Fungal Endophytes as Biocontrol Agents against the Main Soil-Borne Diseases of Melon and Watermelon in Spain


1
Unidad de Protección Vegetal, Centro de Investigación y Tecnología Agroalimentaria de Aragón (CITA), Avda. Montañana, 930, 50059 Zaragoza, Spain
2
Instituto Agroalimentario de Aragón-IA2 (CITA-Universidad de Zaragoza), 50013 Zaragoza, Spain
3
Unidad de Hortofruticultura, Centro de Investigación y Tecnología Agroalimentaria de Aragón, Avda. Montañana, 930, 50059 Zaragoza, Spain
*
Author to whom correspondence should be addressed.
Agronomy 2020, 10(6), 820; https://doi.org/10.3390/agronomy10060820
Submission received: 8 May 2020
/
Revised: 1 June 2020
/
Accepted: 6 June 2020
/
Published: 9 June 2020
(This article belongs to the Special Issue Recent Advances in Genomics, Genetic Resources Evaluation and Breeding of Cucurbitaceae Crops)
Abstract
:Watermelon and melon crops are affected by some important soil-borne fungal diseases like carbonaceous rot (Macrophomina phaseolina), collapse (Monosporascus cannonballus), or the most important pathology at an economic level, the Fusarium wilt (Fusarium oxysporum f. sp. niveum, F. oxysporum f. sp. melonis, F. solani f. sp. cucurbitae, Neocosmospora falciformis, and N. keratoplastica). The methods commonly used for their control are often ineffective, thus new approaches, as the use of biological control agents, are constantly being sought. This work aimed to isolate, identify, and test endophytic fungi for their antagonistic properties against the three mentioned diseases. For this, about 350 endophytic fungal strains were isolated from asymptomatic watermelon plants. Among these, 7 fungal species were selected to evaluate their antagonistic potential against 14 pathogens. Dual culture assays allowed to select two Trichoderma strains according to the high inhibition rates observed (up to 93%), that were further employed in melon and watermelon plants, showing that some of the pathogens were controlled in terms of disease incidence, exhibiting a decrease up to 67% for T. lentiforme. In addition, three concentrations of Epicoccum purpurascens extract was selected to evaluate the germicide effect, obtaining significant differences in the growth of the pathogens depending on fermentation parameters.
1. Introduction
Watermelon (Citrullus lanatus [Thunb.] Matsum. & Nakai) and melon (Cucumis melo L.) are horticultural species of the Cucurbitaceae, widely cultivated throughout the world. Both species represent economically important crops, especially in Spain, which is the most important producer in Europe, with an annual production of 853,600 tons [1]. An important limiting factor for these cultures around the world has long been the several diseases and pests that affect them, causing significant losses [2,3]. Some of these pathologies have been recently characterized, while others are known for more than a century [4]. In Spain and other producing areas of the Mediterranean, diseases like collapse produced by Monosporascus cannonballus Pollack and Uecker (Diatrypaceae), the so-called charcoal rot caused by Macrophomina phaseolina (Tassi) Goid. (Botryospharieceae) and Fusarium wilt are responsible for annual important yield losses in both greenhouses and open–air fields [5,6,7,8].
Monosporascus cannonballus infects the root of many species of cucurbits in warm, arid, and semi-arid areas, and also in subtropical regions around the world [9]. The most visible symptoms of its infection are located at the root level, consisting of active rot and necrotic lesions, which causes wilting and gradual yellowing, and collapse of the plant [10]. Macrophomina phaseolina is considered as one of the most harmful seed and soil-borne diseases. Its symptoms in watermelon and melon are similar, with the presence of yellowing and death of the growing buds accompanied by aqueous lesions that surround affected plant tissues, also with exudates that appear dark brown when drying, and even cracked areas in the stem [10]. Together with these, special attention must be put on a group of diseases commonly named as vascular fusariosis (Fusarium wilt). Symptoms of Fusarium wilt can appear at any phenological stage, attacking seeds, seedlings, or adult plants. Several species and formae specialis (f. sp.) of the genus Fusarium are known to be responsible for this disease. There are three well-known species implicated, with slight differences and particularities according to host range: Fusarium oxysporum f. sp. niveum (E.F. Sm.) W.C. Snyder and H.N. Hansen (Fon), F. oxysporum f. sp. melonis (Leach and Currence) W.C. Snyder and H.N. Hansen (Fom) and F. solani (Mart.) Sacc. f. sp. cucurbitae W.C. Snyder and H.N. Hansen (Fsc) (actually included in genus Neocosmospora). The first one (Fon), with four pathogenic races (race 0, 1, 2, and 3), specifically affects watermelon [11], while Fom, with four pathogenic races (0, 1, 2, and 1.2) [12], is reported to be associated with melon. The third mentioned species, Fsc, is associated with cucurbit species like watermelon, melon, squash, or pumpkin. For Fsc there have been described two pathogenic races, 1 and 2, currently named Neocosmospora cucurbitae Sand.–Den., L. Lombard and Crous, and N. petroliphila (Q.T. Chen and X.H. Fu) Sand.-Den. and Crous, respectively, both cited at the national level [7,8,12]. In addition, the other two species, N. falciformis (Carrión) L. Lombard and Crous (=F. falciforme) and N. keratoplastica (D. Geiser, O’Donnell, Short, and Ning Zhang) Sand.-Den. and Crous (=F. keratoplasticum), of the so-called F. solani species complex (FSSC) have been reported to cause Fusarium wilt and root and stem rot in melon, watermelon, and squash [8,13].
Concerning disease management of cucurbits, factors associated with both climatic change and social demand are producing changes in agricultural systems. Currently, progress is made towards integrated and ecological production models, in which a limitation of the application of phytosanitary treatments is necessary [14]. Those integrated methods include the use of biological control agents (BCAs) for disease control [15], including those associated with cucurbits [16,17,18]. Optimization of biological control involves the use of microorganisms preferably isolated from the crop under the specific conditions in which they have to be applied, constituting the best way to get them more efficient [19]. In this respect, although studies testing the antagonistic capabilities of certain microorganisms against cucurbit pathogens have been reported, in most of these the BCAs were often isolated from other crops and/or environments [16,20,21], and only a few were focused on the isolation and characterization of fungal endophytes from cucurbits [22]. In-depth knowledge of microbial communities living endophytically in these plant hosts could allow the recruitment of microorganisms with the potential to be used as BCAs against the mentioned soil-borne diseases in Spain and other affected areas of Mediterranean. In the present work, we report the evaluation of the antagonistic potential of some endophytic fungi isolated from watermelon plants against a collection of some watermelon and melon pathogens. The potential BCAs identified in this study could be used to control the mentioned diseases per se or in combination with other useful strategies such as, for example, grafting and/or breeding.
2. Materials and Methods
2.1. Fungal Isolates
Plant materials prospected consisted of healthy watermelon plants and their associated rhizospheres, coming from two sampling surveys carried out in some of the main producing areas of Aragón (Spain). The first sampling was performed in a commercial field of Peñaflor (Zaragoza, Aragón, Spain), consisting of 11 watermelon plants. A second sampling survey was carried out in 2 watermelon commercial fields in Alfamén (Aragón, Zaragoza, Spain), where 6 plants were collected in each field. Watermelon plants were processed and surface sterilized with an EtOH solution (70%) for 1 min, then with a 5% (v/v) commercial sodium hypochlorite solution for 3 min, and finally, washed 4 times with sterile bi-distilled water. Then, plant fragments (roots and stems) were placed in 9 cm Ø Petri dishes with Potato Dextrose Agar (PDA) medium supplemented with streptomycin sulfate (17 g of PDA, 13 g of agar, 1000 mL of distilled water, 0.3 g of streptomycin sulfate). Plates were then incubated at 25 °C for 48 h in the dark to stimulate the development of endophytic fungi. After this, emerging colonies were transferred to 6 cm Ø PDA plates, and each isolate subcultured to get pure cultures for subsequent characterization.
2.2. Morphological and Molecular Characterization of Fungal Isolates
Fungal strains obtained were primarily identified and characterized morphologically. Morphological characterization was carried out based on the study and comparison of different somatic structures and/or sexual and asexual reproductive structures, from the observation of microscopical slides of pure colonies on PDA plates. Besides this, to further confirm the identity of some fungal isolates, molecular identification was made by direct sequencing of the ribosomal ITS (Internal Transcribed Spacer) fragment for those taxa usually reported in the specialized literature as microbial antagonists, as well as for certain sterile strains that did not show sufficient diagnostic characters. DNA extraction of each isolate was carried out with the Extract-N-Amp Plant kit PCR Kit (Sigma Aldrich Chemical Co., St. Louis, MO, USA) from mycelium of pure cultures, following manufacturer’s instructions. For ITS amplification, ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCT-TATTGATATGC-3′) primers were used. The final volume of each PCR reaction was 25 μL, containing 2 μL of genomic DNA of each isolate; 0.5 μL dNTPs, 0.8 μL MgCl2; 2.5 μL of 10× PCR; 1 μL of each ITS primer; 0.2 μL of Taq polymerase (Invitrogen, Carlsbad, CA, USA) and 17 μL H2O dd. PCR conditions were as follows: 1 initial cycle of 6 min at 94 °C, followed by 30 cycles of 1 min at 94 °C, 1 min at 54 °C and 1 min at 72 °C. Amplified products were sequenced by the Sanger method (Stab Vida, Caparica, Portugal). The ITS ribosomal DNA sequences obtained allowed the identification of isolates by their comparison with homologous sequences deposited in public databases like GenBank (using BLASTn tool) or Fusarium ID Database (http://www.westerdijkinstitute.nl/fusarium/).
2.3. Selection of Microbial Antagonists
Among the isolates obtained, 7 strains, belonging to species that are usually reported as biocontrol agents in the specialized literature, were selected to evaluate their antagonistic capacity against 14 pathogenic isolates of watermelon and melon belonging to the culture collection at CITA’s Mycology Lab (Table 1).
2.4. Dual Plate Cultures
All possible combinations of pathogen-antagonist were tested, as well as each fungus alone. Tests were performed using the dual culture technique [23] in 9 cm Ø Petri dishes containing Potato Dextrose Agar (PDA) supplemented with streptomycin sulfate in 3 biological replicates. Then, two 4 mm Ø agar plugs with mycelium coming from a 2 to 5 days culture of both pathogens and antagonists were placed, each one at both ends of the plate, 2 cm from its margin, respectively. Individual controls for pathogens and antagonists were prepared by inoculating each of them (4 mm Ø discs) in the center of a 9 mm Ø PDA plate. Plates were incubated at 25 ± 1 °C for 10 days in the dark, taking measurements of the diameters of the colonies every 48 hours. To evaluate dual plate assays, 2 radial measurements were taken with the aid of a digital gauge (Power Fix Profi) for each confrontation. These radial measurements, together with the presence of hyphal interactions, or extracellular secretions, were employed to characterize the mechanism of action (hyperparasitism, antibiosis, etc.) exerted by each antagonist. For dual cultures of the Trichoderma species tested, mycelial tufts were removed from the contact area of the facing colonies and observed under optical microscopy to characterize the different structures and hyphal modifications associated with the hyperparasitism produced.
Quantification of the antagonistic effects were determined by calculating the Percentage of Radial Growth Inhibition (PRGI) [24,25]. An analysis of variance (ANOVA) of the PRGI values was carried out after 10 days of culture, identifying differences between treatments, with the Tukey significance test (p < 0.05). A statistical program JMP13 (JMP®, Version <13>. SAS Institute Inc., Cary, NC, USA, 1989–2007) was used. For each dual confrontation, a mean PRGI value of each of them was used to perform analysis of the variance. Moreover, the degree of invasion/destruction of the pathogen colonies caused by T. lentiforme and T. harzianum in dual cultures was recorded 10 days after inoculation, using a visual scale with values from 0–4 described by Ortiz & Orduz [26], in which values above 3 were considered a satisfactory antagonistic capacity for a given strain of Trichoderma.
2.5. Evaluation of Fungicide Effects of Secondary Metabolites from Epicoccum Purpurascens
In addition to dual cultures, the fungicide capacity of the different secondary metabolites produced by Epicoccum purpurascens was tested by incorporating a liquid extract of this species into solid agar media. To do this, 3 flasks containing 300 mL of CM liquid medium (5 g malt extract, 5 g yeast extract, 5 g glucose, 1000 mL of distilled water) were inoculated with 4 mycelial plugs coming from a fresh culture of the fungus. Flasks were shaken (135 rpm) and incubated at 25 °C during 5, 10, and 15 days post-inoculation (dpi), respectively. After these 3 incubation periods, each one of the cultures was centrifuged, and the supernatants were filtered with a vacuum filtration system of 0.45 µm pore (Nalgene, Thermo Fischer Scientific). Then particle-free filtrate obtained was incorporated at 3 concentrations (0.5%, 1%, and 2%) to PDA plates where the pathogenic isolates were inoculated at the center of the dish and incubated at 25 °C. Three repetitions were made per pathogen, days of extract, and concentration with their respective controls consisting of PDA plates without filtrate. The evaluation of the germicidal effect was determined by measuring the radial growth of the different colonies in comparison with the controls. An analysis of variance (ANOVA) was carried out, including 2 factors, days of fermentation before extracting and doses (%) supplemented to the medium. For comparison between means, a post hoc Tukey test at a 5% significance level was used. Finally, filtrate treatments of E. purpurascens, that showed significant values of inhibition of fungal growth were compared with those obtained from dual cultures with the same strain to determine which treatment exhibited the best antagonistic effect. For the statistical analysis of these results, a comparison of means with Student’s t-test at a 5% significance level was performed.
