Subirrigation Effects on Larch Seedling Growth, Root Morphology, and Media Chemistry

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Nursery Cultural Conditions
2.2. Root Morphology and EC Measurement
2.3. Statistical Analysis
3. Results
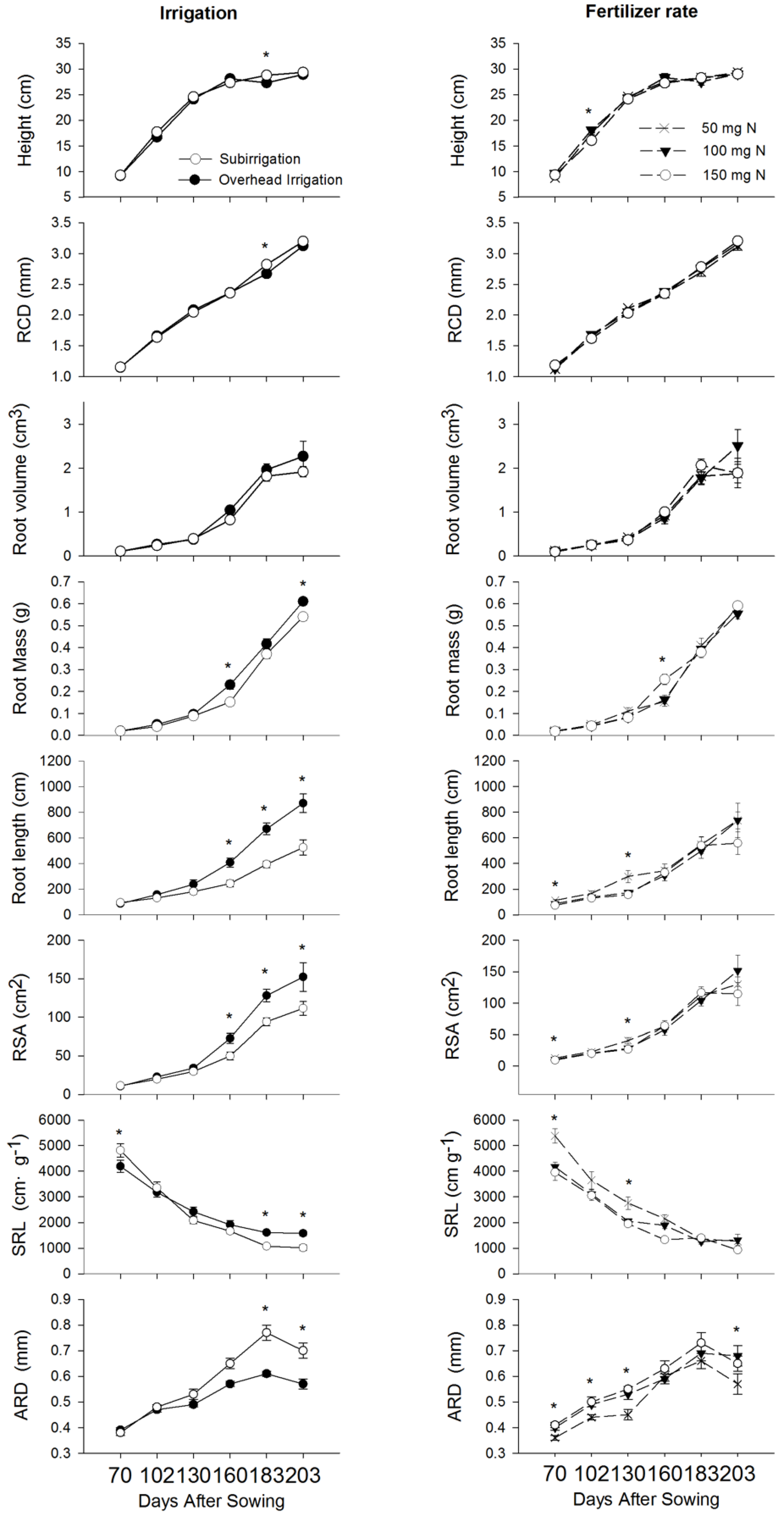
3.1. Effects of Irrigation and Fertilizer Rate on Larch Seedling Morphology
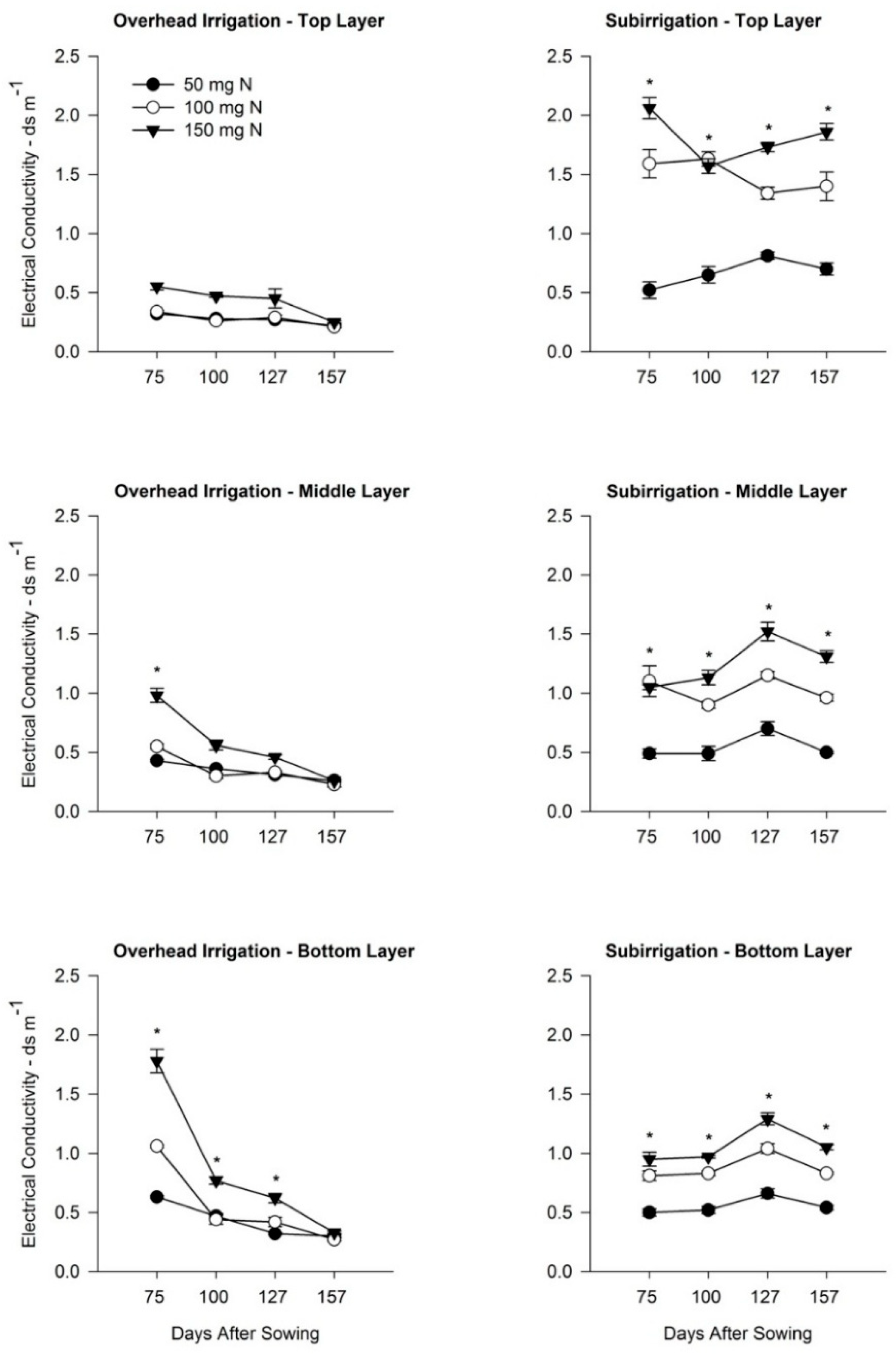
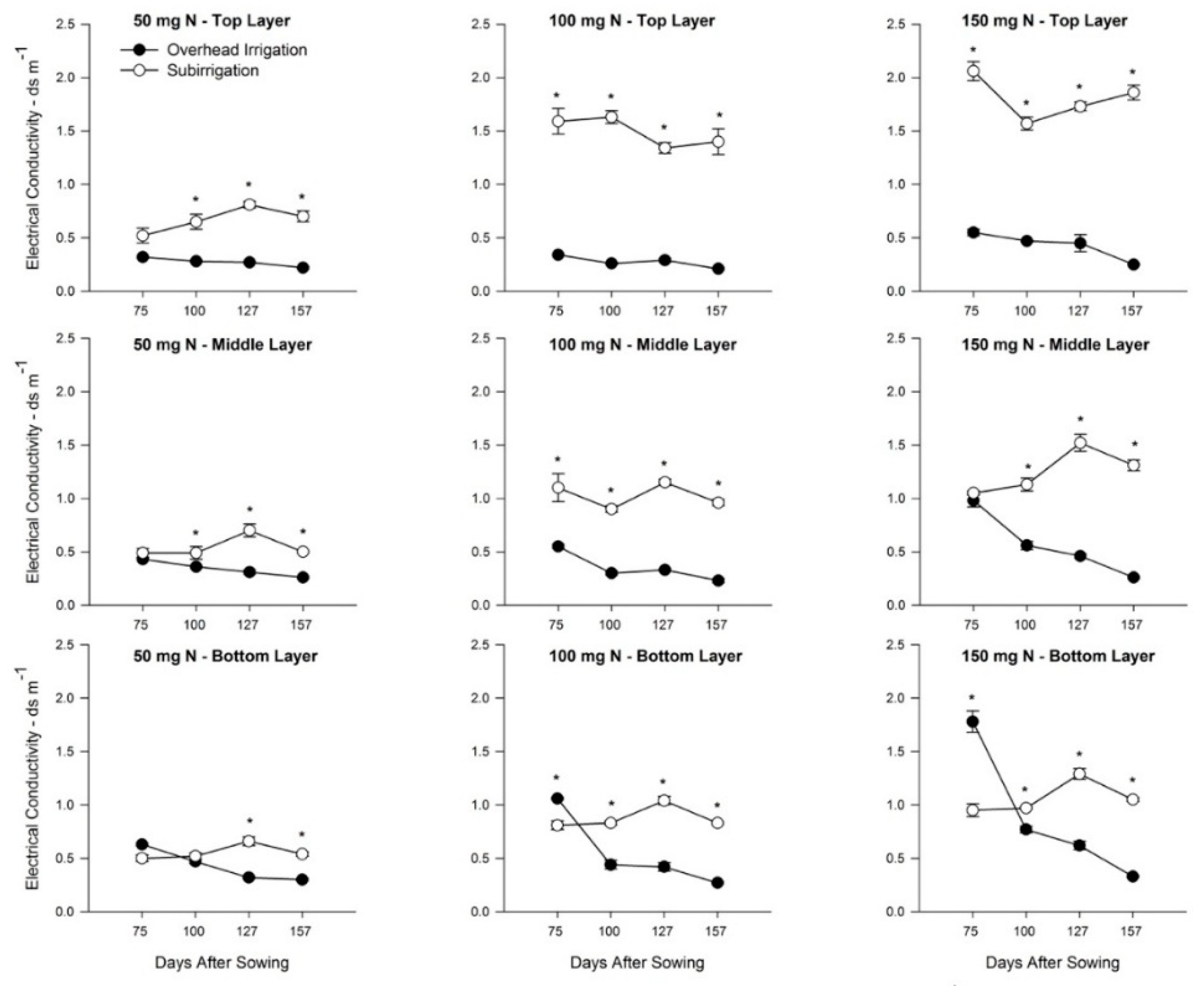
3.2. Effects of Irrigation and Fertilizer Rate on Growing Media EC
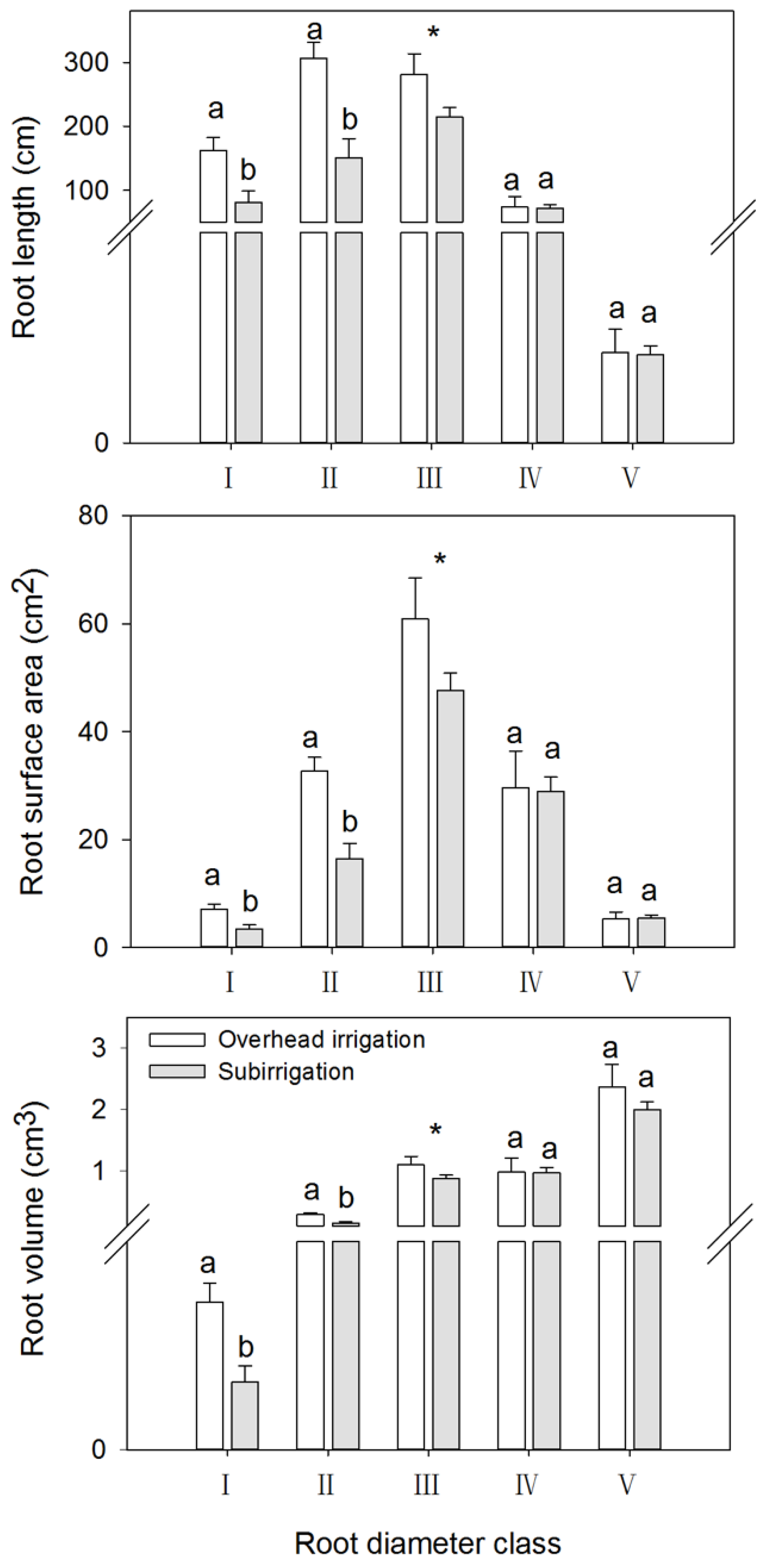
3.3. Effects of Irrigation and Fertilizer Rate among Root Diameter Classes
4. Discussion
4.1. Effects of Irrigation on Shoot Morphology and Root Architecture
4.2. Effects of Fertilizer Rates on Roots Morphology
4.3. Root Morphology Response to Growing Media EC Changes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haase, D.L.; Davis, A.S. Developing and supporting quality nursery facilities and staff are necessary to meet global forest and landscape restoration needs. Reforesta 2017, 4, 69–93. [Google Scholar] [CrossRef]
- Argo, W.R.; Biernbaum, J.A. The effect of irrigation method, water-soluble fertilization, preplant nutrient charge, and surface evaporation of early vegetative and root growth of poinsettia. J. Am. Soc. Hortic. 1995, 120, 163–169. [Google Scholar]
- Biernbaurn, J.A. Root-zone management of greenhouse container grown crops to control water and fertilizer use. HortTechnology 1992, 2, 127–132. [Google Scholar] [CrossRef]
- Landis, T.D.; Tinus, R.W.; McDonald, S.E.; Barnett, J.P. Seedling nutrition and irrigation. In The Container Tree Nursery Manual; USDA Forest Service: Washington, DC, USA, 1989; Volume 4. [Google Scholar]
- Dumroese, R.K.; Page-Dumroese, D.S.; Salifu, K.F.; Jacobs, D.F. Exponential fertilization of Pinus monticola, seedlings: Nutrient uptake efficiency, leaching fractions, and early outplanting performance. Can. J. For. Res. 2005, 35, 2961–2967. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Pinto, J.R.; Jacobs, D.F.; Davis, A.S.; Horiuchi, B. Subirrigation reduces water use, nitrogen loss, and moss growth in a container nursery. Nativ. Plants J. 2006, 7, 253–261. [Google Scholar]
