Genome-Wide Transcriptome and Metabolome Analyses Provide Novel Insights and Suggest a Sex-Specific Response to Heat Stress in Pigs

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Heat Stress Experimental Setup
2.3. RNA Isolation and Sequencing
2.4. Sample Preperation and Metabolite Analysis with 1H-NMR
2.5. Data Analysis
2.5.1. RNA-Seq Data
2.5.2. Metabolome Data
2.5.3. Pathway Enrichment Analysis of Differentially-Enriched Metabolites
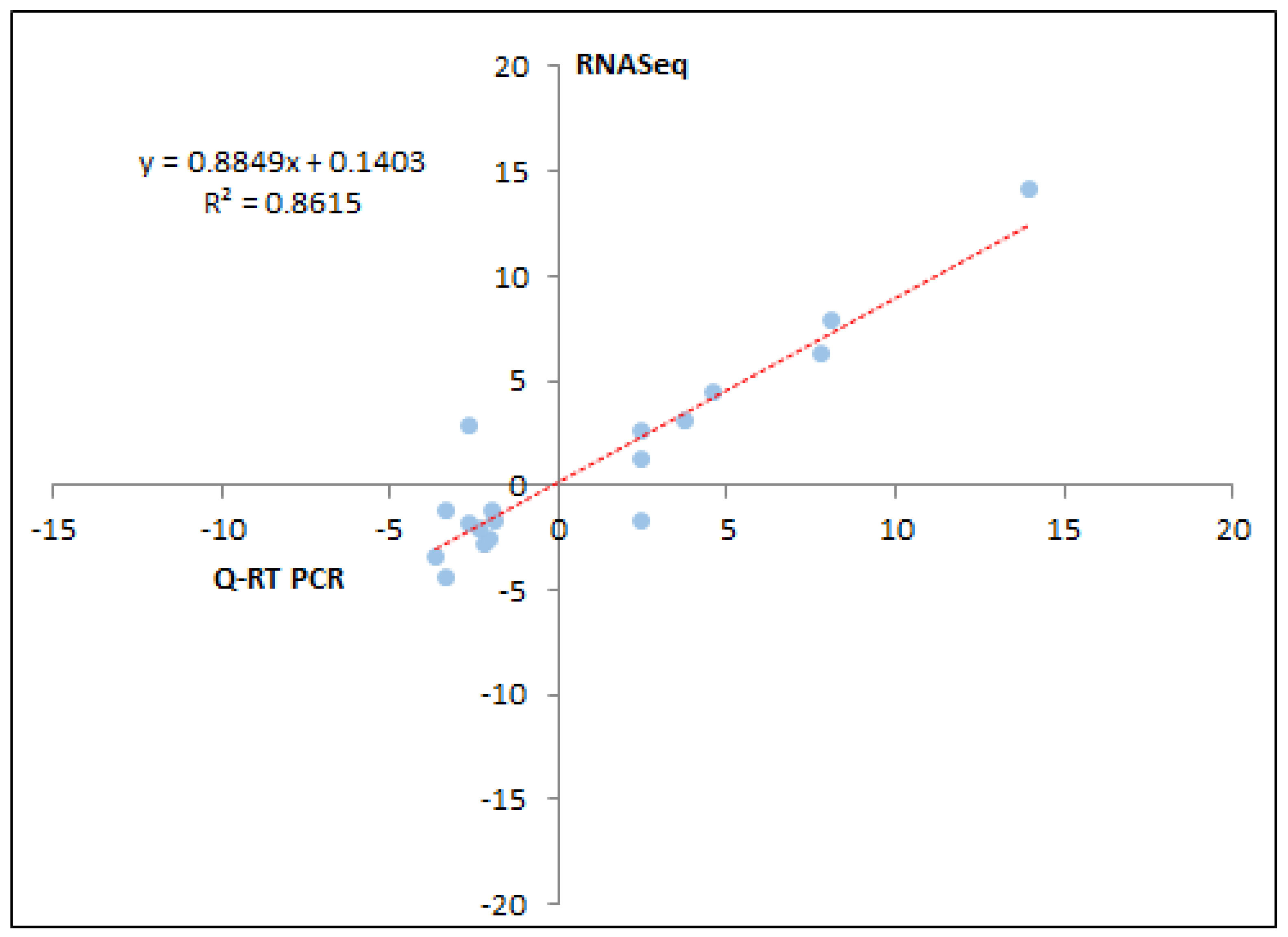
2.6. Real-Time PCR Validation
3. Results
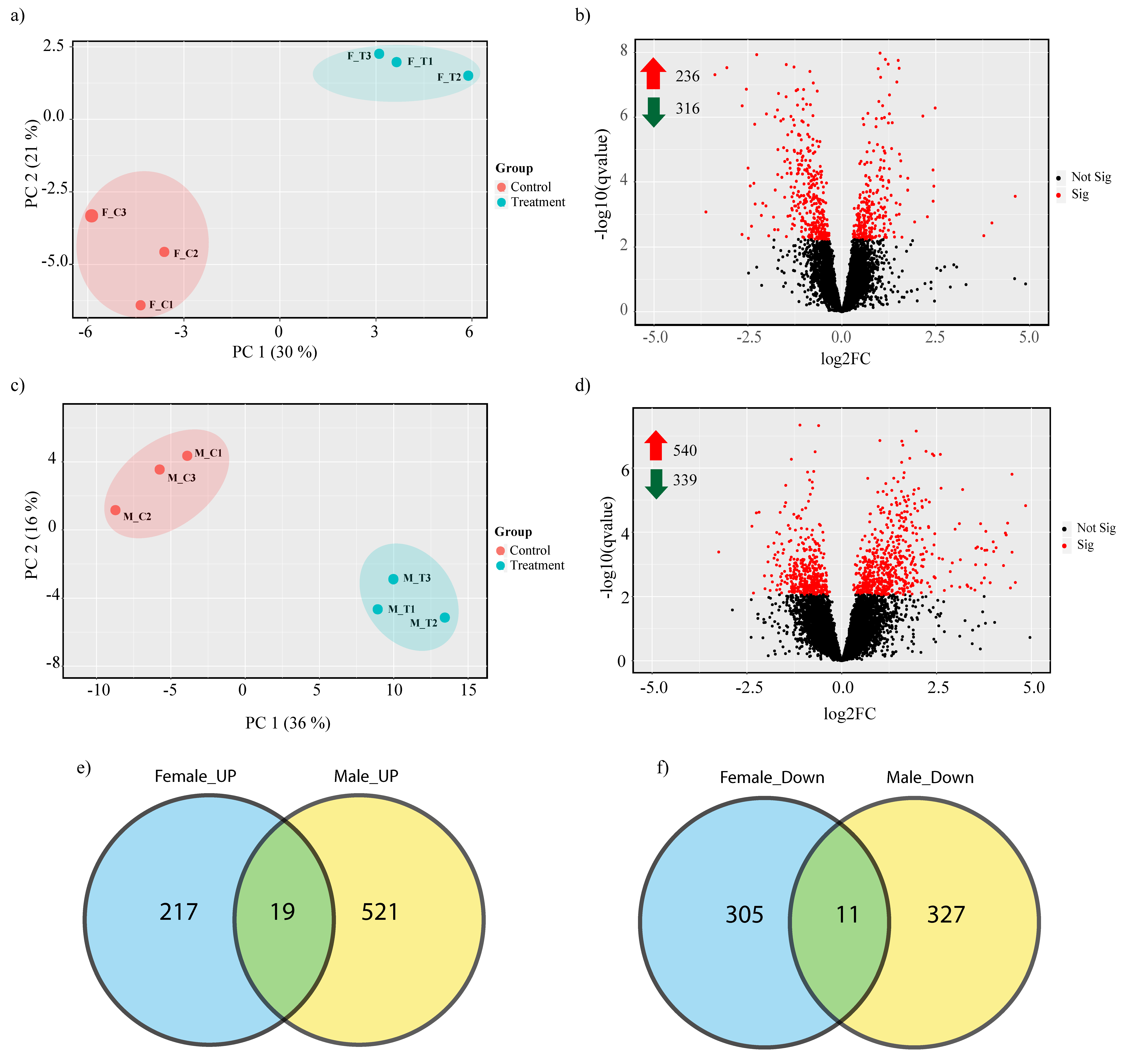
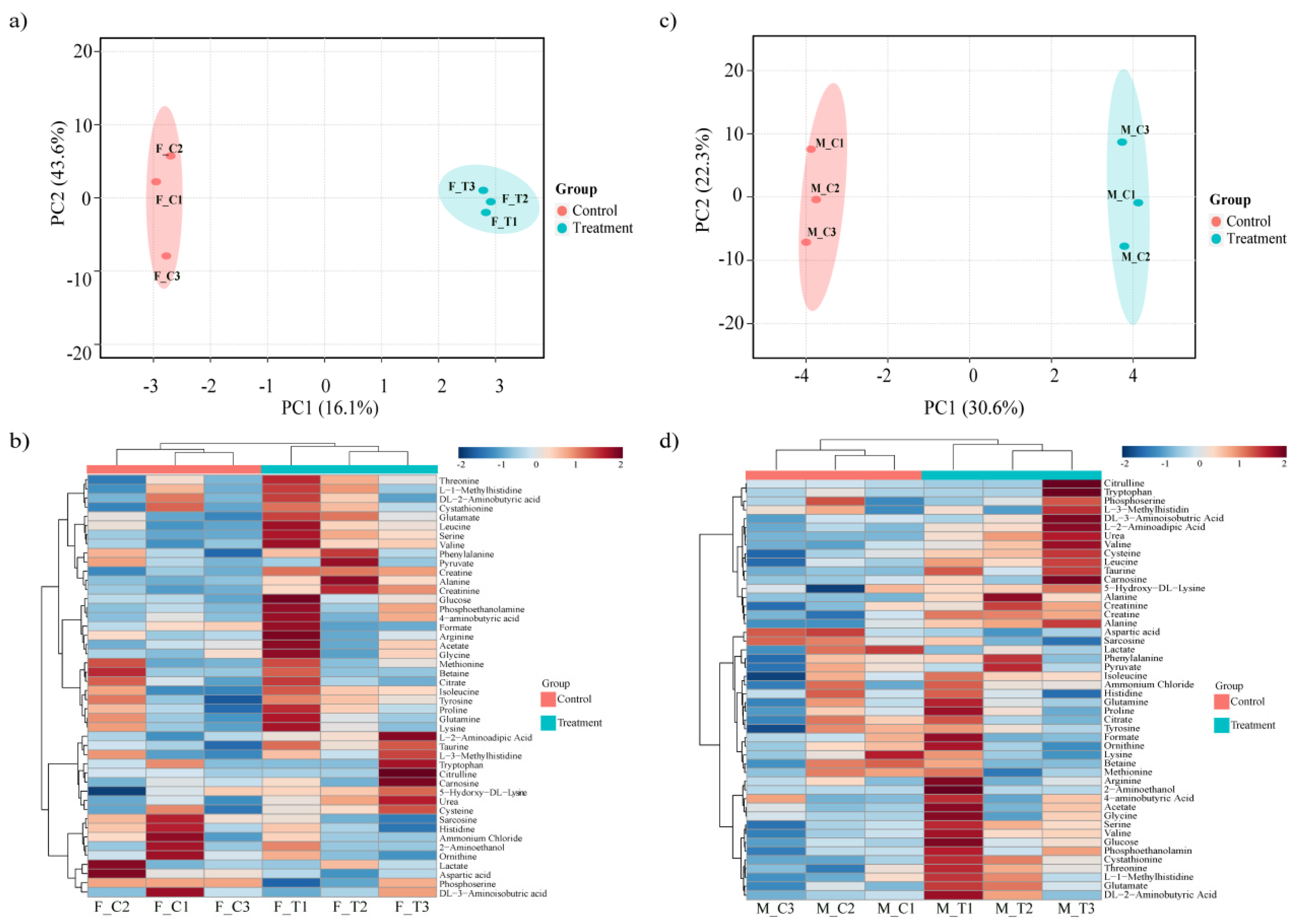
3.1. Transcriptome Alignment, Mapping and Principle Component Analysis
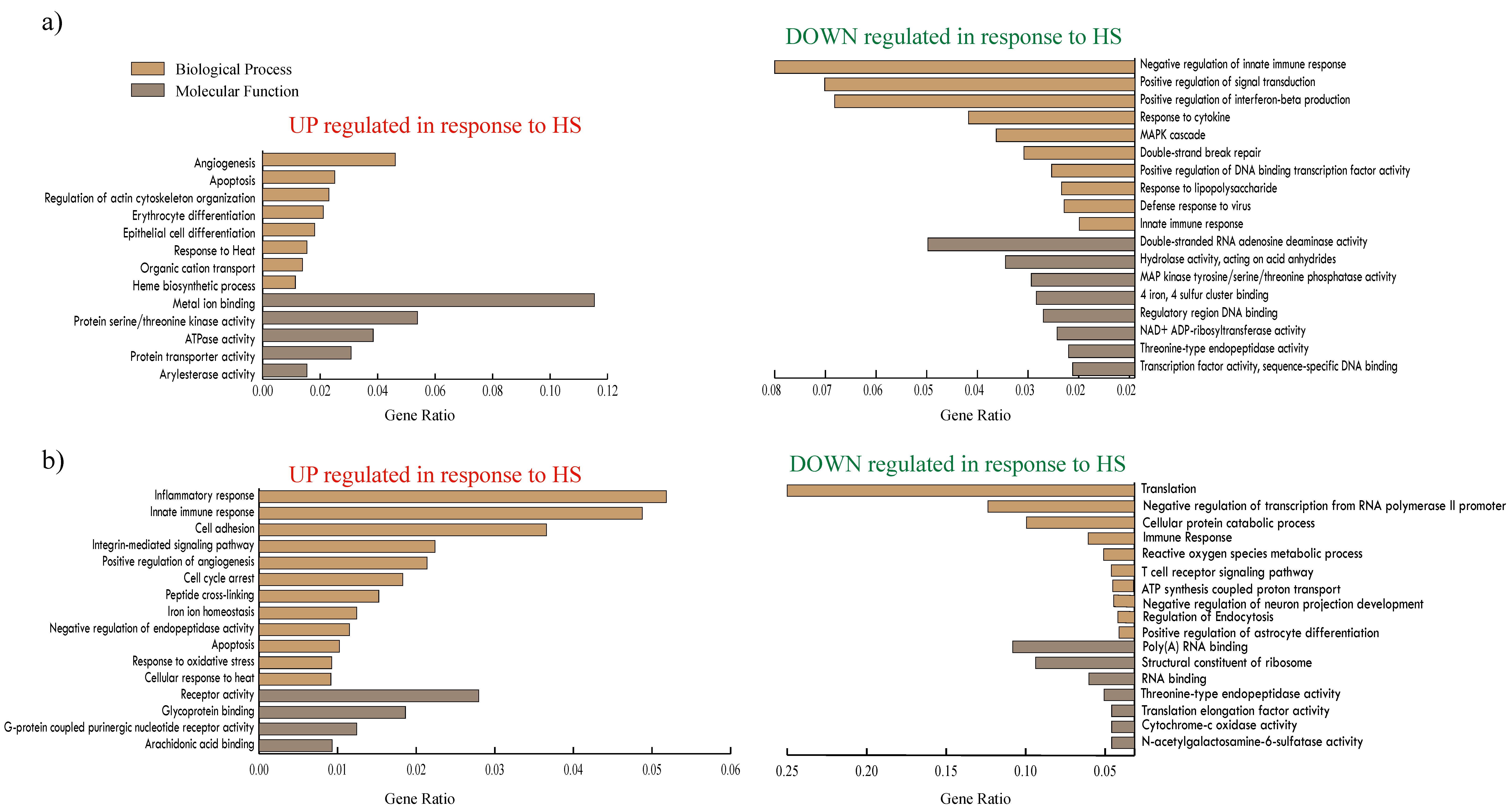
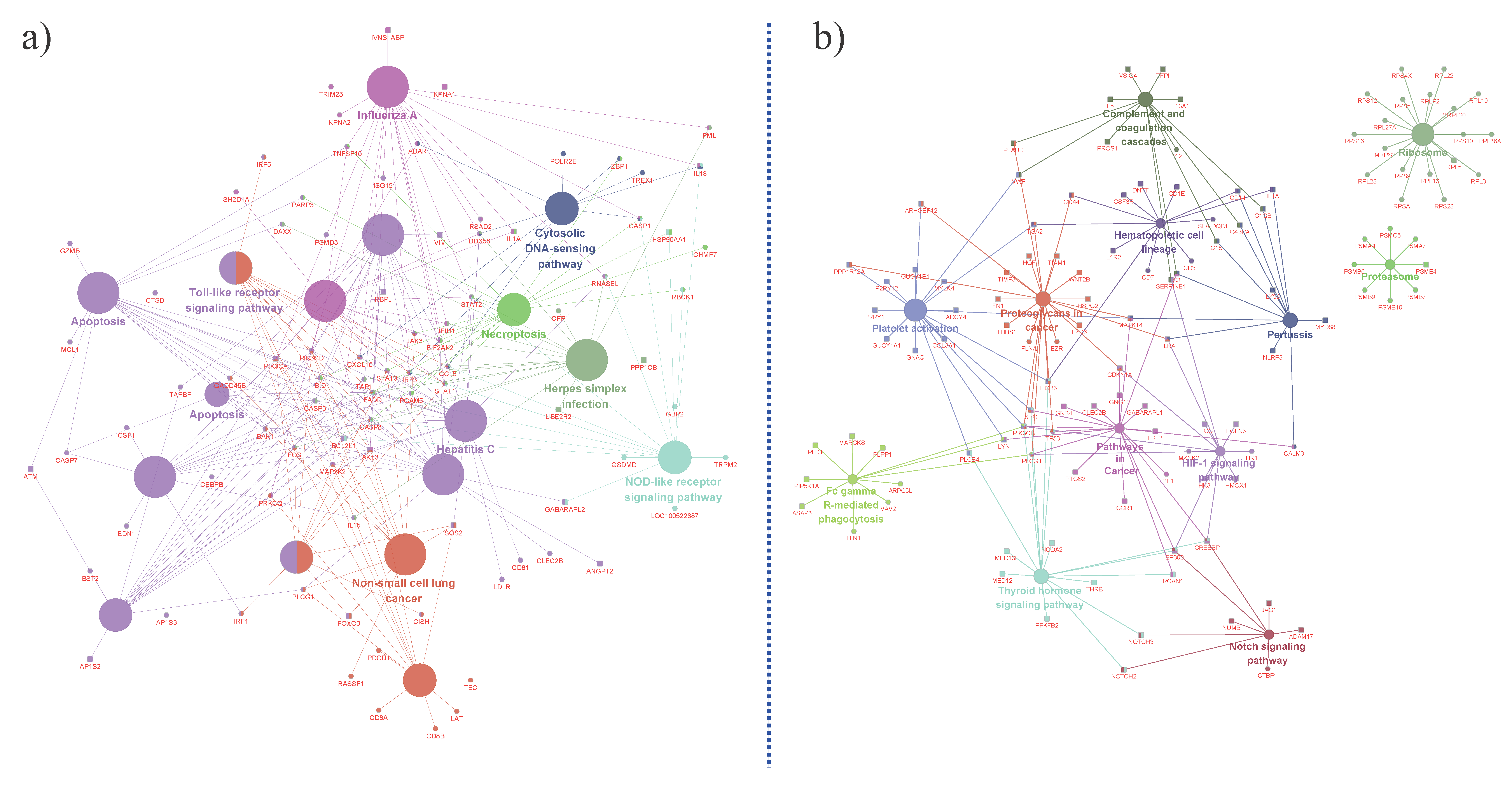
3.2. Porcine Transcriptome Response to Heat Stress
3.3. Porcine Metabolome Response to Heat Stress
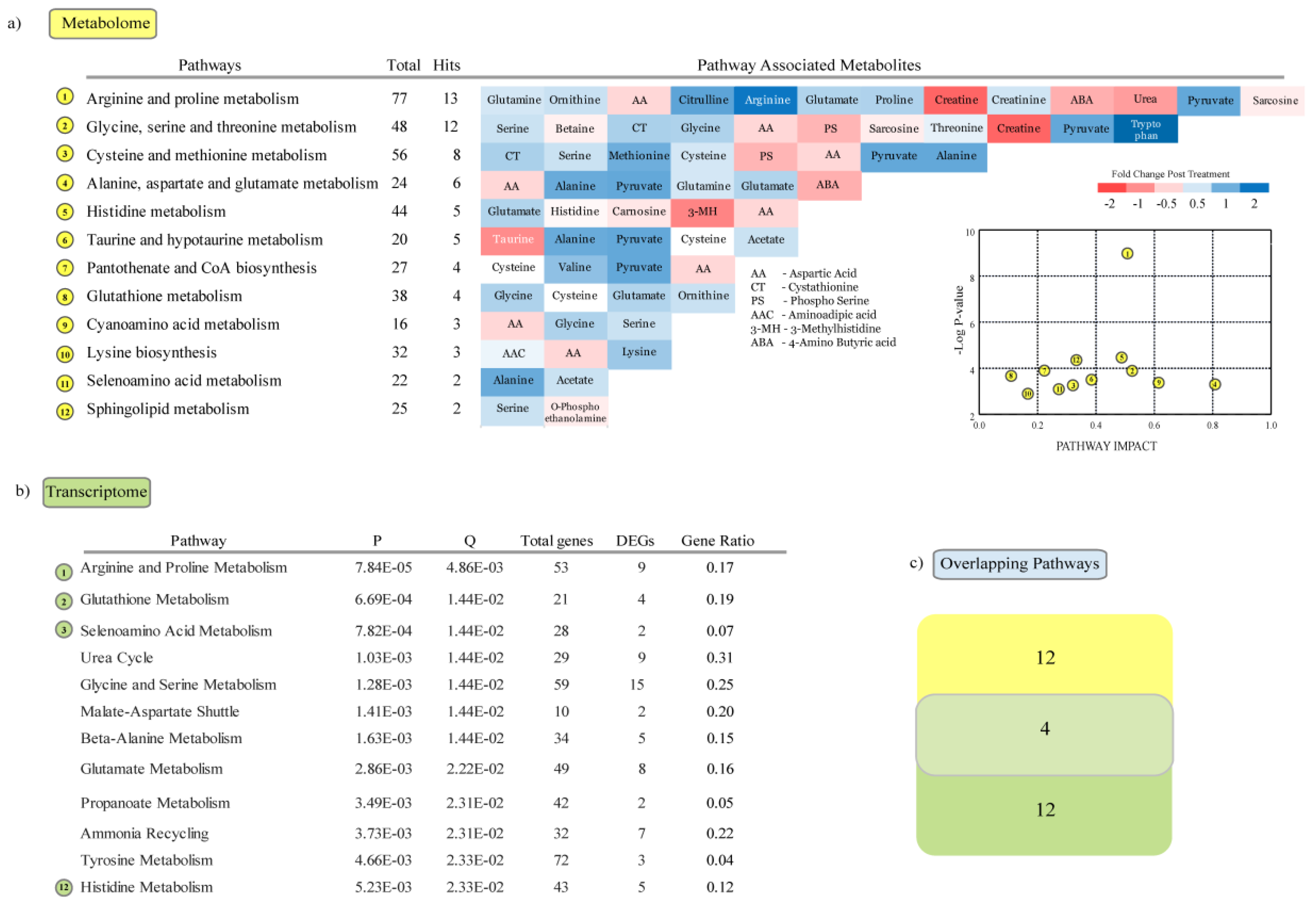
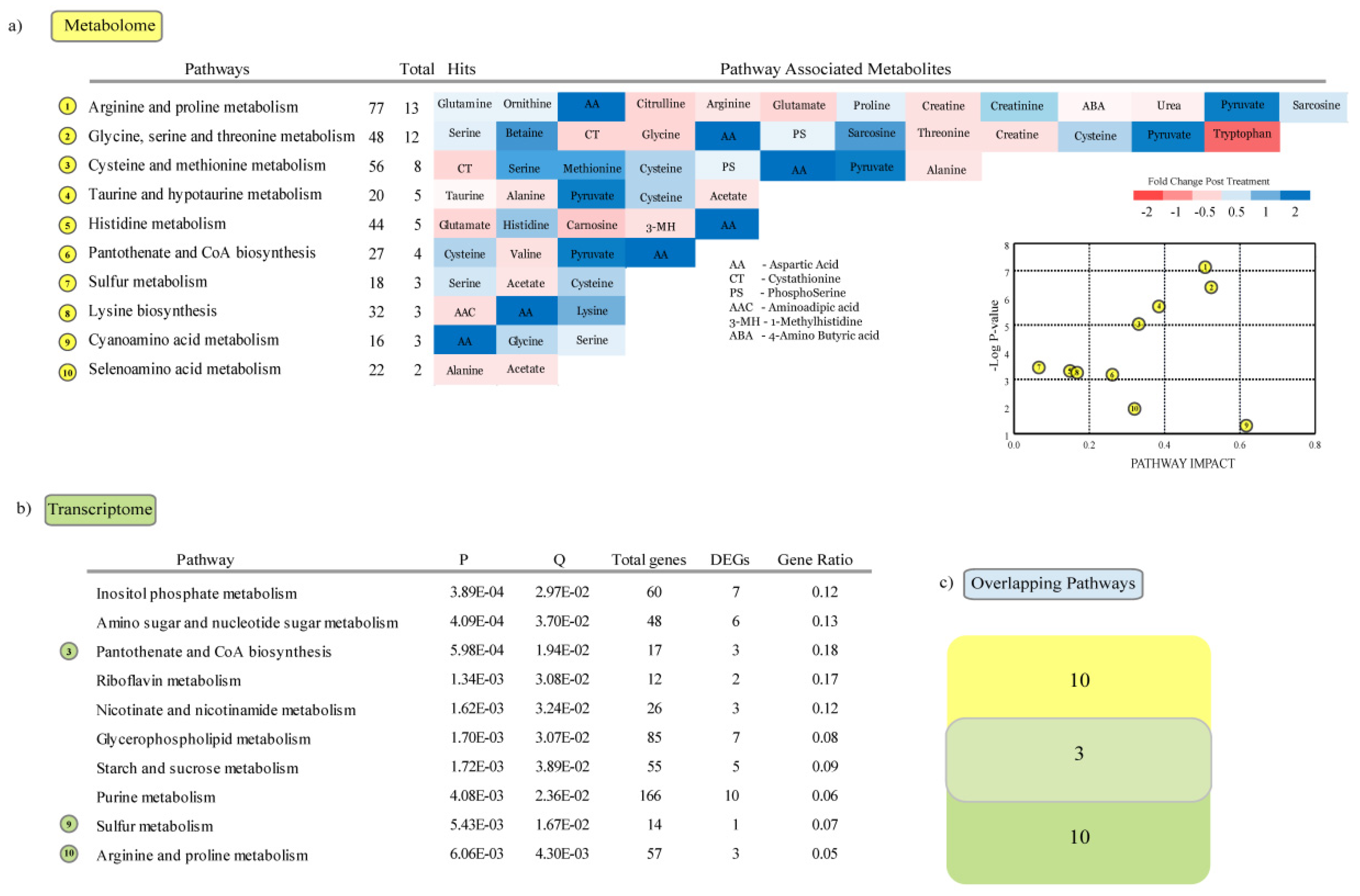
3.4. Metabolic Pathways Enriched in Response to Heat Stress
4. Discussion
4.1. Transcriptome Regulation in Response to Heat Stress
4.2. Metabolome Regulation in Response to Heat Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Renaudeau, D.; Collin, A.; Yahav, S.; De Basilio, V.; Gourdine, J.; Collier, R. Adaptation to hot climate and strategies to alleviate heat stress in livestock production. Animal 2012, 6, 707–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sejian, V.; Bhatta, R.; Gaughan, J.; Dunshea, F.; Lacetera, N. Adaptation of animals to heat stress. Animal 2018, 12, s431–s444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, J.W.; Hale, B.J.; Seibert, J.T.; Romoser, M.R.; Adur, M.K.; Keating, A.F.; Baumgard, L.H. Physiological mechanisms through which heat stress compromises reproduction in pigs. Mol. Reprod. Dev. 2017, 84, 934–945. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Cui, Y.; Gu, X. Genome-wide DNA methylation profiles changes associated with constant heat stress in pigs as measured by bisulfite sequencing. Sci. Rep. 2016, 6, 27507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Guo, H.; Zheng, W.; Xue, Y.; Zhao, R.; Yao, W. Heat stress affects fecal microbial and metabolic alterations of primiparous sows during late gestation. J. Anim. Sci. Biotechnol. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Baumgard, L.H.; Rhoads Jr, R.P. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [Green Version]