2.6. In Vivo Biocontrol Assay
To confirm the potential as biocontrol agents of the 2 Trichoderma isolates (T. harzianum and T. lentiforme) used in the dual culture plate bioassays detailed above, it was decided to test their antagonistic effect against a set of pathogens in watermelon and melon seedlings in a growth chamber. Thus, ‘Black Diamond’ watermelon seeds and ‘Charentais T’ melon seeds were impregnated with the fungicide TMTD (Pomasol) and deposited in a 9 mm Ø Petri dish on a bed of sterilized filter paper moistened with 5 mL of sterile bi-distilled water. Pre-germination was carried out at 28 °C for 5 days. Germinated seeds were then placed in plastic trays with substrate (Professional substrate, Projar, Valencia, Spain) previously autoclaved (20 min, 120 °C) and grown for 10 days in a growth chamber at 28 °C day/25 °C night (16 h day) cycle. For this bioassay, T. harzianum and T. lentiforme were used against 11 pathogens: Fon 0, Fon 1, Fon 2, Fsc R1, Fp, Mcr, and Mon for watermelon and Fom 0, Fom 1, Fom 2, and Fom 1.2w for melon, respectively.
All pathogens were previously grown in Petri dishes in PDA medium. Then, 9 flasks were prepared with 200 mL of liquid CM medium and inoculated with 3 mycelial plugs of each one of the pathogens (except for both Mcr and Mon, whose inoculum preparation is described below) and incubated at room temperature under agitation (120 rpm). After 3 days of incubation, the fermentation broth was filtered, and the conidial concentration of all isolates adjusted to 106 conidia/mL with the help of a haematocytometer. For Mcr and Mon, inoculum consisted of colonized wheat grains previously autoclaved for 3 h, inoculated with 6–8 agar plugs coming from fresh colonies of both taxa, and incubated in 12 mm Ø Petri dishes for 10 days at 25 °C in the dark. Watermelon and melon seedlings were then inoculated by adding these wheat grains to the potting substrate at 5 g/L.
Finally, a conidial solution (106 conidia/mL) of the antagonist isolates was also prepared. To do this, 9 mm Ø Petri dishes containing mature, sporulated colonies from both Trichoderma species were flooded with sterile bi-distilled water, and vigorously shaken to detach the conidia in the aqueous medium, and then collected in sterile conditions. The conidial solution thus obtained was adjusted to 106 conidia/mL and a final volume of 600 mL of inoculum was disposed per antagonist.
To perform the inoculation, both watermelon and melon seedlings were extracted from the substrate and their roots were washed with sterile water. Then, they were inoculated either by root immersion or by adding colonized wheat grains to the substrate. For the first case, they were submerged 2 min in the mixed conidial solutions of both each pathogen and the corresponding antagonist. In the case of wheat grains, plants were first submerged 2 min in the conidial solution of each Trichoderma species, and then they were sown in the potting medium containing those colonized wheat grains. For each treatment and combination, 6 plants were inoculated and sown in two pots each (3 + 3).
Control plants were disposed of, inoculated only with pathogens or antagonists and also negative controls consisting of non-inoculated plants. Pots were incubated in a growth chamber for 30 days in a 28 °C day/25 °C night (16 h day) cycles. Counts for disease progress were made at the end of the culture period, measuring the degree of symptomatology of the seedlings, employing a visual scale from 1 to 4 as follows: 1 = healthy plants; 2 = stunted growth and damage of up to 50%; 3 = presence of foliar lesions, yellowing, wilting and root damage; 4 = dead plants. From the annotated disease rate data, disease incidence (%) was obtained to determine percent wilt incidence, based on the number of wilted plants (values 3 and 4) indicate out of total plants [27]. Dead plants were removed at the end of the growth period and processed to fulfill Koch’s postulates.
3. Results
3.1. Endophytic Fungi Associated with Watermelon Plants. Selection of Microbial Antagonists
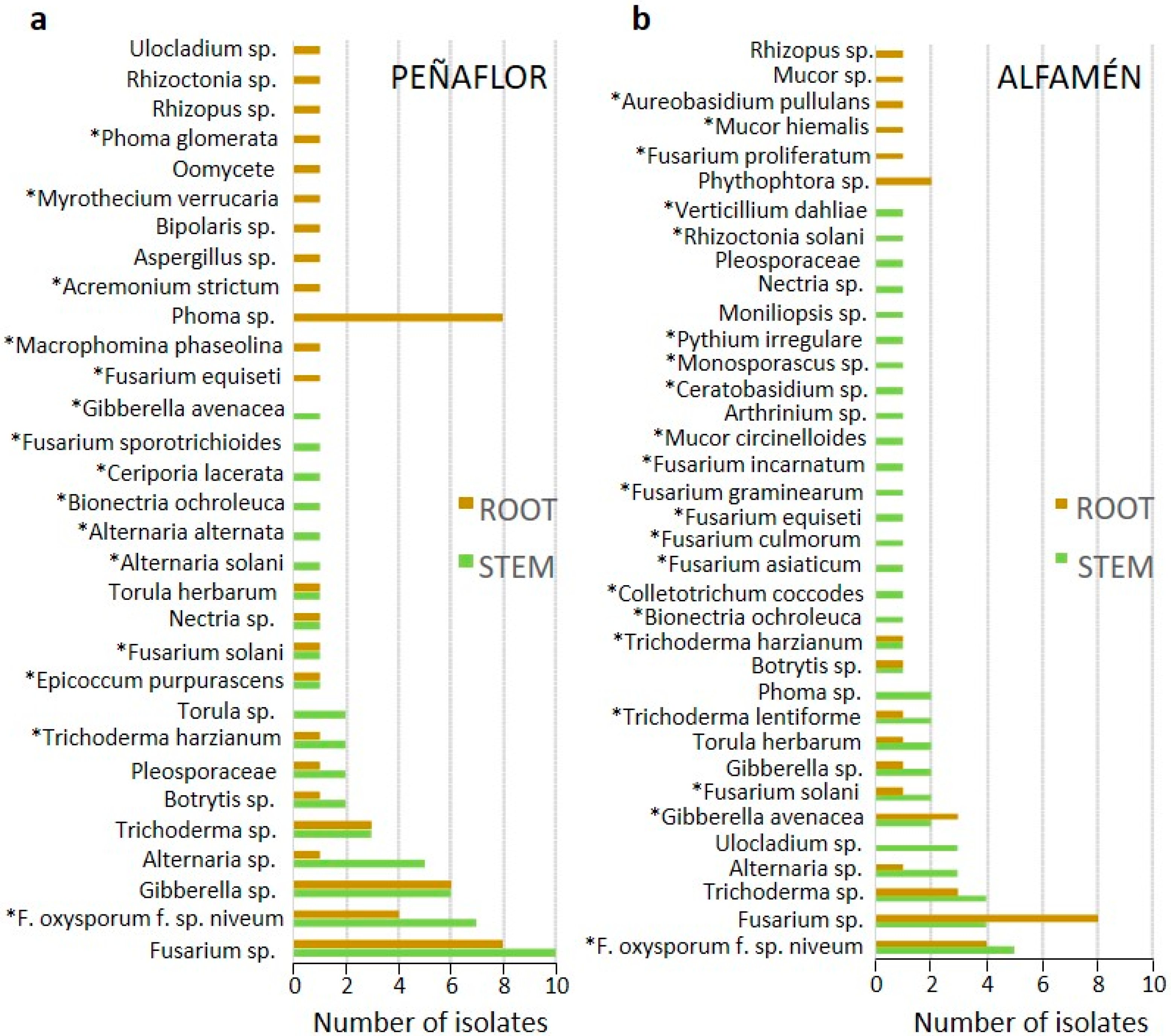
From a total of 23 watermelon plants surveyed, 347 endophytic fungal strains were obtained from the tissues analyzed (roots and stems), belonging to 67 OTUs (Operational Taxonomical Unit) (Figure 1). Most of endophytes recovered were not specifically associated with a given host tissue (phyllosphere, rhizosphere, rhizoplane, or vascular tissue). However, some degree of tissue specialization could be found, existing differences especially among the taxa recovered in both aerial and hypogeous tissues, although not so much at the level of the rhizosphere and the rhizoplane. Thus, only a few taxa like those belonging to genera Trichoderma, Fusarium, Phoma, Macrophomina, or Rhizoctonia were recovered from the rhizoplane, although species from these genera were also found infecting the inner root tissues or living saprophytically in the rhizosphere.
In the survey sampling carried out in Peñaflor, a total of 218 endophytic isolates (representing 31 OTUs) were identified; among them, the genus Fusarium was one of the most prevalent (31%), including isolates belonging to taxa like F. oxysporum f. sp. niveum (9%) or Gibberella (8%). Some of the isolates (23% from the total) could not be identified morphologically. In addition, certain isolates isolated here were identified as species with antagonistic potential, belonging to genera like Ceratobasidium, Bionectria, Myrothecium, Trichoderma, and Epicoccum. In one of the two fields of the locality of Alfamén, 92 isolates were identified (representing 25 OTUs). Again, some of the most prevalent isolates represented taxa from the genera Fusarium (25%), Giberella (9%), or Alternaria (7%). Finally, in the second field of Alfamén, 37 isolates (representing 11 OTUs) were identified, including species of genus Trichoderma (41%), pathogens like the above mentioned F. oxysporum f. sp. niveum (30%) and other potential antagonists such as Aureobasidium pullulans. After taxonomic identifications made up by morphological and/or molecular methods among all the isolates obtained, seven endophytic taxa belonging to species usually reported as ABCs were selected (Table 1).
3.2. Dual Culture Pathogens-Biocontrol Agents (BCA)
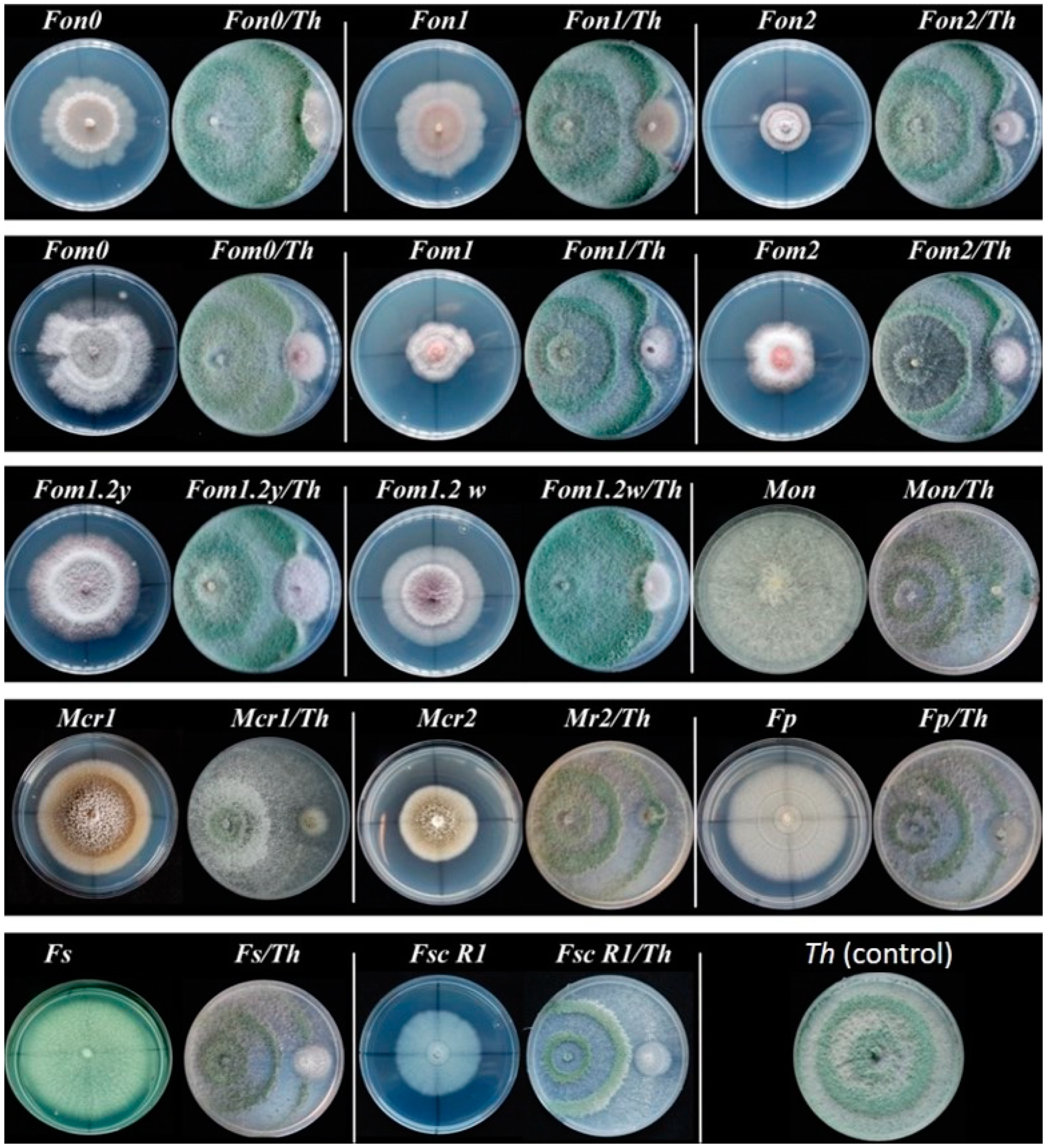
To evaluate the protective effect of the 7 isolates previously selected against the 14 pathogens employed, dual plate confrontations were made by culturing both species simultaneously. It was observed that T. harzianum (Th) exhibited significant high values of growth inhibition against the 14 pathogens selected (Table 2). The highest average PRGI value was obtained in the confrontation with Mon with 93.4%. It was visually observed how the antagonist stopped or minimized the diameter of the colony of this pathogen (Figure 2). Growth inhibition rates of Th with the rest of pathogens were always above 39.2%, corresponding to the lower mean value observed in confrontation with Fon 2.