- Dumroese, R.K.; Jacobs, D.F.; Davis, A.S.; Pinto, J.R.; Landis, T.D. An introduction to subirrigation in forest and conservation nurseries and some preliminary results of demonstrations. Phys. Rev. 2007, 165, 582–591. [Google Scholar]
- Dumroese, R.K.; Davis, A.S.; Jacobs, D.F. Nursery response of acacia koa seedlings to container size, irrigation method, and fertilization rate. J. Plant Nutr. 2011, 34, 877–887. [Google Scholar] [CrossRef]
- Cox, D. Growth, nutrient content, and growth medium electrical conductivity of poinsettia irrigated by subirrigation or from overhead. J. Plant Nutr. 2001, 24, 11. [Google Scholar] [CrossRef]
- Davis, A.S.; Jacobs, D.F.; Overton, R.P.; Dumroese, R.K. Influence of irrigation method and container type on northern red oak seedling growth and media electrical conductivity. Nativ. Plants J. 2008, 9, 5–12. [Google Scholar] [CrossRef]
- Davis, A.S.; Aghai, M.M.; Pinto, J.R.; Apostol, K.G. Growth, gas exchange, foliar nitrogen content, and water use of subirrigated and overhead-irrigated Populus tremuloides Michx. seedlings. Hortscience 2011, 46, 1249–1253. [Google Scholar]
- Ferrarezi, R.S.; Weaver, G.M.; Iersel, M.W.V.; Testezlaf, R. Subirrigation: Historical overview, challenges, and future prospects. Horttechnology 2015, 25, 262–276. [Google Scholar] [CrossRef]
- Hunter, M.; Leong, G.; Mitchell, J.; Dieters, M.; Fujinuma, R. Constant water table sub-irrigation of pots allows derivation of root weights (without physical recovery) and repeated measures of in situ growth and water use efficiencies. Plant Soil 2017, 1–19. [Google Scholar] [CrossRef]
- Montesano, F.; Parente, A.; Santamaria, P. Closed cycle subirrigation with low concentration nutrient solution can be used for soilless tomato production in saline conditions. Sci. Hortic. 2010, 124, 338–344. [Google Scholar] [CrossRef]
- Macherla, K.; Mcavoy, R.J. The effect of salinity on the growth and nutrient status of zinnia grown under short- and long-cycle subirrigation management. Hortscience 2017, 52, 770–773. [Google Scholar] [CrossRef]