- Brown-Brandl, T.; Eigenberg, R.; Nienaber, J.A.; Kachman, S.D. Thermoregulatory profile of a newer genetic line of pigs. Livest. Prod. Sci. 2001, 71, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Patience, J.; Umboh, J.; Chaplin, R.; Nyachoti, C. Nutritional and physiological responses of growing pigs exposed to a diurnal pattern of heat stress. Livest. Prod. Sci. 2005, 96, 205–214. [Google Scholar] [CrossRef]
- Seibert, J.; Graves, K.; Hale, B.; Keating, A.; Baumgard, L.; Ross, J. Characterizing the acute heat stress response in gilts: I. Thermoregulatory and production variables. J. Anim. Sci. 2018, 96, 941–949. [Google Scholar] [CrossRef]
- Pollmann, D. Seasonal effects on sow herds: industry experience and management strategies. J. Anim. Sci 2010, 88, 9. [Google Scholar]
- St-Pierre, N.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Quiniou, N.; Noblet, J. Influence of high ambient temperatures on performance of multiparous lactating sows. J. Anim. Sci. 1999, 77, 2124–2134. [Google Scholar] [CrossRef] [PubMed]
- Dou, S.; Villa-Vialaneix, N.; Liaubet, L.; Billon, Y.; Giorgi, M.; Gilbert, H.; Gourdine, J.-L.; Riquet, J.; Renaudeau, D. 1HNMR-Based metabolomic profiling method to develop plasma biomarkers for sensitivity to chronic heat stress in growing pigs. PLoS ONE 2017, 12, e0188469. [Google Scholar] [CrossRef] [Green Version]
- Mayorga, E.J.; Renaudeau, D.; Ramirez, B.C.; Ross, J.W.; Baumgard, L.H. Heat stress adaptations in pigs. Anim. Front. 2018, 9, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Hsieh, J.C.; Schmidt, C.J.; Zhu, Q.; Lamont, S.J. Liver transcriptome response to hyperthermic stress in three distinct chicken lines. Bmc Genom. 2016, 17, 955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikanth, K.; Kwon, A.; Lee, E.; Chung, H. Characterization of genes and pathways that respond to heat stress in Holstein calves through transcriptome analysis. Cell Stress Chaperones 2017, 22, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Srikanth, K.; Lee, E.; Kwan, A.; Lim, Y.; Lee, J.; Jang, G.; Chung, H. Transcriptome analysis and identification of significantly differentially expressed genes in Holstein calves subjected to severe thermal stress. Int. J. Biometeorol. 2017, 61, 1993–2008. [Google Scholar] [CrossRef]