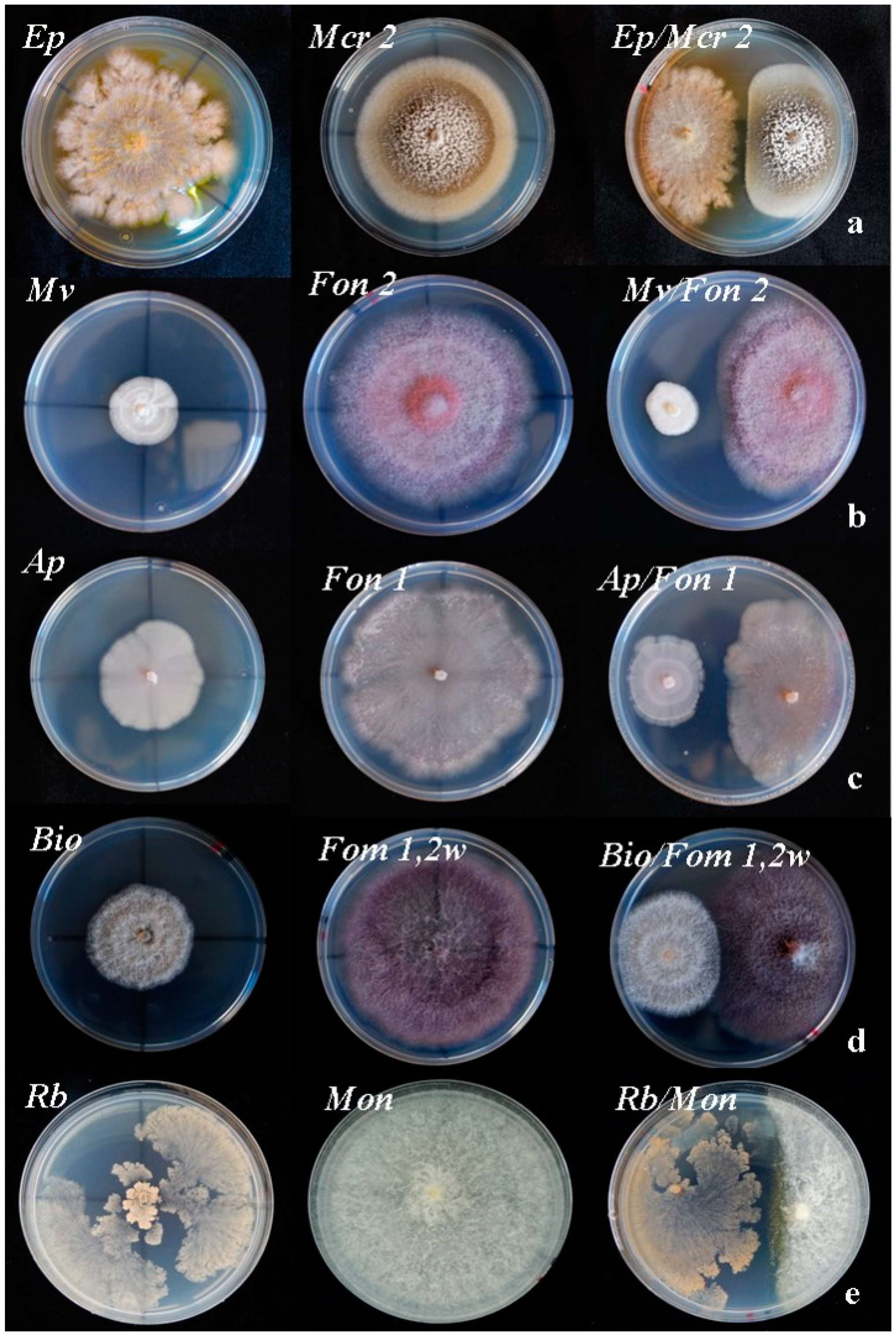
When analyzing the inhibition effects of T. lentiforme (Tl), it was observed that average PRGI values were slightly higher than in the case of T. harzianum, varying from 42.4% with Fom 1 to 93.3% for Mon. For the rest of the antagonistic strains (Figure 3), growth inhibition rates were in general lower. For Ep, the highest PRGI value was 41.3% in the confrontation with Macrophomina phaseolina isolate 2 (Mcr2) (Figure 3a), which represents a much lower PRGI value than those obtained with Trichoderma species. In plate confrontations, Mv exhibited low levels of antagonism with PRGI values between 27.2% and 3.7% (Figure 3b). In the dual confrontations against Ap it was observed that average PRGI values were found between 42.9% for Fon 1 and 7.3% for Fon 2 (Figure 3c). In the case of Bo, the average values of PRGI ranged from 32.9%, for Fom 1.2 w, to 7.7% for Fon 2 (Figure 3d).
Finally, in dual confrontations with Rb, average PRGI values were slightly higher than those observed for Ap and Bo. Thus, the values obtained ranged between 64.8% for the confrontation with Mon (Figure 3e) to 4.3% against both Mcr1 and Fsc R1.
On the other hand, the differences of each antagonist on each pathogen were also evaluated. The results of the post-hoc Tukey’s test showed that the mean value of PRGI were significantly different among the antagonists for all pathogens. For Fon 1, Fon 2, Fom 0, Fom 1.2w, Mon, Mcr1, Mcr2, Fp, and Fs, the Th and Tl antagonists caused higher mean percentages of inhibited of growth than the other antagonists tested, being the mean PRGI significantly higher than those caused by the rest of the antagonists (Supplementary material Table S1).
The degree of in vitro invasion and destruction of the mycelium of the two Trichoderma species employed on the pathogens was measured (Table 3). Both T. lentiforme and T. harzianum invaded the total surface of the M. cannonballus colony (Mon) and their mycelia were able to actively sporulate on it with a value of 4. Trichoderma lentiforme exhibited no invasion of the colony surface with the pathogens Fon 1, Fom 0, Fom 1, or Fom 2, as T. harzianum with the pathogens Fom 1, Fom 2, and Fsc R1. It should also be noted that the two strains of Trichoderma exhibited a profuse mycelial growth of cotton-white appearance, that after 4 days began to produce sporulation of olive green color in the confronting plates. This fact suggested that the pathogens tested did not produce any inhibitory effect on the Trichoderma spp., since their mycelium developed normally on the plate.
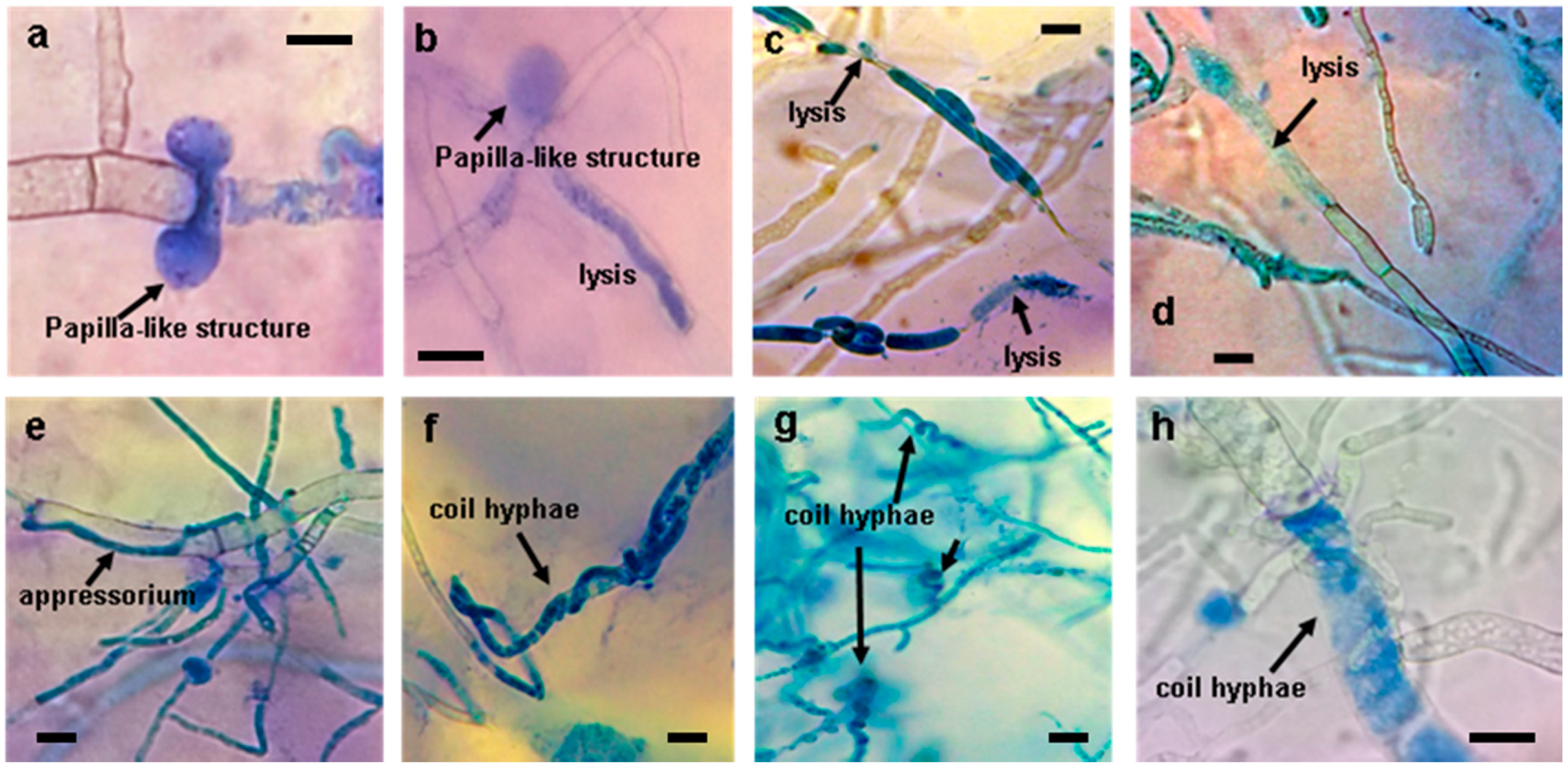
As a result of its hyperparasite behavior, different types of interactions and hyphal structures of Trichoderma spp. have been observed when Trichoderma interacted with pathogens. Some of these structures were present in the confrontations with the different pathogens that were inhibited in dual plate confrontation and included the formation of papilla-like structures, appressoria, lysis of pathogen hyphae, or the presence of typical coil hyphae in the antagonist (Figure 4).
3.3. Fungicide Effects of Secondary Metabolites from Epicoccum Purpurascens
The radial growth measured for the 14 pathogens inoculated with the three doses (0,5%, 1%, and 2%) of E. purpurascens (Ep) extract fermented by 5, 10, and 15 dpi are shown in Table 4. Decrease of the radial growth of pathogens inoculated with E. purpurascens metabolites was observed only for some of them, as Fon 0, Fom 2, Fom 1.2y, Fom 1.2w, and Fp, being the mean radial growth slightly lower than those without inoculating.
Analysis of variance revealed significant differences (p < 0.05) among the days, the doses, and the pathogens. There were also significant interactions between day and pathogen, dose and pathogen, as well as the three-way interaction (Table 5).
Whereas significant differences of the radial growth were found among the days of fermentation of the Ep for Fon 0, Fon 2, Fom 1, Fsc, and Mcr2; the radial growth varied significantly among both the days of fermentation of the Ep and the doses of Ep extract for isolates Fon 1 and Fon 2. It was highlighted the case of Fom 0, in which the radial growths at 15 dpi (23.8–28.5 mm) were significantly lower than those measured at 10 and 5 dpi and that of the control (38.1 mm) (Table 4). On the other hand, isolates Fp, Mon, and Mcr1 did not show significant differences with their controls, comparing both the fermentation days and the extract dose (%). Overall, results seem to indicate that whereas the dose (%) of filtered extract influenced growth inhibition lightly, data coming from the days of fermentation of the extract resulted in significantly different in most cases. Thus, the highest concentration and antifungal activity of secondary metabolites of Ep was obtained after 15 days of agitation in liquid culture (Table 4). Nevertheless, to achieve the highest values of PRGI, an optimization of days of fermentation and dose of extract parameters should be necessary for each pathogen.