- Sun, Q.; Dumroese, R.K.; Liu, Y. Container volume and subirrigation schedule influence Quercus variabilis seedling growth and nutrient status in the nursery and field. Scand. J. For. Res. 2018, 33, 560–567. [Google Scholar] [CrossRef]
- Klockmoore, K.A.; Broschat, T.K. Use of subirrigation to reduce fertilizer runoff. Proc. Fla. State Hortic. Soc. 2000, 113, 149–151. [Google Scholar]
- Uva, W.F.L.; Weiler, T.C.; Milligan, R.A. Economic analysis of adopting zero runoff subirrigation systems in greenhouse operations in the northeast and north central united states. Hortscience 2001, 36, 221–224. [Google Scholar]
- Hong, C.X.; Moorman, G.W. Plant pathogens in irrigation water: Challenges and opportunities. Crit. Rev. Plant Sci. 2005, 24, 189–208. [Google Scholar] [CrossRef]
- Strong, S.S.; Behe, B.K.; Deneke, C.F.; Bowen, K.L.; Keever, G.J. Cultivar and spacing effects on transmission of phytophthora parasitica in an ebb-and-flow subirrigation system. Plant Dis. 1997, 81, 89–95. [Google Scholar] [CrossRef]
- Elmer, W.H.; Mpn, G.; Mcavoy, R.J. Partial saturation under ebb and flow irrigation suppresses pythium root rot of ornamentals. Crop Prot. 2012, 33, 29–33. [Google Scholar] [CrossRef]
- Kent, M.W.; Reed, D.W. Nitrogen nutrition of new guinea impatiens ‘barbados’ and spathiphyllum ‘petite’ in a subirrigation system. J. Am. Soc. Hortic. Sci. 1996, 121, 816–819. [Google Scholar]
- Todd, N.M.; Reed, D.W. Characterizing salinity limits of new guinea impatiens in recirculating subirrigation. J. Am. Soc. Hortic. Sci. Am. Soc. Hortic. Sci. 1998, 123, 156–160. [Google Scholar]
- Richards, D.L.; Reed, D.W. New guinea impatiens growth response and nutrient release from controlled-release fertilizer in a recirculating subirrigation and top-watering system. Hortscience 2004, 39, 280–286. [Google Scholar]
- Bumgarner, M.L.; Salifu, K.F.; Mickelbart, M.V.; Jacobs, D.F. Effects of fertilization on media chemistry and Quercus rubra seedling development under subirrigation. Hortscience 2015, 50, 454–460. [Google Scholar]