- Hao, Y.; Feng, Y.; Yang, P.; Cui, Y.; Liu, J.; Yang, C.; Gu, X. Transcriptome analysis reveals that constant heat stress modifies the metabolism and structure of the porcine longissimus dorsi skeletal muscle. Mol. Genet. Genom. 2016, 291, 2101–2115. [Google Scholar] [CrossRef]
- Qu, H.; Ajuwon, K.M. Metabolomics of heat stress response in pig adipose tissue reveals alteration of phospholipid and fatty acid composition during heat stress. J. Anim. Sci. 2018, 96, 3184–3195. [Google Scholar] [CrossRef]
- He, J.; Zheng, W.; Lu, M.; Yang, X.; Xue, Y.; Yao, W. A controlled heat stress during late gestation affects thermoregulation, productive performance, and metabolite profiles of primiparous sow. J. Therm. Boil. 2019, 81, 33–40. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, C.; Hao, Y.; Gu, X.; Wang, H. Chronic Heat Stress Induces Acute Phase Responses and Serum Metabolome Changes in Finishing Pigs. Animals 2019, 9, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Close, W.; Le Dividich, J.; Duee, P. Influence of environmental temperature on glucose tolerance and insulin response in the new-born piglet. Neonatology 1985, 47, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Black, J.; Mullan, B.; Lorschy, M.; Giles, L. Lactation in the sow during heat stress. Livest. Prod. Sci. 1993, 35, 153–170. [Google Scholar] [CrossRef]
- Jeong, J.Y.; Kim, M.S.; Jung, H.J.; Kim, M.J.; Lee, H.J.; Lee, S.D. NMR-based metabolomic profiling of the liver, serum, and urine of piglets treated with deoxynivalenol. Korean J. Agric. Sci. 2018, 45, 455–461. [Google Scholar]
- Srikanth, K.; Kumar, H.; Park, W.; Byun, M.-J.; Lim, D.; Kemp, S.; Te Pas, M.; Kim, J.-M.; Park, J.-E. Cardiac and skeletal muscle transcriptome response to heat stress in Kenyan chicken ecotypes adapted to low and high altitudes reveal differences in thermal tolerance and stress response. Front. Genet. 2019, 10, 993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 23 November 2019).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Leek, J.T. Svaseq: Removing batch effects and other unwanted noise from sequencing data. Nucleic Acids Res. 2014, 42, e161. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2008, 4, 44. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Lin, Y.; Jiang, Z.; Zheng, C.; Zhou, G.; Yu, D.; Cao, T.; Wang, J.; Chen, F. Dietary arginine supplementation enhances antioxidative capacity and improves meat quality of finishing pigs. Amino Acids 2010, 38, 95–102. [Google Scholar] [CrossRef]