For those pathogens in which the radial growth was significantly lower than that controls, it was determined that of the two types of treatment (liquid extract of Ep or dual plate confrontation with Ep), it resulted in being effective to inhibit mycelial growth of these, comparing between treatments (Table 6).
Isolates Fon 0, Fon 2, Fom 0, and Fom 2 showed no significant differences between the two types of biocontrol tests. It was observed that only the radial growth of Fon 1 was significantly minor when cultured in PDA with filtrates, while the rest of pathogens (Fom 1.2w, Fom 1.2y, Mrc2, Fsc R1, Fp, and Fs) significantly decreased their growth in dual plate confrontations. Fast-growing pathogens (i.e., Mon and Mcr1) were not evaluated because of their growth rates in both tests, which did not allow the establishment of the antagonist in the plate to secrete antifungal substances to the medium, preventing fungal growth.
3.4. Bioassay Tests of Biocontrol with Trichoderma Spp in Watermelon and Melon Seedlings
Two strains of Trichoderma (Th and Tl), which showed higher growth inhibitory capacity in dual culture, were selected to test its capacity to reduce disease symptoms in both melon and watermelon plants artificially inoculated with a set of fungal pathogens. Results obtained for each plant host are indicated in Table 7.
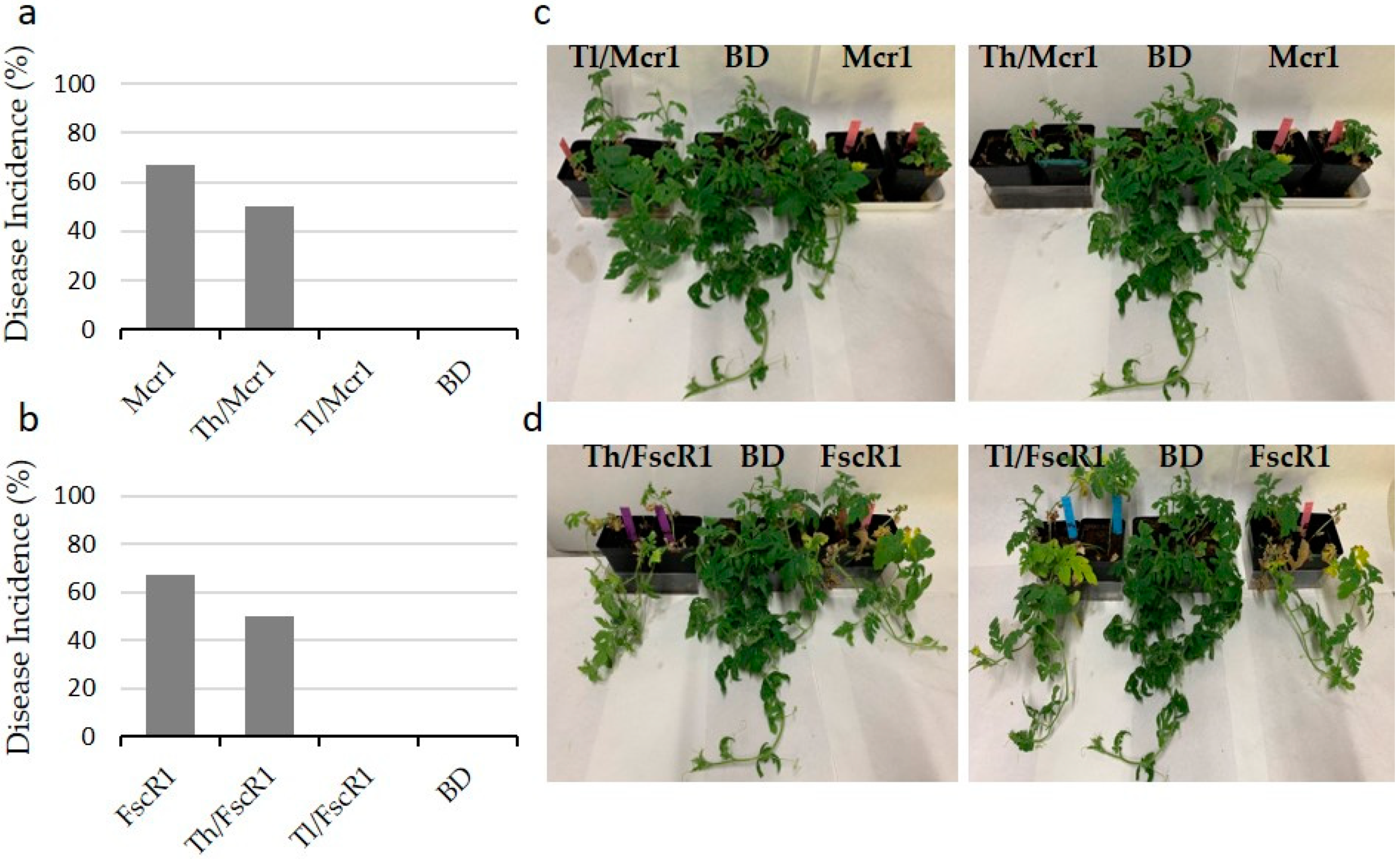
In the case of watermelon plants, there were slight to moderate reductions in the incidence of the symptoms in plants treated with Trichoderma for most of the pathogens. This was especially patent for the case of those plants infected with Mcr1 and Fsc R1 treated with T. lentiforme, which did not show symptomatology or it was practically invaluable (with a disease incidence of 0%), versus the plants inoculated with the pathogens alone, which were highly affected (67%) (Figure 5).
Conversely, the results of disease incidence for melon seedlings did not show great rates of symptom reductions, except in the case the melon plants inoculated with Tl and Fom 2, which did not show symptoms (0%), whereas the plants inoculated with Fom2 showed a disease incidence of 87% (Table 7).
To confirm that the degree of symptomatology was related to the inoculation of antagonist and/or pathogen and ensure that plants were effectively colonized at the root level, all inoculated plants were submitted to Koch’s postulates. In all the pathogen control plants analyzed, the initially inoculated pathogens were recovered. In addition, in the treatments combining the two Trichoderma strains and the different pathogens, both fungi were recovered in all cases.
4. Discussion
4.1. Fungal Isolates
Endophytic fungal species associated with asymptomatic watermelon plants were mostly ascomycete fungi (including mitosporic fungi), followed by a small group of taxa from both the basidiomycetes and zygomycetes, together with some oomycetes (genus Phytophthora). The percentages of isolation of these taxonomic groups were in agreement with those obtained in the majority of studies on the analysis of the endophytic fungal diversity associated with numerous plant hosts (symptomatic or healthy) [28]. It was also observed that identified watermelon endophytes were mostly composed of fungal genera and species not associated specifically with this host, being usually reported as ubiquitous taxa, living on a wide range of plant hosts (e.g., Aureobasidium, Ulocladium, Alternaria, Epicoccum, etc.). An exception was the presence of some specific pathogens of watermelon, including F. oxysporum f. sp. niveum (Fon) (20 isolates), together with five isolates of Fusarium solani and other species of this genus, not specifically related with cucurbits. This seems to suggest that, specifically associated pathogens are commonly present as dormant of facultative parasites in the inner tissues of healthy plants [29]. The existence of such type of non-specialist fungal communities was expected, as it has been reported in other studies on similar hosts, due to the annual cycle of the crop, where the time that the plant remains in the soil does not allow the colonization and infection by specific and complex endophytic communities [28].
Among the isolates obtained, there were identified fungal taxa exhibiting different life strategies including saprophytes, latent pathogens, or antagonists. This last group was the basis to select potential BCAs in the several biocontrol assays performed. Such kind of strategies are commonly designed to exploit the antagonistic potential of microbial communities associated with plants [30]. Characterization of the isolated fungi allowed the selection of potential microbial antagonists with different modes of protection mechanisms according to existing literature, including hyperparasitism, antibiosis, or competition for space or nutrients. In this sense, it was selected an isolate of B. ochroleuca, an ascomycetous species usually employed as a biocontrol agent [31] by its complex lifestyle, which combines a hyperparasitic behavior against other fungi with the production and secretion of mycotoxins. Another microorganism selected was Aureobasidium pullulans, which is currently considered as one of the most common fungal plant endophytes. In this sense, A. pullulans is a ubiquitous microorganism with proven properties as a biocontrol agent, whose mechanisms of action include the production of antifungal enzymes, competition for nutrients, or the induction of defense mechanisms in the plant host [32]. Myrothecium verrucaria is another selected soil-borne ascomycete with antagonism properties. In addition to living on the soil, this fungus is also present as an endophyte in a broad range of hosts, having been commonly cited as a potent natural mycoherbicide due to the production of phytotoxins or as a promising biocontrol tool as an entomopathogenic fungi [33]. Furthermore, M. verrucaria is considered to possess potential antagonistic properties against phytopathogenic fungi, through the production of several types of lytic enzymes [34]. Another selected taxon, Epicoccum purpurascens, is a well-known ubiquitous endophyte, usually inhabiting the tissues of numerous plant species, with proven antagonistic action on certain pathogens due to the production and secretion of antibiotic substances [35]. In addition, selected, the basidiomycete genus Ceratobasidium (binucleate Rhizoctonia) belongs to the so-called Rhizoctonia species complex that includes taxa with potential as biocontrol agents, with a mechanism of action based on competition for nutrients and/or infection sites [36]. Finally, two species of the genus Trichoderma (T. harzianum and T. lentiforme) were also selected, both belonging to a group of highly studied hyperparasitic fungi and widely marketed as biological control agents against a wide range of fungal plant pathogens [37], including soil species [38].
4.2. In Vitro Antagonism
Dual confrontations of the above mentioned BCAs against 14 cucurbit-specific soil-borne pathogens resulted in very different microbial growth reduction effects, depending on the type of microorganism considered. Inhibition effects were more effective when the two Trichoderma strains were confronted in comparison with the rest of antagonists, being their PRGI values sensibly higher for most of the combinations performed. As previously pointed in numerous studies [39], including bioassays against cucurbit pathogens [40,41], the combination of several simultaneous mechanisms of inhibition occurring in Trichoderma (i.e., hyperparasitism, antibiosis or competition for space and nutrients) [42], allows this genus to obtain very notable growth inhibition results in the plate. Concerning in vitro assays, there are numerous studies evaluating the antagonistic potential of genus Trichoderma. Thus, Fernández and Suárez, [43] demonstrated in a research with T. harzianum and Fusarium oxysporum in Passiflora edulis, that the first can slow the growth of the pathogen more than 50%, obtaining PRGI values of 65.91%. In relation to cucurbit diseases, several studies [40,44] have found variable efficacies reducing mycelial growth in plate assays employing different Trichoderma species against several Fusarium species and other specific soil-borne pathogens. In this sense, Boughalleb et al. [40] assayed the antagonistic effects of four Trichoderma species among others, against several cucurbit-associated Fusarium taxa, and found inhibition values not above 50% unlike the results here obtained.
Dual culture assays conducted with both Th and Tl showed high percentages of PRGI, especially against M. cannonballus (Mon). For this pathogen, it was observed that Trichoderma isolates exhibited (in addition to their lytic activity) rapid mycelial growth, preventing the fast-growing rates of Mon once the strain began to grow on the plate. This type of simultaneous effect was already observed by authors like Zhang [45] or more recently Rhouma [46], where these authors reported inhibitory activity of several Trichoderma species (including T. harzianum), combining both parasitism effects with the secretion of antibiotic substances like gliotoxins. Concerning other fast-growing cucurbit pathogens, Aly [47] demonstrated the potential of T. harzianum as a biocontrol agent against M. phaseolina (Mcr), registering PRGI values above 70%.
For the rest of the protective strains assayed, it was observed that PRGI values obtained against most pathogenic strains did not vary sensibly between them for a given pathogen. An exception was observed for M. verrucaria (Mv), which exhibited low rates of PRGI against most of the soil-borne pathogens tested. This could be explained, either by the slow growth rate of this species thus that most of the pathogens invaded the entire plate before M. verrucaria could secrete enzymes to the medium, or due to the nutrient composition used (standard PDA medium), that may not favor their production. Although M. verrucaria is well-known both as a bioherbicide agent [48] or as a common entomopathogenic species mainly due to its ability to secrete hydrolytic enzymes, antibiotics or cyclopeptides [49], the obtained growth inhibition values do not seem to suggest the use of this species as a microbial antagonist against certain soil-borne species.
Relatively fast-growing species like B. ochroleuca, E. purpurascens, and Ceratobasidium sp. exhibited similar discrete PRGI values in general terms, comparing with the other slow-growing antagonist, A. pullulans. Again, it should be mentioned that the growth of this antagonist was usually slower than that of the pathogens. We observed both the secretion of inhibitory substances and the presence of hyperparasitism (presence of hyphal lysis of the pathogen) in dual culture confrontations for B. ochroleuca (Figure 3d). Similar results were reported by Soares de Melo [50], in which a B. ochroleuca strain showed a hyperparasitic mechanism against pathogenic genera like Rhizoctonia or Pythium, and by Rodriguez [51], who indicated the mixed nature of control mechanisms of a Clonostachys rosea (=B. ochroleuca) strain, since it produced either antibiotic substances and hyperparasitized Sclerotinia sclerotiorum in dual culture assays. Ceratobasidium sp. strain assayed showed a capacity to reduce the growth of the pathogens tested, with a high capacity of colonization of the medium in all plate confrontations, and even exhibiting a certain effect of hyphal lysis, like in the confrontation against Mon. Interestingly, control mechanisms described for binucleate Rhizoctonia are usually the competition for nutrients and space (as revealed by the high rates of colonization observed in our study), induced resistance [52], and even growth promotion effects have been reported in cucumber plants treated against damping-off pathogens [53], but no hyperparasitism has been usually reported between hyphae of Ceratobasidium spp. and other plant pathogenic fungi.