- Pinto, J.R.; Chandler, R.A.; Dumroese, R.K. Growth, nitrogen use efficiency, and leachate comparison of subirrigated and overhead irrigated pale purple coneflower seedlings. Hortscience 2008, 43, 897–901. [Google Scholar]
- Jacobs, D.F.; Timmer, V.R. Fertilizer-induced changes in rhizosphere electrical conductivity: Relation to forest tree seedling root system growth and function. New For. 2005, 30, 147–166. [Google Scholar] [CrossRef]
- Doi, R.; Tanikawa, T.; Miyatani, K.; Hirano, Y. Intraspecific variation in morphological traits of root branch orders in Chamaecyparis obtusa. Plant Soil 2017, 1–11. [Google Scholar] [CrossRef]
- Wang, G.; Liu, F.; Xue, S. Nitrogen addition enhanced water uptake by affecting fine root morphology and coarse root anatomy of chinese pine seedlings. Plant Soil 2017, 1–13. [Google Scholar] [CrossRef]
- Adams, T.S.; Mccormack, M.L.; Eissenstat, D.M. Foraging strategies in trees of different root morphology: The role of root lifespan. Tree Physiol. 2013, 33, 940–948. [Google Scholar] [CrossRef]
- Jia, S.; Mclaughlin, N.B.; Gu, J.; Li, X.; Wang, Z. Relationships between root respiration rate and root morphology, chemistry and anatomy in Larix gmelinii and Fraxinus mandshurica. Tree Physiol. 2013, 33, 579. [Google Scholar] [CrossRef]
- Ostonen, I.; Helmisaari, H.S.; Borken, W.; Tedersoo, L.; Mai, K.; Bahram, M.; Lindroos, A.-J.; Nojd, P.; Uri, V.; Merila, P.; et al. Fine root foraging strategies in Norway spruce forests across a European climate gradient. Glob. Chang. Biol. 2011, 17, 3620–3632. [Google Scholar] [CrossRef]
- Miyatani, K.; Mizusawa, Y.; Okada, K.; Tanikawa, T.; Makita, N.; Hirano, Y. Fine root traits in Chamaecyparis obtusa, forest soils with different acid buffering capacities. Trees 2016, 30, 415–429. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Laskowski, M.J.; Burton, A.J.; Lessard, V.C.; Zak, D.R. Variation in sugar maple root respiration with root diameter and soil depth. Tree Physiol. 1998, 18, 665–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008; pp. 101–149. [Google Scholar]
