRef]
- Chapin, F.S.; Kedrowski, R.A. Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees. Ecology 1983, 64, 376–391. [Google Scholar] [CrossRef]
- Keyes, M.R.; Grier, C.C. Above- and below-ground net production in 40-year-old Douglas-fir stands on low and high productivity sites. Can. J. For. Res. 1981, 11, 599–605. [Google Scholar] [CrossRef]
- Bollard, E.G. Nitrogenous compounds in plant xylem sap. Nature 1956, 178, 1189–1190. [Google Scholar] [CrossRef]
- Cooke, J.E.; Weih, M. Nitrogen storage and seasonal nitrogen cycling in Populus: Bridging molecular physiology and ecophysiology. New Phytol. 2005, 167, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wetzel, S.; Demmers, C.; Greenwood, J.S. Spherical organelles, analogous to seed protein bodies, fluctuate seasonally in parenchymatous cells of hardwoods. Can. J. Bot. 1989, 67, 3439–3445. [Google Scholar] [CrossRef]
- Brix, H. Effects of thinning and nitrogen fertilization on growth of Douglas-fir: Relative contribution of foliage quantity and efficiency (Pseudotsuga menziesii). Can. J. For. Res. 1983, 13, 167–175. [Google Scholar] [CrossRef]
- Sheriff, D.W. Gas Exchange of Field-Grown Pinus radiate—Relationships with Foliar Nutrition and Water Potential, and with Climatic Variables. Funct. Plant Biol. 1995, 22, 1015–1026. [Google Scholar] [CrossRef]
- Cakmak, I.; Hengeler, C.; Marschner, H. Changes in phloem export of sucrose in leaves in response to phosphorus, potassium and magnesium deficiency in bean plants. J. Exp. Bot. 1994, 45, 1251–1257. [Google Scholar] [CrossRef]
- Lott, J.N.A.; Ockenden, I.; Raboy, V.; Batten, G.D. Phytic acid and phosphorus in crop seeds and fruits: A global estimate. Seed Sci. Res. 2000, 10, 11–33. [Google Scholar] [CrossRef]
- Seufferheld, M.; Curzi, M.J. Recent discoveries on the roles of polyphosphates in plants. Plant Mol. Biol. Rep. 2010, 28, 549–559. [Google Scholar] [CrossRef]
- Shane, M.W.; Mccully, M.E.; Lambers, H. Tissue and cellular phosphorus storage during development of phosphorus toxicity in Hakea prostrata (Proteaceae). J. Exp. Bot. 2004, 55, 1033–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakie, T. Phosphorus fractions in tobacco plants as affected by phosphate application. Soil Sci. Plant Nutr. 1969, 15, 81–85. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Seedling height, root collar diameter (RCD), whole-seedling (stem and root) biomass, and root to shoot ratio (R/S) (mean and standard error (SE)) of P. chinensis under specific nutrient treatments at the end of the growing season. Nitrogen fertilizer rates of 500 mg, 400 mg, 300 mg, 200 mg, and 100 mg were applied per seedling. Bars with different letters differ statistically according to Duncan’s test at α = 0.05.
Figure 1.
Seedling height, root collar diameter (RCD), whole-seedling (stem and root) biomass, and root to shoot ratio (R/S) (mean and standard error (SE)) of P. chinensis under specific nutrient treatments at the end of the growing season. Nitrogen fertilizer rates of 500 mg, 400 mg, 300 mg, 200 mg, and 100 mg were applied per seedling. Bars with different letters differ statistically according to Duncan’s test at α = 0.05.

Figure 2.
Nutrient content and nutrient content increment of the P. chinensis seedling roots and stems, and the nutrient content proportion of the roots or stems to the storage organs under specific nutrient treatment conditions at the end of the growing season. The nutrient content increment represents the N, P, or K content of the roots or stems that were reduced between two contiguous nutrient treatments. The mean and SE are shown for each nutrient treatment. Error bars with different letters differ statistically according to Duncan’s test at α = 0.05.
Figure 2.
Nutrient content and nutrient content increment of the P. chinensis seedling roots and stems, and the nutrient content proportion of the roots or stems to the storage organs under specific nutrient treatment conditions at the end of the growing season. The nutrient content increment represents the N, P, or K content of the roots or stems that were reduced between two contiguous nutrient treatments. The mean and SE are shown for each nutrient treatment. Error bars with different letters differ statistically according to Duncan’s test at α = 0.05.

Figure 3.
Nutrient content and nutrient content increment of the P. chinensis seedling leaves, and the proportion of nutrient storage and loss under specific nutrient treatment conditions at the end of the growing season. The mean and SE are shown for each nutrient treatment. Error bars with different letters differ statistically according to Duncan’s test at α = 0.05.
Figure 3.
Nutrient content and nutrient content increment of the P. chinensis seedling leaves, and the proportion of nutrient storage and loss under specific nutrient treatment conditions at the end of the growing season. The mean and SE are shown for each nutrient treatment. Error bars with different letters differ statistically according to Duncan’s test at α = 0.05.