Aureobasidium pullulans was able to exert certain growth inhibition in most pathogens tested, despite the absence of hyphal contact observed between the different antagonist/pathogen combinations. In this sense, colony patterns of Fon 1 and Fon 2 isolates were clearly modified and their growth slowed probably by the interaction with compounds secreted to the media by both M. verrucaria and A. pullulans. Some authors [54], hypothesize about a mixed mechanism of antagonism of A. pullulans, including competition for nutrients, induction of defense responses and the production of lytic enzymes. Wachowska and Glowacka [55] reported that when employing A. pullulans against Fusarium culmorum (a winter wheat pathogen) this fungus was able to reduce disease severity up to 21%. Given the fact that A. pullulans exhibits a complex control mechanism [56], inhibition rates observed in our work could be compatible by the presence of such mechanisms in agreement with the mentioned studies, in spite of the slow growing and colonization rates observed.
4.3. Fungicide Effects of Secondary Metabolites from Epicoccum Purpurascens
Concerning the inhibitory effects of culture filtrates of E. purpurascens, results indicated that the dose of the filtered extract (%) did not influence the ability for mycelial growth inhibition, although there were differences with controls. Conversely, the variable days of fermentation employed to obtain filtrates were significantly different in most cases. The highest antifungal activity of secondary metabolites of Ep was obtained after 15 days of liquid culture, probably associated with a stationary phase of the culture. When comparing average data of radial growth of pathogens from both dual confrontations in a plate and incorporating liquid extracts of Ep, results obtained showed that inhibition effects observed in dual confrontations of Ep in a plate were higher (in terms of mycelial growth) than those recorded with liquid culture extracts. It has been reported that the type of control mechanism exerted by Ep is based on antibiosis through certain metabolites diffused to the culture medium [57]. Although PRGI values recorded for Ep were below 50% against all pathogens, in some cases (e.g., Mcr2), values were comparable to those obtained with Trichoderma species.
Epicoccum purpurascens has been largely reported and tested as a biocontrol agent alone or in combination with chemical fungicides, due to its antagonistic properties not only against fungal pathogens [58] including cucurbit pathogens [59], but also controlling other plant parasites like nematodes [60]. Several studies have compared the inhibition effects of antibiotic compounds produced by E. purpurascens from both living dual cultures and filtrates. In this sense, Fávaro [61] found that organic extracts of E. nigrum resulted in more discrete inhibition effects comparing with those observed in plate culture, although the differences observed depended on the range of concentrations employed. In addition, Guerra-Guimaraes [62] also reported that the inhibition effects of E. nigrum on the development of Colletotrichum kahawae were different depending on the incubation times from which the extracts were obtained.
4.4. Plant Protection Assays
Results of biocontrol assays seem to reflect, according to many other similar studies in cucurbits [40,44], the difficulty in obtaining the same rates of growth inhibition of pathogens when treatments with antagonists are scaled to living plants. This protection is difficult to obtain, even when formulations based on Trichoderma are applied preventively, allowing prior root colonization of the antagonist. Just in the same way as in our study (Figure 4), several authors have reported remarkable reductions of disease rates when applying Trichoderma isolates to watermelon and melon plants artificially infected with pathogens like Fusarium solani f. sp. cucurbitae [63] or F. oxysporum f. sp. cucumerinum [44]. Another important effect related to Trichoderma, is the ability of the different species of the genus to promote and stimulate plant growth, widely studied in different plant species including cucurbits [64]. However, our results (data not shown) showed that inoculation of both protective Trichoderma strains only increased root biomass comparing with data of control plants, as previously pointed for this genus in numerous plant hosts, suggesting another relevant feature of the genus from the biotechnological point of view, its potential use as a microbial fertilizer [65].
5. Conclusions
The present study has aimed to explore the capabilities as microbial antagonists of certain endophytic fungal isolates obtained from healthy tissues of watermelon plants, allowing to test different species against some of the most important soil pathogens affecting melon and watermelon in Spain and other Mediterranean areas. Results have highlighted the in vitro antagonistic capacity of the genus Trichoderma above the rest of the antagonists tested. It was also found that the use of liquid culture extracts of the antagonist Epicoccum purpurascens did not offer advantages comparing with the direct plate confrontation of this species with the pathogens. In addition, the two species of Trichoderma selected for testing their abilities in melon and watermelon plants against some of these pathogens, offered discrete symptom reduction rates for most of them, compared with direct plate assays. This type of studies constitute a starting point for the adoption of IPM methods in cucurbits, where the integration of data coming from the screening of natural resistances, a correct selection of varieties and rootstocks or the employment of microbial antagonists can contribute to the control and management of certain soil diseases in these crops in our latitudes.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/10/6/820/s1. Table S1: Evaluation of the differences of each antagonist on each pathogen. Average values of the PRGI (mean ± standard deviation) obtained in each dual plate confrontation for 14 pathogens and 7 antagonists, after 10 days of culture.
Author Contributions
V.G. and A.G.-C. conceived the work. Conceptualization: V.G. and A.G.-C.; methodology, E.A., V.G. and A.G.-C.; software: A.G.-C. and E.A.; validation: V.G. and A.G.-C.; formal analysis: E.A., V.G. and A.G.-C.; investigation: V.G. and A.G.-C.; resources: A.G.-C. and V.G.; data curation: E.A.; writing—original draft preparation: V.G.; writing—review and editing: V.G. and A.G.-C.; visualization: V.G., A.G.-C. and E.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Spanish Ministery of Science, Innovation and Universities-FEDER funds grants AGL2017-85563-C2 (1-R and 2-R) and by the Aragon Government grant for Reference Research Groups, A11-17R-Producción Vegetal Sostenible (PROVESOS).
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. Food and Agricultural Organization of the United Nations: FAOSTAT. 2018. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 1 January 2020).
- Pitrat, M. Melon. In Handbook of Plant Breeding, vol Vegetables I: Asteraceae, Brassicaceae, Chenopodiaceae, and Cucurbitaceae; Prohens, J., Nuez, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 283–315. [Google Scholar]
- Wehner, T.C. Watermelon. In Handbook of Plant Breeding, vol Vegetables I: Asteraceae, Brassicaceae, Chenopodiaceae, and Cucurbitaceae; Prohens, J., Nuez, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 381–418. [Google Scholar]
- Martyn, R.D. Fusarium Wilt of Watermelon: 120 Years of Research. Hortic. Rev. 2014, 42, 349–442. [Google Scholar] [CrossRef]
- Boughalleb, N.; Ben Salem, I.; Beltrán, R.; Vicent, A.; Pérez–Sierra, A.; Abad–Campos, P.; García–Jiménez, J.; Armengol, J. Occurrence of Monosporascus cannonballus in watermelon fields in Tunisia and factors associated with ascospore density in soil. J. Phytopathol. 2010, 158, 137–142. [Google Scholar] [CrossRef]
- Gómez, J.; Serrano, Y.; Pérez, A.; Porcel, E.; Gómez, R.; Aguilar, M.I. Fusarium solani f. sp. cucurbitae, affecting melon in Almería province, Spain. Australas. Plant Dis. Notes 2014, 9, 136. [Google Scholar]
- Armengol, J.; José, C.M.; Moya, M.J.; Sales, R.; Vicent, A.; García–Jiménez, J. Fusarium solani f. sp. cucurbitae race 1, a potential pathogen of grafted watermelon production in Spain. Bull. OEPP 2000, 30, 179–183. [Google Scholar] [CrossRef]
- González, V.; Armengol, J.; Garcés–Claver, A. First report of Fusarium petroliphilum causing fruit rot of butternut squash in Spain. Plant Dis. 2018, 102, 1662. [Google Scholar] [CrossRef]
- Ben Salem, I.; Correia, K.C.; Boughalleb, N.; Michereff, S.J.; León, M.; Abad–Campos, P.; García–Jiménez, J.; Armengol, J. Monosporascus eutypoides as another cause of Monosporascus root rot and vine decline in Tunisia, and evidence that M. cannonballus and M. eutypoides are distinct species. Plant Dis. 2013, 97, 737–743. [Google Scholar] [CrossRef] [Green Version]
- Zitter, T.; Hopkins, D.; Claude, T. Plagas y Enfermedades de las Cucurbitáceas; Mundi–Prensa: Madrid, España, 2004; p. 88. [Google Scholar]
- Petkar, K.; Harris–Shultz, H.; Wang, M.T.; Brewer, L.; Sumabat, P.J. Genetic and phenotypic diversity of Fusarium oxysporum f. sp. niveum populations from watermelon in the southeastern United States. PLoS ONE 2019, 14, e0219821. [Google Scholar] [CrossRef] [Green Version]
- Gómez, J.; Guerra–Sanz, J.M.; Sánchez–Guerrero, M.C.; Serrano, Y.; Melero–Vara, J.M. Crown rot of zucchini squash caused by Fusarium solani f. sp. cucurbitae in Almería Province, Spain. Plant Dis. 2008, 92, 1137. [Google Scholar]
- González, V.; García–Martínez, S.; Ruiz–Martínez, J.J.; Flores–León, A.; Picó, B.; Garcés–Claver, A. First report of Neocosmospora falciformis causing witl and root rot of Muskmelon in Spain. Plant Dis. 2019. [Google Scholar] [CrossRef]
- Shafique, H.A.; Sultana, V.; Ehteshamul–Haque, S.; Athar, M. Management of soil–borne diseases of organic vegetables. J. Plant Prot. Res. 2016, 56, 221–230. [Google Scholar] [CrossRef]
- Giménez, C.; Cabrera, R.; Reina, M.; González–Coloma, A. Fungal Endophytes and their Role in Plant Protection. Curr. Org. Chem. 2007, 11, 707–720. [Google Scholar] [CrossRef]
- Gafni, A.; Calderon, C.E.; Harris, R.; Buxdorf, K.; Dafa–Berger, A.; Zeilinger–Reichert, E.; Levy, M. Biological control of the cucurbit powdery mildew pathogen Podosphaera xanthii by means of the epiphytic fungus Pseudozyma aphidis and parasitism as a mode of action. Front. Plant Sci. 2015, 6, 132. [Google Scholar] [CrossRef] [Green Version]
- Mei, L.; Guang–Shu, M.A.; Lian, H.; Su, X.; Tian, Y.; Huang, W.; Mei, J.; Jiang, X. The effects of Trichoderma on preventing cucumber fusarium wilt and regulating cucumber physiology. J. Integr. Agric. 2019, 18, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Romero, D.; Vicente, A.; Zeriouh, H.; Cazorla, F.; Fernández–Ortuño, D.; Tores, J.; Pérez–García, A. Evaluation of biological control agents for managing cucurbit powdery mildew on greenhouse–grown melon. Plant Pathol. 2007, 56, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Collinge, D.B.; Jørgensen, H.J.L.; Meike, A.C.L.; Manzotti, A.; Ntana, F.; Rojas, E.C.; Jensen, B. Searching for novel fungal biological control agents for plant disease control among endophytes. In Endophytes for a growing World; Hodkinson, T.R., Doohan, F.M., Saunders, M.J., Murphy, B.R., Eds.; Cambrigde Univerisity Press: Cambrigde, UK, 2019; pp. 23–164. [Google Scholar]
- Singh, P.P.; Shin, Y.C.; Park, C.S.; Chung, Y.R. Biological control of fusarium wilt of cucumber by chitinolytic bacteria. Phytopathology 1999, 89, 92–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hartiti, A.; Hichar, A.; Bazdi, O.; El Habchi, S.; Ounine, K. Biocontrol of Fusarium wilt of cucumber (Cucumis sativus L.) by Bacillus sp. World J. Pharm. Life Sci. 2016, 2, 314–322. [Google Scholar]
- Lin–Qi, H.; Niu, Y.-C.; Su, L.; Deng, H.; Lyu, H. The potential of endophytic fungi isolated from cucurbit plants for biocontrol of soilborne fungal diseases of cucumber. Microbiol. Res. 2019, 231, 126369. [Google Scholar] [CrossRef]