- Burton, A.J.; Jarvey, J.C.; Jarvi, M.P.; Zak, D.R.; Pregitzer, K.S. Chronic N deposition alters root respiration-tissue N relationship in northern hardwood forests. Glob. Chang. Biol. 2011, 18, 258–266. [Google Scholar] [CrossRef]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.R.; Lõhmus, K.; Majdi, H.; Metcalfe, D.; Olsthoorn, A.F.M.; Pronk, A.; et al. Specific root length as an indicator of environmental change. G. Bot. Ital. 2007, 141, 426–442. [Google Scholar] [CrossRef] [Green Version]
- Xi, W.; Liu, Y.; Ma, L.; Li, G.; Jia, Z.; Lou, J.; Hu, J.; Wang, Y. Effects of sub-irrigation with different water and fertilizer supplies on growth, media pH and electric conductance of containerized Larix principis-rupprechtii seedlings. Sci. Silvae Sin. 2015, 51, 36–43. [Google Scholar]
- Teng, F.; Liu, Y.; Wang, Y.; Hu, J.; Sun, Q.; Wan, F.; Zhang, J. Sub-irrigation with different container types and sizes for containerized root growth of Pinus tabuliformis seedlings. J. Zhejiang A F Univ. 2017, 34, 449–458. [Google Scholar]
- Dumroese, R.K.; Pinto, J.R.; Montville, M.E. Using container weights to determine irrigation needs: A simple method. Nativ. Plants J. 2015, 16, 67–71. [Google Scholar] [CrossRef]
- Scoggins, H.L.; Van Iersel, M.W. In situ probes for measurement of electrical conductivity of soilless substrates: Effects of temperature and substrate moisture content. Hortscience 2006, 41, 210–214. [Google Scholar]
- Bumgarner, M.L.; Salifu, K.F.; Jacobs, D.F. Subirrigation of Quercus rubra seedlings: Nursery stock quality, media chemistry, and early field performance. Hortscience 2008, 43, 2179–2185. [Google Scholar]
- Hirano, Y.; Mizoguchi, T.; Brunner, I. Root parameters of forest trees as sensitive indicators of acidifying pollutants: A review of research of Japanese forest trees. J. For. Res. 2007, 12, 134–142. [Google Scholar] [CrossRef]
- Bechmann, M.; Schneider, C.; Carminati, A.; Vetterlein, D.; Attinger, S.; Hildebrandt, A. Parameterizing complex root water uptake models—The arrangement of root hydraulic properties within the root architecture affects dynamics and efficiency of root water uptake. Hydrol. Earth Syst. Sci. Discuss. 2014, 11, 757–805. [Google Scholar] [CrossRef]