- Cui, Y.; Hao, Y.; Li, J.; Bao, W.; Li, G.; Gao, Y.; Gu, X. Chronic heat stress induces immune response, oxidative stress response, and apoptosis of finishing pig liver: A proteomic approach. Int. J. Mol. Sci. 2016, 17, 393. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, K.Z.; Edens, F. Influence of selenium sources on age-related and mild heat stress-related changes of blood and liver glutathione redox cycle in broiler chickens (Gallus domesticus). Comp. Biochem. Physiol. Part B Biochem. Mol. Boil. 2003, 136, 921–934. [Google Scholar] [CrossRef]
- Rudolph, T.R.R.; Baumgard, L.; Selsby, J. aWhy we should sweat heat stress. National Hog Farmer Daily, 3 December 2019. [Google Scholar]
- Min, L.; Zhao, S.; Tian, H.; Zhou, X.; Zhang, Y.; Li, S.; Yang, H.; Zheng, N.; Wang, J. Metabolic responses and “omics” technologies for elucidating the effects of heat stress in dairy cows. Int. J. Biometeorol. 2017, 61, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.; Hale, B.; Gabler, N.; Rhoads, R.; Keating, A.; Baumgard, L. Physiological consequences of heat stress in pigs. Anim. Prod. Sci. 2015, 55, 1381–1390. [Google Scholar] [CrossRef]
- Qu, H. Mechanism of Adipose Tissue Specific Response to Heat Stress in Pigs; Purdue University: West Lafayette, IN, USA, 2018. [Google Scholar]
- Moeser, A. Gender and stress matter in pig gut health. National Hog Farmer Daily, 17 April 2018. [Google Scholar]
- Renaudeau, D.; Giorgi, M.; Silou, F.; Weisbecker, J. Effect of breed (lean or fat pigs) and sex on performance and feeding behaviour of group housed growing pigs in a tropical climate. Asian-Australas. J. Anim. Sci. 2006, 19, 593–600. [Google Scholar] [CrossRef]
- Jaturasitha, S.; Kamopas, S.; Suppadit, T.; Khiaosa-ard, R.; Kreuzer, M. The effect of gender of finishing pigs slaughtered at 110 kilograms on performance, and carcass and meat quality. Sci. Asia 2006, 32, 297–305. [Google Scholar] [CrossRef]