- Dennis, C.; Webster, J. Antagonistic properties of species–groups of Trichoderma: III. Hyphal interaction. Trans. Br. Mycol. Soc. 1971, 57, 363–369. [Google Scholar] [CrossRef]
- Ezziyyani, M.; Pérez, S.; Requena, M.; Rubio, L.; Candela, L. Biocontrol de Streptomyces rochei –Ziyani–, de la podredumbre del pimiento (Capsicum annuum L.) causada por Phytophthora capsici. An. Biol. 2004, 26, 69–78. [Google Scholar]
- Worasatit, N.; Sivasithamparam, K.; Ghisalberti, E.L.; Rowland, C. Variation in pyrone production, lytic enzymes and control of rhizoctonia root rot of wheat among single–spore isolates of Trichoderma koningii. Mycol. Res. 1994, 98, 1357–1363. [Google Scholar] [CrossRef]
- Ortiz, A.; Orduz, S. In vitro evaluation of Trichoderma and Gliocladium antagonism against the symbiotic fungus of the leaf–cutting ant Atta cephalotes. Mycopathology 2001, 150, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Martyn, R.D.; Bruton, B.D. An initial survey of the United States for races of Fusarium oxysporum f. sp. niveum. HortScience 1989, 24, 696–698. [Google Scholar]
- Zabalgogeazcoa, I. Review. Fungal endophytes and their interaction with plant pathogens. Span. J. Agric. Res. 2008, 6. [Google Scholar] [CrossRef] [Green Version]
- Pacifico, D.; Squartini, A.; Crucitti, D.; Barizza, E.; Schiavo, F.; Muresu, R.; Carimi, F.; Zottini, M. The Role of the Endophytic Microbiome in the Grapevine Response to Environmental Triggers. Front. Plant Sci. 2019, 10, 1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez–Fernández, R.E.; Sánchez–Ortiz, B.L.; Sandoval–Espinosa, Y.K.; Ulloa–Benítez, A.; Armendáriz–Guillén, B.A.; García–Méndez, M.C.; Macías–Rubalcava, M.L. Hongos endófitos: Fuente potencial de metabolitos secundarios bioactivos con utilidad en agricultura y medicina. Revista Especializada En Ciencias Químico–Biológicas 2013, 16, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Cota, L.V.; Maffia, L.A.; Mizubuti, E.S.G.; Macedo, P.E.F.; Antunes, R.F. Biological control of strawberry gray mold by Clonostachys rosea under field conditions. Biol. Control. 2008, 46, 515–522. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; D’Aquino, S.; Pagnotta, E.; Mari, M. Biocontrol of Monilinia laxa by Aureobasidium pullulans strains: Insights on competition for nutrients and space. Ind. J. Food Microbiol. 2017, 248, 32–38. [Google Scholar] [CrossRef]
- Clarke, T.C.; Shetty, K.G.; Jayachandran, K.; Norland, M.R. Myrothecium verrucaria– a potential biological control agent for the invasive ‘old world climbing fern’ (Lygodium microphyllum). BioControl 2007, 52, 349–411. [Google Scholar] [CrossRef] [Green Version]
- Gupta, J. Efficacy of Biocontrol Agents against Phytophthora infestans on Potato. Int. J. Eng. Sci. Comput. 2016, 6, 2249–2252. [Google Scholar]
- Talontsi, F.M.; Dittrich, B.; Schüffler, A.; Sun, H.; Laatsch, H. Epicoccolides: Antimicrobial and antifungal polyketides from an endophytic fungus Epicoccum sp. associated with Theobroma cacao. Eur. J. Or Chem. 2013, 15, 3174–3180. [Google Scholar] [CrossRef]
- González, V.; Onco, M.P.; Susan, V.R. Biology and systematics of the form genus Rhizoctonia. Span. J. Agric. Res. 2006, 4, 55–79. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.K.; Schmoll, M.; Herrera–Estrella, A.; Upadhyay, R.S.; Druzhinina, I.; Tuohy, M.G. (Eds.) Biotechnology and Biology of Trichoderma; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–650. [Google Scholar] [CrossRef]
- Bastakoti, S.; Belbase, S.; Manandhar, S.; Arjyal, C. Trichoderma species as biocontrol agents against soil–borne fungal pathogens. Nepal J. Biotechnol. 2017, 5, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Brunner, K.; Zeilinger, S.; Ciliento, R.; Woo, S.; Lorito, M.; Kubicek, C.; Mach, R. Improvement of the Fungal Biocontrol Agent Trichoderma atroviride To Enhance both Antagonism and Induction of Plant Systemic Disease Resistance. Appl. Environ. Microbiol. 2005, 71, 3959–3965. [Google Scholar] [CrossRef] [Green Version]
- Boughalleb, N.; Ben Salem, I.; M’Hamdi, M. Evaluation of the efficiency of Trichoderma, Penicillium, and Aspergillus species as biological control agents against four soil–borne fungi of melon and watermelon. Egypt. J. Biol. Pest Col. 2018, 28. [Google Scholar] [CrossRef] [Green Version]
- Kareem, K.; Ugoji, O.; Aboaba, O. Biocontrol of Fusarium wilt of cucumber with Trichoderma longibrachiatum NGJ167 (Rifai). Br. Microbiol. Res. J. 2016, 16, 1–11. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.M. Biocontrol mecanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar]
- Fernández, R.J.; Suárez, C.L. Antagonismo invitro de Trichoderma harzianum Rifai sobre Fusarium oxysporum. Rev. Fac. Nac. Agron. Medellín 2009, 62, 4743–4748. [Google Scholar]
- Srivastava, M. Biological Control of Soil Borne Pathogens (Fusarium oxysporum f.sp. cucumerinum.) of Cucumber (Cucumis sativus) by Trichoderma sp. Int. J. Adv. Res. 2016, 5, 1–9. [Google Scholar] [CrossRef]
- Zhang, J.X.; Bruton, B.D.; Howell, C.R.; Miller, M.E. Potential of Trichoderma virens for biocontrol of root rot and vine decline in Cucumis melo L. caused by Monosporascus cannonballus. Subtrop. Plant Sci. 1999, 51, 29–37. [Google Scholar]
- Rhouma, A.; Ben Salem, I.; M’Hamdi, M.; Boughalleb, N. Antagonistic potential of certain soilborne fungal bioagents against Monosporascus root rot and vine decline of watermelon and promotion of its growth. Nov. Res. Microbiol. J. 2018, 2, 85–100. [Google Scholar] [CrossRef]
- Aly, A.; Abdel–Sattar, M.; Omar, M.; Abd–Elsalam, K. Differential antagonism of Trichoderma sp. against Macrophomina phaseolina. J. Plant Prot. Res. 2007, 47, 91–102. [Google Scholar]
- Lee, H.; Kim, J.-C.; Hong, K.-S.; Kim, C.-J. Evaluation of a Fungal Strain, Myrothecium roridum F0252, as a Bioherbicide Agent. Plant Pathol. J. 2008, 24, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Elkhateeb, W.; Daba, G. Myrothecium as Promising Model for Biotechnological Applications, Potentials and Challenges. J. Sci. Res. 2019, 16, 12126–12131. [Google Scholar] [CrossRef]
- Soares de Melo, I.; Valente, A.; Nessner–Kavamura, V.; Vilela, E.; Nicklin, J. Mycoparasitic nature of Bionectria sp. strain 6.21. J. Plant Prot. Res. 2014, 54. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.; Cabrera, G.; Gozzo, F.; Eberlin, M.; Godeas, A. Clonostachys rosea BAFC3874 as a Sclerotinia sclerotiorum antagonist: Mechanisms involved and potential as a biocontrol agent. J. Appl. Microbiol. 2011, 110, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Olson, H.; Benson, D. Induced systemic resistance and the role of binucleate Rhizoctonia and Trichoderma hamatum 382 in biocontrol of Botrytis blight in geranium. Biol. Control 2007, 42, 233–241. [Google Scholar] [CrossRef]
- Erper, İ.; Turkkan, M. Binucleate Rhizoctonia repens Bernard as a Biocontrol Agent against Damping–off Disease of Cucumber Plants. Eur. J. Plant Sci. Biotechnol. 2018, 7, 58–61. [Google Scholar]
- Lima, G.; Ippolito, A.; Nigro, F.; Salerno, M. Effectiveness of Aureobasidium pullulans and Candida oleophila against postharvest strawberry rots. Postharvest Biol. Technol. 1997, 10, 169–178. [Google Scholar] [CrossRef]
- Wachowska, U.; Glowacka, K. Antagonistic interactions between Aureobasidium pullulans and Fusarium culmorum, a fungal pathogen of winter wheat. BioControl 2014, 59, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Bozoudi, D.; Tsaltas, D. The Multiple and Versatile Roles of Aureobasidium pullulans in the Vitivinicultural Sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, S.; Che, Y.; Liu, X. Epicoccins A–D, epipolythiodioxopiperazines from a Cordyceps–colonizing isolate of Epicoccum nigrum. J. Nat. Prod. 2007, 70, 1522–1525. [Google Scholar] [CrossRef] [PubMed]
- Ogórek, R.; Plaskowska, E. Epicoccum nigrum for biocontrol agents in vitro of plant fungal pathogens. Commun. Agric. Appl. Biol. Sci. 2011, 76, 691–697. [Google Scholar] [PubMed]
- Derbalah, A.; Elkot, G. Cultural filtrates of certain microbial isolates as an alternative to powdery mildew chemical control in cucumbers. J. Plant Prot. Res. 2011, 53, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Arya, R. Effect of culture filtrates of Epicoccum purpurascens and Penicillium vermiculatum grown of different nitrogen sources on hatching and mortality of root knot nematode Meloidogyne incognita. In Proceedings of the National Conference on Conquering Impact of Climate Change on Agriculture through Organic Farming–A Global Perspective; Shweta, V.K., Sing, Eds.; B.R.A. University: Hyderabad, India, 20 February 2011; pp. 101–106. [Google Scholar]
- Fávaro, L.; Sebastianes, F.; Araújo, W. Epicoccum nigrum P16, a Sugarcane Endophyte, Produces Antifungal Compounds and Induces Root Growth. PLoS ONE 2012, 7, e36826. [Google Scholar] [CrossRef]
- Guerra–Guimarães, L.; Azinheira, G.; Martins, A.C.; Silva, M.C.; Gichuru, E.K.; Varzea, V.; Bertrand, B. Antagonistic interaction between Epicoccum nigrum and Colletotrichum kahawae, the causal agent of coffee berry disease. In Proceedings of the 21st International Conference on Coffee Science, Montpellier, France, 11 September 2006; ASIC: Allenwinden, Switzerland, 2007; pp. 1284–1290. [Google Scholar]
- Bernal–Vicente, A.; Ros, M.; Pascual, J.A. Increased effectiveness of the Trichoderma harzianum isolate T–78 against Fusarium wilt on melon plants under nursery conditions. J. Sci. Food Agric. 2009, 89, 827–833. [Google Scholar] [CrossRef]
- Marín–Guirao, J.I.; Rodríguez–Romera, P.; Lupión–Rodríguez, B.; Camacho–Ferre, F.; Tello–Marquina, J. Effect of Trichoderma on horticultural seedlings growth promotion depending on inoculum and substrate type. J. Appl. Microbiol. 2016, 121, 1095–1102. [Google Scholar] [CrossRef]
- Kamal, R.; Athisayam, V.; Gusain, S.Y.; Kumar, V. Trichoderma: A Most Common Biofertilizer with Multiple Roles in Agriculture. Biomed. J. Sci. Tech. Res. 2018, 4. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Endophytic fungi (genera and/or species) obtained in the present study in each of the localities sampled. Bars indicate the number of strains obtained per operational taxonomic unit (OTU). Green and brown bars indicate the type of tissue from which they were isolated, stem or root, respectively. *Asterisks indicate that the taxonomic determinations were made from internal transcribed spacer (ITS) sequences; (a) Fungi isolated from a commercial field of Peñaflor; (b) Fungi isolated from a commercial field of Alfamén.
Figure 1.
Endophytic fungi (genera and/or species) obtained in the present study in each of the localities sampled. Bars indicate the number of strains obtained per operational taxonomic unit (OTU). Green and brown bars indicate the type of tissue from which they were isolated, stem or root, respectively. *Asterisks indicate that the taxonomic determinations were made from internal transcribed spacer (ITS) sequences; (a) Fungi isolated from a commercial field of Peñaflor; (b) Fungi isolated from a commercial field of Alfamén.

Figure 2.
Growth inhibition effects of Trichoderma harzianum (Th) against 14 selected pathogens. From left to right there is a pathogen control plate followed by the dual culture plate (pathogen/T. harzianum).
Figure 2.
Growth inhibition effects of Trichoderma harzianum (Th) against 14 selected pathogens. From left to right there is a pathogen control plate followed by the dual culture plate (pathogen/T. harzianum).

Figure 3.
Effect of inhibition of the rest of antagonists on the growth of several pathogens. From left to right there are control plates of both antagonists and pathogens followed by the dual culture plate. (a): E. purpurascens (Ep)/M. phaseolina (Mrc2). (b) M. verrucaria (Mv)/Fon 2. (c): A. pullulans (Ap)/Fon 1. (d): B. ochroleuca (Bio)/Fom 1.2w. (e): Binucleate Rhizoctonia (Rb)/M. cannonballus (Mon).
Figure 3.
Effect of inhibition of the rest of antagonists on the growth of several pathogens. From left to right there are control plates of both antagonists and pathogens followed by the dual culture plate. (a): E. purpurascens (Ep)/M. phaseolina (Mrc2). (b) M. verrucaria (Mv)/Fon 2. (c): A. pullulans (Ap)/Fon 1. (d): B. ochroleuca (Bio)/Fom 1.2w. (e): Binucleate Rhizoctonia (Rb)/M. cannonballus (Mon).

Figure 4.