- Jia, S.; Wang, Z.; Li, X.; Sun, Y.; Zhang, X.; Liang, A. N fertilization affects on soil respiration, microbial biomass and root respiration in Larix gmelinii and Fraxinus mandshurica plantations in china. Plant Soil 2010, 333, 325–336. [Google Scholar] [CrossRef]
- George, E.; Kircher, S.; Schwarz, P.; Tesar, A.; Seith, B. Effect of varied soil nitrogen supply on growth and nutrient uptake of young Norway spruce plants grown in a shaded environment. J. Plant Nutr. Soil Sci. 1999, 162, 301–307. [Google Scholar] [CrossRef]
- Hill, J.O.; Simpson, R.J.; Moore, A.D.; Chapman, D.F. Morphology and response of roots of pasture species to phosphorus and nitrogen nutrition. Plant Soil 2006, 286, 7–19. [Google Scholar] [CrossRef]
- Cuesta, B.; Vega, J.; Villar-Salvador, P.; Rey-Benayas, J. Root growth dynamics of aleppo pine (Pinus halepensis Mill.) seedlings in relation to shoot elongation, plant size and tissue nitrogen concentration. Trees 2010, 24, 899–908. [Google Scholar] [CrossRef]
- Fan, J.W.; Du, Y.L.; Turner, N.C.; Wang, B.R.; Fang, Y.; Xi, Y.; Guo, X.; Li, F. Changes in root morphology and physiology to limited phosphorus and moisture in a locally-selected cultivar and an introduced cultivar of Medicago sativa, l. growing in alkaline soil. Plant Soil 2015, 392, 215–226. [Google Scholar] [CrossRef]
- Shi, W.; Bloomberg, M.; Li, G.; Su, S.; Jia, L. Combined effects of cotyledon excision and nursery fertilization on root growth, nutrient status and outplanting performance of Quercus variabilis container seedlings. PLoS ONE 2017, 12, e0177002. [Google Scholar] [CrossRef]
- Genenger, M.; Zimmermann, S.; Hallenbarter, D.; Landolt, W.; Frossard, E.; Brunner, I. Fine root growth and element concentrations of Norway spruce as affected by wood ash and liquid fertilisation. Plant Soil 2003, 255, 253–264. [Google Scholar] [CrossRef]
- Deng, S.; Yin, Q.; Zhang, S.; Shi, K.; Jia, Z.; Ma, L. Drip irrigation affects the morphology and distribution of olive roots. Hortscience 2017, 52, 1298–1306. [Google Scholar] [CrossRef]