Figure 4.
Root length, root surface area, and root volume of the P. chinensis seedlings among different root diameter classes under specific nutrient treatment conditions at the final harvest. The mean and SE are shown for each nutrient treatment and root diameter class. Nutrient treatments marked with different lowercase letters differ statistically at each root diameter class according to Duncan’s test at α = 0.05. Class 1, 0–0.2 mm; class 2, 0.2–0.5 mm; class 3, 0.5–1.0 mm; class 4, 1.0–2.0 mm; class 5, 2.0–3.0 mm; class 6 > 3.0 mm.
Figure 4.
Root length, root surface area, and root volume of the P. chinensis seedlings among different root diameter classes under specific nutrient treatment conditions at the final harvest. The mean and SE are shown for each nutrient treatment and root diameter class. Nutrient treatments marked with different lowercase letters differ statistically at each root diameter class according to Duncan’s test at α = 0.05. Class 1, 0–0.2 mm; class 2, 0.2–0.5 mm; class 3, 0.5–1.0 mm; class 4, 1.0–2.0 mm; class 5, 2.0–3.0 mm; class 6 > 3.0 mm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Nutrient composition for the five nutrient treatments.
| Treatments | N (mg) | P (mg) | K (mg) |
|---|---|---|---|
| 500 | 500 | 204 | 386 |
| 400 | 400 | 163 | 309 |
| 300 | 300 | 122 | 231 |
| 200 | 200 | 81 | 154 |
| 100 | 100 | 41 | 77 |
Table 2.
Nutrient concentration of root and stem in P. chinensis under specific nutrient treatment conditions at the end of the growing season.
Table 2.
Nutrient concentration of root and stem in P. chinensis under specific nutrient treatment conditions at the end of the growing season.
| Organs | Treatments | N Concentration (mg/g) | P Concentration (mg/g) | K Concentration (mg/g) |
|---|---|---|---|---|
| 500 | 14.66 ± 0.74a | 2.87 ± 0.13a | 4.88 ± 0.26a | |
| 400 | 13.66 ± 0.76a | 2.81 ± 0.14a | 4.79 ± 0.15a | |
| Roots | 300 | 13.44 ± 0.44a | 2.66 ± 0.16a | 4.76 ± 0.22a |
| 200 | 14.20 ± 0.91a | 2.69 ± 0.13a | 4.75 ± 0.12a | |
| 100 | 10.59 ± 0.53b | 2.47 ± 0.14a | 3.81 ± 0.20b | |
| 500 | 13.15 ± 0.62a | 2.65 ± 0.14a | 2.63 ± 0.13a | |
| 400 | 12.10 ± 0.98a | 2.69 ± 0.10a | 2.68 ± 0.12a | |
| Stems | 300 | 12.08 ± 0.32a | 2.65 ± 0.12a | 2.69 ± 0.09a |
| 200 | 11.34 ± 0.49a | 2.48 ± 0.19ab | 2.60 ± 0.16a | |
| 100 | 9.43 ± 0.23b | 2.12 ± 0.16b | 2.57 ± 0.12a |
The mean and SE are shown for each nutrient treatment. Column means ± SE followed by different letters indicate significant differences among the treatments according to Duncan’s test at α = 0.05.
Table 3.
F-values and significance (p-values) of the two-way ANOVA on the effects of nutrient treatment, root diameter, and their interaction on the root length, root surface area, and root volume of the P. chinensis seedlings at final harvest.
Table 3.
F-values and significance (p-values) of the two-way ANOVA on the effects of nutrient treatment, root diameter, and their interaction on the root length, root surface area, and root volume of the P. chinensis seedlings at final harvest.
| Source | Root Length | Root Surface Area | Root Volume |
|---|---|---|---|
| Nutrient treatment | 39.4 (<0.001) | 28.6 (<0.001) | 14.2 (<0.001) |
| Root diameter | 464.2 (<0.001) | 83.2 (<0.001) | 168.3 (<0.001) |
| Nutrient treatment × Root diameter | 7.9 (<0.001) | 2.0 (0.019) | 7.2 (<0.001) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Song, X.; Wan, F.; Chang, X.; Zhang, J.; Sun, M.; Liu, Y. Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings. Forests 2019, 10, 1035. https://doi.org/10.3390/f10111035
AMA Style
Song X, Wan F, Chang X, Zhang J, Sun M, Liu Y. Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings. Forests. 2019; 10(11):1035. https://doi.org/10.3390/f10111035
Chicago/Turabian StyleSong, Xiehai, Fangfang Wan, Xiaochao Chang, Jin Zhang, Minghui Sun, and Yong Liu. 2019. "Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia chinensis Bunge Seedlings" Forests 10, no. 11: 1035. https://doi.org/10.3390/f10111035
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.