Interaction of Trichoderma spp. with pathogenic species in dual plate confrontation. (a,b): Papillae-like structures. (c,d): Cell lysis. (e): Appressorium formation. (f–h): Coil hyphae. Bars = 10 µm.
Figure 4.
Interaction of Trichoderma spp. with pathogenic species in dual plate confrontation. (a,b): Papillae-like structures. (c,d): Cell lysis. (e): Appressorium formation. (f–h): Coil hyphae. Bars = 10 µm.

Figure 5.
Comparison of protection effects of Trichoderma strains, T. harzianum (Th) and T. lentiforme (Tl), in ‘Black Diamond’ watermelon plants (BD) against the cucurbit pathogens Macrophomina phaseolina (Mcr1) and Fusarium solani f. sp. cucurbitaceae race 1 (Fsc R1) when T. harzianum or T. lentiforme and these pathogens were inoculated simultaneously and individually in climatic chamber bioassay. (a,b): Disease incidence (%) of plants inoculation with Mcr1 or Fsc R1 alone or in combination with Th or Tl. (c,d): Response of the plants inoculated with Mcr1 or FscR1 alone or in combination with Th or Tl. BD represented watermelon plants without inoculate.
Figure 5.
Comparison of protection effects of Trichoderma strains, T. harzianum (Th) and T. lentiforme (Tl), in ‘Black Diamond’ watermelon plants (BD) against the cucurbit pathogens Macrophomina phaseolina (Mcr1) and Fusarium solani f. sp. cucurbitaceae race 1 (Fsc R1) when T. harzianum or T. lentiforme and these pathogens were inoculated simultaneously and individually in climatic chamber bioassay. (a,b): Disease incidence (%) of plants inoculation with Mcr1 or Fsc R1 alone or in combination with Th or Tl. (c,d): Response of the plants inoculated with Mcr1 or FscR1 alone or in combination with Th or Tl. BD represented watermelon plants without inoculate.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Pathogenic isolates and potential microbial antagonists used in this study for their evaluation in dual plate cultures.
Table 1.
Pathogenic isolates and potential microbial antagonists used in this study for their evaluation in dual plate cultures.
| Pathogens | Plant Host | Abbreviation |
|---|---|---|
| Fusarium oxysporum f. sp. niveum (race 0) | Citrullus lanatus | Fon 0 |
| Fusarium oxysporum f. sp. niveum (race 1) | Citrullus lanatus | Fon 1 |
| Fusarium oxysporum f. sp. niveum (race 2) | Citrullus lanatus | Fon 2 |
| Fusarium oxysporum f. sp. melonis (race 0) | Cucumis melo | Fom 0 |
| Fusarium oxysporum f. sp. melonis (race 1) | Cucumis melo | Fom 1 |
| Fusarium oxysporum f. sp. melonis (race 2) | Cucumis melo | Fom 2 |
| Fusarium oxysporum f. sp. melonis (race 1.2y) | Cucumis melo | Fom 1.2 y |
| Fusarium oxysporum f. sp. melonis (race 1.2w) | Cucumis melo | Fom 1.2 w |
| Monosporascus cannonballus isolate | Citrullus lanatus | Mon |
| Macrophomina phaseolina isolate 1 | Cucumis melo | Mcr1 |
| Macrophomina phaseolina isolate 2 | Cucumis melo | Mcr2 |
| Fusarium solani f. sp. cucurbitae race 1 (=Neocosmospora cucurbitae) | Cucurbita moschata | Fsc R1 |
| Fusarium solani f. sp. cucurbitae race 2 (=Neocosmospora petroliphila) | Cucurbita moschata | Fp |
| Fusarium solani (=Neocosmospora keratoplastica) | Cucumis melo | Fs |
| Antagonists | Source | Abbreviation |
| Epicoccum purpurascens Ehrenb. | Citrullus lanatus | Ep |
| Bionectria ochroleuca Schroers & Samuels (=Clonostachys rosea (Link) Schroers, Samuels, Seifert & W. Gams) | Citrullus lanatus | Bo |
| Myrothecium verrucaria (Alb. & Schwein.) Ditmar | Citrullus lanatus | Mv |
| Aureobasidium pullulans (de Bary) G. Arnaud | Citrullus lanatus | Ap |
| Ceratobasidium sp. (binucleate Rhizoctonia) | Citrullus lanatus | Rb |
| Trichoderma harzianum Rifai | Citrullus lanatus | Th |
| Trichoderma lentiforme (Rehm) P. Chaverri, Samuels & F.B. Rocha | Citrullus lanatus | Tl |
Table 2.
Average values of the percentage of radial growth inhibition (PRGI) (mean ± standard deviation) obtained in each dual plate confrontation for 14 pathogens and 7 antagonists, after 10 days of culture.
Table 2.
Average values of the percentage of radial growth inhibition (PRGI) (mean ± standard deviation) obtained in each dual plate confrontation for 14 pathogens and 7 antagonists, after 10 days of culture.
| Trichoderma harzianum | Trichoderma lentiforme | Epicoccum purpurascens | Aureobasidium pullulans | Myrothecium verrucaria | Ceratobasidium sp. | Bionectria ochroleuca | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fon 0 | 56.3 | de | ± | 4.5 | 44.3 | fg | ± | 11.3 | 29.6 | abcd | ± | 1.8 | 23.3 | cde | ± | 0.8 | 15.3 | bc | ± | 1.4 | 45.6 | ab | ± | 6.9 | 24.1 | cd | ± | 4.3 |
| Fon 1 | 56.7 | de | ± | 2.1 | 60.6 | cd | ± | 0.8 | 29.0 | bcd | ± | 4.2 | 42.9 | a | ± | 2.2 | 4.0 | cd | ± | 4.5 | 39.9 | bc | ± | 4.6 | 19.8 | d | ± | 1.7 |
| Fon 2 | 39.2 | f | ± | 1.0 | 51.0 | efgh | ± | 1.0 | 4.9 | fg | ± | 6.0 | 7.3 | fg | ± | 5.0 | 3.9 | cd | ± | 4.0 | 21.0 | cd | ± | 5.3 | 7.7 | e | ± | 2.0 |
| Fom 0 | 67.6 | bcd | ± | 1.3 | 65.6 | bcd | ± | 1.5 | 35.9 | ab | ± | 2.7 | 26.7 | bcde | ± | 3.6 | 10.4 | bc | ± | 2.6 | 33.6 | bc | ± | 10.1 | 30.0 | abc | ± | 1.6 |
| Fom 1 | 40.9 | f | ± | 9.8 | 42.4 | h | ± | 6.1 | 16.3 | ef | ± | 0.9 | 20.0 | def | ± | 6.0 | 7.7 | bcd | ± | 4.7 | 37.6 | bc | ± | 9.1 | 32.1 | ab | ± | 2.5 |
| Fom 2 | 47.1 | ef | ± | 2.2 | 49.4 | efgh | ± | 6.6 | 20.2 | de | ± | 3.5 | 40.2 | ab | ± | 6.1 | 27.2 | a | ± | 5.7 | 29.9 | bc | ± | 8.7 | 30.5 | abc | ± | 1.1 |
| Fom 1.2y | 56.2 | de | ± | 1.3 | 47.1 | fgh | ± | 2.5 | 22.3 | cde | ± | 2.2 | 34.6 | abc | ± | 9.6 | 8.6 | bcd | ± | 3.5 | 32.2 | bc | ± | 6.3 | 21.7 | d | ± | 1.4 |
| Fom 1.2w | 60.3 | cd | ± | 0.6 | 59.9 | def | ± | 0.9 | 33.9 | abc | ± | 1.2 | 29.4 | abcd | ± | 2.3 | 14.2 | bc | ± | 3.4 | 36.7 | bc | ± | 14.1 | 32.9 | a | ± | 2.1 |
| Mon | 93.4 | a | ± | 0.7 | 93.3 | a | ± | 0.7 | 10.1 | efg | ± | 11.6 | x 1 | 3.7 | cd | ± | 6.5 | 64.8 | a | ± | 7.6 | x 1 | ||||||
| Mcr1 | 70.2 | bc | ± | 6.3 | 56.1 | defg | ± | 2.4 | x 1 | x 1 | x 1 | 4.3 | d | ± | 7.4 | x 1 | ||||||||||||
| Mcr2 | 72.2 | b | ± | 1.5 | 76.1 | b | ± | 5.1 | 41.3 | a | ± | 2.5 | 19.4 | def | ± | 3.8 | 16.4 | ab | ± | 1.8 | 31.0 | bc | ± | 9.1 | 7.8 | e | ± | 4.7 |
| Fsc R1 | 74.1 | b | ± | 1.2 | 73.4 | bc | ± | 1.6 | 16.6 | ef | ± | 3.5 | 29.3 | abcd | ± | 8.4 | 14.8 | bc | ± | 2.4 | 4.3 | d | ± | 1.3 | 20.1 | d | ± | 4.4 |
| Fp | 70.4 | bc | ± | 1.3 | 65.1 | bcd | ± | 3.6 | 31.1 | abcd | ± | 1.3 | 24.5 | cde | ± | 4.8 | 19.7 | ab | ± | 1.9 | 23.9 | bcd | ± | 4.2 | 25.5 | bcd | ± | 0.5 |
| Fs | 66.1 | bcd | ± | 3.6 | 68.2 | bcd | ± | 1.3 | 30.6 | abcd | ± | 1.7 | 13.8 | fg | ± | 1.9 | 15.0 | bc | ± | 4.7 | 44.9 | ab | ± | 2.6 | 25.3 | bcd | ± | 2.0 |
For each column, means followed with different letters are significantly different (Tukey, p ≤ 0.05). x1: PRGI values could not be calculated because the radial length of the pathogen in dual culture was greater than radial length in control plate. Columns are alternately shaded for easy reading
Table 3.
Degree of invasion (mean ± SD of three replicates) of the 14 pathogen colonies caused by Th and Tl in dual culture, 10 days after inoculation. A visual scale with values from 0 to 4 described by Ortiz & Orduz [26] was used, in which: 0, no invasion of pathogen colony; 1, invasion of 25% of the pathogen colony surface area; 2, invasion of 50% of the pathogen colony surface area; 3, invasion of 100% of the pathogen colony surface area; 4 invasion of 100% of the pathogen colony surface area and sporulation.
Table 3.
Degree of invasion (mean ± SD of three replicates) of the 14 pathogen colonies caused by Th and Tl in dual culture, 10 days after inoculation. A visual scale with values from 0 to 4 described by Ortiz & Orduz [26] was used, in which: 0, no invasion of pathogen colony; 1, invasion of 25% of the pathogen colony surface area; 2, invasion of 50% of the pathogen colony surface area; 3, invasion of 100% of the pathogen colony surface area; 4 invasion of 100% of the pathogen colony surface area and sporulation.
| Combination Pathogen-Th | Colonization Ability | Combination Pathogen-Tl | Colonization Ability |
|---|---|---|---|
| Fon 0/Th | 1.0 ± 0.0 | Fon 0/Tl | 1.3 ± 0.6 |
| Fon 1/Th | 1.0 ± 0.0 | Fon 1/Tl | 1.0 ± 0.0 |
| Fon 2/Th | 0.7 ± 0.6 | Fon 2/Tl | 1.7 ± 1.6 |
| Fom 0/Th | 1.0 ± 0.0 | Fom 0/Tl | 1.0 ± 0.0 |
| Fom 1/Th | 0.0 ± 0.0 | Fom 1/Tl | 0.0 ± 0.0 |
| Fom 2/Th | 0.0 ± 0.0 | Fom 2/Tl | 0.0 ± 0.0 |
| Fom 1.2 y/Th | 0.7 ± 0.6 | Fom 1.2 y/Tl | 1.0 ± 0.0 |
| Fom 1.2 w/Th | 2.0 ± 0.0 | Fom 1.2 w/Tl | 1.0 ± 0.0 |
| Mon/Th | 4.0 ± 0.0 | Mon/Tl | 4.0 ± 0.0 |
| Mcr1/Th | 3.0 ± 0.0 | Mcr1/Tl | 2.0 ± 0.0 |
| Mcr2/Th | 4.0 ± 0.0 | Mcr2/Tl | 3.0 ± 0.0 |
| Fsc R1/Th | 0.0 ± 0.0 | Fsc R1/Tl | 0.3 ± 0.6 |
| Fp/Th | 2.0 ± 0.0 | Fp/Tl | 1.0 ± 0.0 |
| Fs/Th | 2.0 ± 1.7 | Fs/Tl | 1.0 ± 0.0 |
Table 4.
Radial growth (in mm) of pathogens (mean value ± SD of three replicates) inoculated in a potato dextrose agar (PDA) medium containing 0.5, 1, and 2% extract of metabolites filtered from liquid cultures of Epicoccum purpurascens at 5, 10, and 15 dpi.
Table 4.