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Ding, G.; Shi, J.; Zhu, J.; Zhao, L. Effect of fertilization on fine root diameter, root length and specific root length in Larix kaempferi plantation. Chin. J. Appl. Ecol. 2007, 18, 957–962. [Google Scholar]
- Wang, G.; Fahey, T.J.; Xue, S.; Liu, F. Root morphology and architecture respond to N addition in Pinus tabuliformis, west China. Oecologia 2013, 171, 583–590. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Growth Parameter | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Height (cm) | Root Collar Diameter (mm) | Stem Mass (g) | Root Mass (g) | Whole Seedling Mass (g) (Stem + Root) | Root Length (cm) | Root Surface Area (cm2) | Root Volume (cm3) | Average Diameter (mm) | Specific Root Length (cm/g) | |
| Irrigation (I) | ||||||||||
| Overhead irrigation (OI) | 28.94 ± 0.36a | 3.13 ± 0.04a | 0.58 ± 0.02a | 0.61 ± 0.01a | 1.19 ± 0.03a | 869.96 ± 72.83a | 152.4 ± 18.63a | 2.27 ± 0.35a | 0.57 ± 0.02b | 1368.67 ± 136.82a |
| Subirrigation (SI) | 29.34 ± 0.29a | 3.20 ± 0.06a | 0.63 ± 0.02a | 0.54 ± 0.02b | 1.17 ± 0.04a | 524.87 ± 59.22b | 111.46 ± 8.92b | 1.92 ± 0.12a | 0.70 ± 0.03a | 981.07 ± 111.49b |
| Fertilizer Rate (F) | ||||||||||
| 50 mg·N·seedling−1 | 29.36 ± 0.23a | 3.12 ± 0.07a | 0.62 ± 0.03a | 0.58 ± 0.02a | 1.20 ± 0.05a | 736.88 ± 64.82a | 130.02 ± 11.67a | 1.88 ± 0.21a | 0.57 ± 0.04b | 1269.85 ± 104.20a |
| 100 mg·N·seedling−1 | 29.04 ± 0.42a | 3.16 ± 0.07a | 0.60 ± 0.02a | 0.55 ± 0.02a | 1.15 ± 0.04a | 735.90 ± 133.06a | 151.39 ± 24.64a | 2.51 ± 0.37a | 0.68 ± 0.04a | 1320.78 ± 223.47a |
| 150 mg·N·seedling−1 | 29.02 ± 0.53a | 3.20 ± 0.05a | 0.59 ± 0.03a | 0.59 ± 0.02a | 1.18 ± 0.03a | 558.69 ± 87.05a | 114.37 ± 18.53a | 1.89 ± 0.33a | 0.65 ± 0.03ab | 933.99 ± 134.86a |
| p-values | ||||||||||
| Irrigation (I) | 0.434 | 0.363 | 0.122 | 0.004 | 0.695 | 0.003 | 0.023 | 0.323 | 0.001 | 0.028 |
| Fertilizer Rate (F) | 0.822 | 0.692 | 0.723 | 0.370 | 0.695 | 0.448 | 0.214 | 0.262 | 0.025 | 0.125 |
| I × F | 0.950 | 0.576 | 0.754 | 0.986 | 0.869 | 0.100 | 0.075 | 0.215 | 0.703 | 0.132 |
| Time | Source | p Value | ||
|---|---|---|---|---|