- Ekstrom, K.; Miller, E.; Ullrey, D.; Lewis, A. Genetic and sex considerations in swine nutrition. In Swine Nutrition; Miller, E.R., Ullrey, D.E., Lewis, A.J., Eds.; Butterworth-Heinemann: Oxford, UK, 1991; pp. 415–424. [Google Scholar]
- Singer, R.; Harker, C.T.; Vander, A.J.; Kluger, M.J. Hyperthermia induced by open-field stress is blocked by salicylate. Physiol. Behav. 1986, 36, 1179–1182. [Google Scholar] [CrossRef] [Green Version]
- Dymond, K.E.; Fewell, J.E. Gender influences the core temperature response to a simulated open field in adult guinea pigs. Physiol. Behav. 1998, 65, 889–892. [Google Scholar] [CrossRef]
- Martínez-Miró, S.; Tecles, F.; Ramón, M.; Escribano, D.; Hernández, F.; Madrid, J.; Orengo, J.; Martínez-Subiela, S.; Manteca, X.; Cerón, J.J. Causes, consequences and biomarkers of stress in swine: An update. BMC Vet. Res. 2016, 12, 171. [Google Scholar] [CrossRef] [Green Version]
- Bottoms, G.; Roesel, O.; Rausch, F.; Akins, E. Circadian variation in plasma cortisol and corticosterone in pigs and mares. Amer. J. Vet. Res. 1972, 33, 785–790. [Google Scholar]
- Ruis, M.A.; Te Brake, J.H.; Engel, B.; Ekkel, E.D.; Buist, W.G.; Blokhuis, H.J.; Koolhaas, J.M. The circadian rhythm of salivary cortisol in growing pigs: Effects of age, gender, and stress. Physiol. Behav. 1997, 62, 623–630. [Google Scholar] [CrossRef]
- Burdick, N.; Randel, R.; Carroll, J.; Welsh, T. Interactions between temperament, stress, and immune function in cattle. Int. J. Zool. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fagundes, A.C.A.; Negrão, J.A.; Silva, R.G.d.; Gomes, J.D.F.; Souza, L.W.d.O.; Fukushima, R.S. Environmental temperature and serum cortisol levels in growing-finishing pigs. Braz. J. Vet. Res. Anim. Sci. 2008, 45, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Devaraj, C.; Upadhyay, R. Effect of catecholamines and thermal exposure on lymphocyte proliferation, IL–1α & β in buffaloes. Ital. J. Anim. Sci. 2007, 6, 1336–1339. [Google Scholar]
- DeKruyff, R.H.; Fang, Y.; Umetsu, D.T. Corticosteroids enhance the capacity of macrophages to induce Th2 cytokine synthesis in CD4+ lymphocytes by inhibiting IL-12 production. J. Immunol. 1998, 160, 2231–2237. [Google Scholar] [PubMed]