Radial growth (in mm) of pathogens (mean value ± SD of three replicates) inoculated in a potato dextrose agar (PDA) medium containing 0.5, 1, and 2% extract of metabolites filtered from liquid cultures of Epicoccum purpurascens at 5, 10, and 15 dpi.
| Pathogen | Control * | Fermentation Days of Epicoccum purpurascens | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 5 dpi | 10 dpi | 15 dpi | ||||||||
| Extract Doses | Extract Doses | Extract Doses | ||||||||
| 0.5% | 1% | 2% | 0.5% | 1% | 2% | 0.5% | 1% | 2% | ||
| Fon 0 | 41.8 ± 2.3 | 38.3 ± 0.9 | 39.3 ± 1.0 | 39.3 ± 1.3 | 36.7 ± 1.1 | 36.5 ± 0.5 | 38.7 ± 0.8 | 31.0 ± 2.8 | 30.0 ± 2.6 | 29.8 ± 1.5 |
| Fon 1 | 34.0 ± 1.2 | 31.9 ± 0.7 | 27.6 ± 6.3 | 26.7 ± 0.4 | 35.4 ± 1.1 | 31.5 ± 2.6 | 29.4 ± 2.9 | 23.8 ± 4.1 | 20.5 ± 1.3 | 18.0 ± 0.8 |
| Fon 2 | 29.1 ± 2.7 | 32.5 ± 1.7 | 33.4 ± 0.9 | 33.8 ± 0.7 | 28.7 ± 4.9 | 32.5 ± 1.8 | 29.3 ± 1.1 | 19.6 ± 1.5 | 20.0 ± 3.24 | 18.4 ± 2.3 |
| Fom 0 | 38.1 ± 0.9 | 36.9 ± 1.4 | 35.4 ± 1.0 | 30.5 ± 0.6 | 27.1 ± 5.2 | 34.5 ± 1.4 | 25.4 ± 5. 6 | 23.8 ± 2.7 | 25.5 ± 0.9 | 28.5 ± 0.5 |
| Fom 1 | 40.3 ± 0.3 | 41.8 ± 1.6 | 40.6 ± 0.5 | 39.5 ± 2.5 | 39.6 ± 1.2 | 39.1 ± 1.5 | 39.0 ± 1.7 | 37.9 ± 1.2 | 40.2 ± 0.3 | 38.1 ± 0.9 |
| Fom 2 | 30.7 ± 5.0 | 25.5 ± 2.8 | 23.4 ± 2.0 | 23.0 ± 0.7 | 27.7 ± 1.2 | 23.5 ± 1.8 | 23.6 ± 1.7 | 17.4 ± 0.2 | 23.6 ± 4.6 | 28.0 ± 4.1 |
| Fom 1.2y | 41.4 ± 0.4 | 40.5 ± 0.9 | 36.9 ± 0.6 | 36.4 ± 1.6 | 37.5 ± 1.7 | 40.2 ± 0.9 | 39.2 ± 0.8 | 39.0 ± 0.3 | 38.9 ± 1.2 | 35.0 ± 2.8 |
| Fom 1.2w | 39.8 ± 0.8 | 33.0 ± 6.7 | 36.2 ± 2.0 | 38.0 ± 1.0 | 39.0 ± 1.6 | 36.5 ± 4.2 | 36.8 ± 0.7 | 33.8 ± 3.5 | 32.0 ± 2.6 | 25.3 ± 5.4 |
| Mon | 43.4 ± 0.2 | 43.4 ± 0.3 | 43.3 ± 0.0 | 43.3 ± 0.0 | 43.3 ± 0.0 | 43.3 ± 0.0 | 43.3 ± 0.0 | 43.3 ± 0.0 | 43.2 ± 0.0 | 43.2 ± 0.1 |
| Mcr1 | 43.3 ± 0.2 | 42.9 ± 0.6 | 43.5 ± 0.6 | 43.2 ± 0.3 | 43.1 ± 0.3 | 43.2 ± 0.1 | 43.2 ± 0.2 | 43.2 ±0.2 | 42.8 ± 0.4 | 43.1 ± 0.1 |
| Mcr2 | 42.4 ± 1.0 | 36.3 ± 3.3 | 39.5 ± 2.1 | 38.7 ± 2.0 | 42.2 ± 0.7 | 39.8 ± 1.8 | 40.7 ± 0.4 | 34.9 ± 1.0 | 32.2 ± 2.9 | 33.7 ± 1.4 |
| Fsc R1 | 34.1 ± 2.0 | 34.6 ± 0.3 | 33.5 ± 0.3 | 32.1 ± 0.3 | 32.9 ± 0.6 | 32.8 ± 0.1 | 31.8 ± 0.5 | 33.6 ± 0.5 | 32.6 ± 0.1 | 34.1 ± 1.3 |
| Fp | 41.7 ± 1.1 | 38.9 ± 0.6 | 36.2 ± 1.5 | 37.1 ± 0.8 | 40.3 ± 0.3 | 39.7 ± 0.4 | 38.2 ± 0.6 | 33.3 ± 0.3 | 33.2 ± 0.2 | 32.2 ± 0.1 |
| Fs | 43.8 ± 0.0 | 43.3 ± 1.7 | 42.6 ± 1.4 | 42.2 ± 0.1 | 43.3 ± 0.4 | 43.2 ± 0.2 | 43.0 ± 0.2 | 38.9 ± 0.1 | 38.2 ± 0.4 | 38.7 ± 0.2 |
* Control represents pathogenic fungus culture in PDA plates without E. purpurascens filtrate.
Table 5.
Analysis of variance (ANOVA) results for radial growth of the pathogenic fungi cultured in a PDA medium containing 0.5, 1%, and 2% extract of metabolites filtered (dose) from liquid cultures of Epicoccum purpurascens at 5, 10, and 15 dpi (days).
Table 5.
Analysis of variance (ANOVA) results for radial growth of the pathogenic fungi cultured in a PDA medium containing 0.5, 1%, and 2% extract of metabolites filtered (dose) from liquid cultures of Epicoccum purpurascens at 5, 10, and 15 dpi (days).
| Source of Variation | Sum of Squares | df | Mean Square | F | p |
|---|---|---|---|---|---|
| Days | 1685.159 | 2 | 842.579 | 195.100 | 0.000 |
| Dose | 74.032 | 2 | 37.016 | 8.571 | 0.000 |
| Pathogen | 10144.267 | 13 | 780.328 | 180.685 | 0.000 |
| Days*Dose | 5.954 | 4 | 1.489 | 0.345 | 0.848 |
| Days*Pathogen | 1604.482 | 26 | 61.711 | 14.289 | 0.000 |
| Dose*Pathogen | 228.520 | 26 | 8.789 | 2.035 | 0.003 |
| Days *Dose*Pathogen | 749.974 | 52 | 14.423 | 3.340 | 0.000 |
| Error | 1153.096 | 267 | 4.319 | 1153.096 | 267 |
df, degrees of freedom; F, F-ratio; p, level of significance (p < 0.05).
Table 6.
Comparison of radial growth (mean value ± SD) between dual culture and secondary metabolites extract inhibition assays with E. purpurascens.
Table 6.
Comparison of radial growth (mean value ± SD) between dual culture and secondary metabolites extract inhibition assays with E. purpurascens.
| Pathogen | Radial Growth (in mm) of Pathogens Caused by Epicoccum purpurascens | ||||||
|---|---|---|---|---|---|---|---|
| Dual Culture | Liquid Extract | p-Value | |||||
| Fon 0 | 28.0 | ± | 0.7 | 30.3 | ± | 2.1 | 0.1105 |
| Fon 1 | 25.6 | ± | 1.5 | 17.9 | ± | 0.8 | 0.0016 |
| Fon 2 | 19.6 | ± | 1.1 | 19.4 | ± | 2.3 | 0.8456 |
| Fom 0 | 24.6 | ± | 1.0 | 25.9 | ± | 2.5 | 0.3932 |
| Fom 2 | 22.5 | ± | 0.9 | 23.9 | ± | 3.7 | 0.5189 |
| Fom 1.2 y | 30.9 | ± | 0.9 | 38.3 | ± | 2.1 | 0.0001 |
| Fom 1.2 w | 27.0 | ± | 0.5 | 35.7 | ± | 4.1 | 0.0058 |
| Mcr2 | 20.8 | ± | 0.9 | 33.6 | ± | 2.1 | 0.0001 |
| Fsc R1 | 27.1 | ± | 1.1 | 38.2 | ± | 0.6 | 0.0001 |
| Fp | 25.2 | ± | 0.5 | 32.2 | ± | 0.1 | 0.0001 |
| Fs | 28.1 | ± | 0.7 | 38.6 | ± | 0.4 | 0.0001 |
t-Test values indicate the level of significance (p-value) for each pathogen. Significant levels p < 0.05 are indicated in bold.
Table 7.
Degree of symptomatology of ‘Black Diamond’ watermelon and ‘Charentais T’ melon plants inoculated with Trichoderma harzianum (Th) and Trichoderma lentiforme (Tl) alone and in combination (Tl + pathogen and Th + pathogen) with Fon 0, Fon 1, Fon 2, Mon, Mcr1, Fsc R1, and Fp (watermelon), and Fom 0, Fom 1, Fom 2, and Fom 1.2 y (melon), respectively. Degree of symptomatology was evaluated using a visual scale from 1 to 4, in the following way: 1 = healthy plants; 2 = stunted growth and damage of up to 50%; 3 = presence of foliar lesions, yellowing, wilting and root damage; 4 = dead plants. Disease incidence (%) is indicated in parentheses.
Table 7.
Degree of symptomatology of ‘Black Diamond’ watermelon and ‘Charentais T’ melon plants inoculated with Trichoderma harzianum (Th) and Trichoderma lentiforme (Tl) alone and in combination (Tl + pathogen and Th + pathogen) with Fon 0, Fon 1, Fon 2, Mon, Mcr1, Fsc R1, and Fp (watermelon), and Fom 0, Fom 1, Fom 2, and Fom 1.2 y (melon), respectively. Degree of symptomatology was evaluated using a visual scale from 1 to 4, in the following way: 1 = healthy plants; 2 = stunted growth and damage of up to 50%; 3 = presence of foliar lesions, yellowing, wilting and root damage; 4 = dead plants. Disease incidence (%) is indicated in parentheses.
| Black Diamond Watermelon Plants a | |||
| Plants Non-Inoculated | |||
| 1.00 ± 0.00 (0%) | |||
| Plants Inoculated with Antagonists | |||
| Th | Tl | ||
| 1.00 ± 0.00 (0%) | 1.50 ± 0.54 (0%) | ||
| Plants Inoculated with Antagonist + Pathogen and Only Pathogen | |||
| Tl + Pathogen | Th + Pathogen | Pathogen | |
| Fon 0 | 1.50 ± 0.55 (0%) | 1.50 ± 1.22 (17%) | 2.50 ± 0.83 (33%) |
| Fon 1 | 2.66 ± 1.50 (50%) | 3.16 ± 0.98 (67%) | 2.50 ± 0.83 (33%) |
| Fon 2 | 1.50 ± 0.55 (0%) | 1.83 ± 0.98 (33%) | 2.00 ± 0.00 (0%) |
| Mon | 2.17 ± 1.17 (33%) | 1.50 ± 1.22 (17%) | 2.17 ± 0.41 (33%) |
| Mcr1 | 1.00 ± 0.00 (0%) | 2.83 ± 0.98 (50%) | 3.17 ± 1.33 (67%) |
| FscR1 | 1.17 ± 0.41 (0%) | 2.50 ± 1.22 (50%) | 3.00 ± 0.89 (67%) |
| Fp | 1.00 ± 0.00 (0%) | 2.50 ± 1.05 (50%) | 2.17 ± 1.17 (33%) |
| Charentais T Melon Plants a | |||
| Plants Non-Inoculated | |||
| 1.00 ± 0.00 (0%) | |||
| Plants Inoculated with Antagonists | |||
| Th | Tl | ||
| 1.00 ± 0.00 (0%) | 1.00 ± 0.00 (0%) | ||
| Plants Inoculated with Antagonist + Pathogen and Only Pathogen | |||
| Tl + Pathogen | Th + Pathogen | Pathogen | |
| Fom 0 | 1.00 ± 0.00 (0%) | 1.00 ± 0.00 (0%) | 1.50 ± 0.55 (0%) |
| Fom 1 | 3.66 ± 0.82 (100%) | 3.33 ± 0.82 (83%) | 3.50 ± 0.55 (100%) |
| Fom 2 | 2.00 ± 0.00 (0%) | 1.00 ± 0.00 (0%) | 3.33 ± 1.03 (67%) |
| Fom1.2 | 3.50 ± 0.84 (83%) | 3.67 ± 0.82 (83%) | 3.67 ± 0.52 (100%) |
a Data are expressed as means ± SE from experiments with 2 replicates of 3 plants each.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
González, V.; Armijos, E.; Garcés-Claver, A. Fungal Endophytes as Biocontrol Agents against the Main Soil-Borne Diseases of Melon and Watermelon in Spain. Agronomy 2020, 10, 820. https://doi.org/10.3390/agronomy10060820
AMA Style
González V, Armijos E, Garcés-Claver A. Fungal Endophytes as Biocontrol Agents against the Main Soil-Borne Diseases of Melon and Watermelon in Spain. Agronomy. 2020; 10(6):820. https://doi.org/10.3390/agronomy10060820
Chicago/Turabian StyleGonzález, Vicente, Eugenia Armijos, and Ana Garcés-Claver. 2020. "Fungal Endophytes as Biocontrol Agents against the Main Soil-Borne Diseases of Melon and Watermelon in Spain" Agronomy 10, no. 6: 820. https://doi.org/10.3390/agronomy10060820
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.