| Top | Middle | Bottom | ||
| 75 DAS | Irrigation (I) | <0.001 | 0.001 | <0.001 |
| Fertilizer Rate (F) | <0.001 | <0.001 | <0.001 | |
| I × F | <0.001 | 0.002 | <0.001 | |
| 100 DAS | Irrigation (I) | <0.001 | <0.001 | <0.001 |
| Fertilizer Rate (F) | <0.001 | <0.001 | <0.001 | |
| I × F | <0.001 | <0.001 | <0.001 | |
| 127 DAS | Irrigation (I) | <0.001 | <0.001 | <0.001 |
| Fertilizer Rate (F) | <0.001 | <0.001 | <0.001 | |
| I × F | <0.001 | <0.001 | 0.002 | |
| 157 DAS | Irrigation (I) | <0.001 | <0.001 | <0.001 |
| Fertilizer Rate (F) | <0.001 | <0.001 | <0.001 | |
| I × F | <0.001 | <0.001 | <0.001 | |
| Parameter | Source | Class I | Class II | Class III | Class IV | Class V |
|---|---|---|---|---|---|---|
| Root length | Irrigation (I) | 0.004 | <0.001 | 0.023 | 0.893 | 0.939 |
| Fertilizer Rate (F) | 0.102 | 0.013 | 0.025 | 0.297 | 0.441 | |
| I × F | 0.461 | 0.229 | 0.001 | 0.435 | 0.738 | |
| Root surface area | Irrigation (I) | 0.003 | <0.001 | 0.023 | 0.925 | 0.926 |
| Fertilizer Rate (F) | 0.062 | 0.024 | 0.061 | 0.323 | 0.485 | |
| I × F | 0.593 | 0.214 | 0.004 | 0.494 | 0.759 | |
| Root volume | Irrigation (I) | 0.003 | <0.001 | 0.040 | 0.956 | 0.334 |
| Fertilizer Rate (F) | 0.047 | 0.030 | 0.061 | 0.350 | 0.285 | |
| I × F | 0.684 | 0.136 | 0.005 | 0.560 | 0.232 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, F.; Ross-Davis, A.L.; Shi, W.; Weston, C.; Song, X.; Chang, X.; Davis, A.S.; Liu, Y.; Teng, F. Subirrigation Effects on Larch Seedling Growth, Root Morphology, and Media Chemistry. Forests 2019, 10, 38. https://doi.org/10.3390/f10010038
Wan F, Ross-Davis AL, Shi W, Weston C, Song X, Chang X, Davis AS, Liu Y, Teng F. Subirrigation Effects on Larch Seedling Growth, Root Morphology, and Media Chemistry. Forests. 2019; 10(1):38. https://doi.org/10.3390/f10010038
Chicago/Turabian StyleWan, Fangfang, Amy L. Ross-Davis, Wenhui Shi, Christopher Weston, Xiehai Song, Xiaochao Chang, Anthony S. Davis, Yong Liu, and Fei Teng. 2019. "Subirrigation Effects on Larch Seedling Growth, Root Morphology, and Media Chemistry" Forests 10, no. 1: 38. https://doi.org/10.3390/f10010038