- Salak, J.; McGlone, J.; Lyte, M. Effects of in vitro adrenocorticotrophic hormone, cortisol and human recombinant interleukin-2 on porcine neutrophil migration and luminol-dependent chemiluminescence. Vet. Immunol. Immunopathol. 1993, 39, 327–337. [Google Scholar] [CrossRef]
- Montilla, S.I.R.; Johnson, T.P.; Pearce, S.C.; Gardan-Salmon, D.; Gabler, N.K.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Lonergan, S.M.; Selsby, J.T. Heat stress causes oxidative stress but not inflammatory signaling in porcine skeletal muscle. Temperature 2014, 1, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, S.; Summers, C.; Pearce, S.; Gabler, N.; Valentine, R.; Baumgard, L.; Rhoads, R.; Selsby, J. Short-term heat stress causes altered intracellular signaling in oxidative skeletal muscle. J. Anim. Sci. 2017, 95, 2438–2451. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Brownstein, A.J.; Pearce, S.C.; Hudson, M.B.; Gabler, N.K.; Baumgard, L.H.; Rhoads, R.P.; Selsby, J.T. Prolonged environment-induced hyperthermia alters autophagy in oxidative skeletal muscle in Sus scrofa. J. Therm. Biol. 2018, 74, 160–169. [Google Scholar] [CrossRef]
- Brownstein, A.J.; Ganesan, S.; Summers, C.M.; Pearce, S.; Hale, B.J.; Ross, J.W.; Gabler, N.; Seibert, J.T.; Rhoads, R.P.; Baumgard, L.H. Heat stress causes dysfunctional autophagy in oxidative skeletal muscle. Physiol. Rep. 2017, 5, e13317. [Google Scholar] [CrossRef]
- Pan, P.J.; Hsu, C.F.; Tsai, J.J.; Chiu, J.H. The role of oxidative stress response revealed in preconditioning heat stimulation in skeletal muscle of rats. J. Surg. Res. 2012, 176, 108–113. [Google Scholar] [CrossRef]
- Nussbaum, E.L.; Locke, M. Heat shock protein expression in rat skeletal muscle after repeated applications of pulsed and continuous ultrasound. Arch. Phys. Med. Rehabil. 2007, 88, 785–790. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.; Sun, F.; Zhang, J.; Feng, J.; Liu, H.; Rajendran, K.; Sun, L.; Zhang, Y.; Jiang, Y. RNA-Seq reveals expression signatures of genes involved in oxygen transport, protein synthesis, folding and degradation in response to heat stress in catfish. Am. J. Physiol.-Heart Circ. Physiol. 2013, 45, 462–476. [Google Scholar] [CrossRef] [Green Version]
- Moseley, P.L. Heat shock proteins and heat adaptation of the whole organism. J. Appl. Physiol. 1997, 83, 1413–1417. [Google Scholar] [CrossRef]
- Liu, Y.; Steinacker, J.M. Changes in skeletal muscle heat shock proteins: Pathological significance. Front. Biosci. 2001, 6, D12–D25. [Google Scholar] [PubMed]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat. Cell Biol. 2000, 2, 326. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Buchner, J. Hsp90: Chaperoning signal transduction. J. Cell. Physiol. 2001, 188, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-Y.; Tam, M.-F.; Hsu, Y.-T.; Lin, J.-H.; Chen, H.-H.; Chuang, C.-K.; Chen, M.-Y.; King, Y.-T.; Lee, W.-C. Developmental changes of heat-shock proteins in porcine testis by a proteomic analysis. Theriogenology 2005, 64, 1940–1955. [Google Scholar] [CrossRef]
- Lei, L.; Yu, J.; Bao, E. Expression of heat shock protein 90 (Hsp90) and transcription of its corresponding mRNA in broilers exposed to high temperature. Br. Poult. Sci. 2009, 50, 504–511. [Google Scholar] [CrossRef]
- Hubler, T.R.; Scammell, J.G. Intronic hormone response elements mediate regulation of FKBP5 by progestins and glucocorticoids. Cell Stress Chaperones 2004, 9, 243. [Google Scholar] [CrossRef] [Green Version]
- Ponsuksili, S.; Du, Y.; Murani, E.; Schwerin, M.; Wimmers, K. Elucidating molecular networks that either affect or respond to plasma cortisol concentration in target tissues of liver and muscle. Genetics 2012, 192, 1109–1122. [Google Scholar] [CrossRef] [Green Version]
- Sautron, V.; Terenina, E.; Gress, L.; Lippi, Y.; Billon, Y.; Larzul, C.; Liaubet, L.; Villa-Vialaneix, N.; Mormède, P. Time course of the response to ACTH in pig: Biological and transcriptomic study. BMC Genom. 2015, 16, 961. [Google Scholar] [CrossRef] [Green Version]
- Weiß, M.; Kost, B.; Renner-Müller, I.; Wolf, E.; Mylonas, I.; Brüning, A. Efavirenz causes oxidative stress, endoplasmic reticulum stress, and autophagy in endothelial cells. Cardiovasc. Toxicol. 2016, 16, 90–99. [Google Scholar] [CrossRef]
- Paul, C.; Teng, S.; Saunders, P.T. A single, mild, transient scrotal heat stress causes hypoxia and oxidative stress in mouse testes, which induces germ cell death. Boil. Reprod. 2009, 80, 913–919. [Google Scholar] [CrossRef] [Green Version]
- Fridovich, I. Superoxide dismutases. Adv. Enzymol. Relat. Areas. Mol. Biol. 1986, 58, 61–97. [Google Scholar] [PubMed]
- Nazıroğlu, M. New molecular mechanisms on the activation of TRPM2 channels by oxidative stress and ADP-ribose. Neurochem. Res. 2007, 32, 1990–2001. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.N.; Byers, B.; Cord, B.; Shcheglovitov, A.; Byrne, J.; Gujar, P.; Kee, K.; Schüle, B.; Dolmetsch, R.E.; Langston, W. LRRK2 mutant iPSC-derived DA neurons demonstrate increased susceptibility to oxidative stress. Cell Stem Cell 2011, 8, 267–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, Y.; Yoshikawa, T. Oxidative stress involvement and gene expression in indomethacin-induced gastropathy. Redox Rep. 2006, 11, 243–253. [Google Scholar] [CrossRef]
- Berruyer, C.; Martin, F.; Castellano, R.; Macone, A.; Malergue, F.; Garrido-Urbani, S.; Millet, V.; Imbert, J.; Dupre, S.; Pitari, G. Vanin-1−/− mice exhibit a glutathione-mediated tissue resistance to oxidative stress. Mol. Cell. Biol. 2004, 24, 7214–7224. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Han, Q.; Wang, R.; Li, X.; Wang, Q.; Wang, H.; Wang, J.; Ma, Y. PRDX2 protects hepatocellular carcinoma SMMC-7721 cells from oxidative stress. Oncol. Lett. 2016, 12, 2217–2221. [Google Scholar] [CrossRef] [Green Version]
- Ali, D.; Mohammad, D.K.; Mujahed, H.; Jonson-Videsäter, K.; Nore, B.; Paul, C.; Lehmann, S. Anti-leukaemic effects induced by APR-246 are dependent on induction of oxidative stress and the NFE2L2/HMOX1 axis that can be targeted by PI3K and mTOR inhibitors in acute myeloid leukaemia cells. Br. J. Haematol. 2016, 174, 117–126. [Google Scholar] [CrossRef]
- Webb, A.E.; Brunet, A. FOXO transcription factors: Key regulators of cellular quality control. Trends Biochem. Sci. 2014, 39, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Barizzone, N.; Monti, S.; Mellone, S.; Godi, M.; Marchini, M.; Scorza, R.; Danieli, M.G.; D’Alfonso, S. Rare variants in the TREX1 gene and susceptibility to autoimmune diseases. BioMed Res. Int. 2013, 2013, 471703. [Google Scholar] [CrossRef] [Green Version]
- Suraweera, A.; Becherel, O.J.; Chen, P.; Rundle, N.; Woods, R.; Nakamura, J.; Gatei, M.; Criscuolo, C.; Filla, A.; Chessa, L. Senataxin, defective in ataxia oculomotor apraxia type 2, is involved in the defense against oxidative DNA damage. J. Cell Boil. 2007, 177, 969–979. [Google Scholar] [CrossRef] [Green Version]
- Calderwood, S.K.; Theriault, J.R.; Gong, J. How is the immune response affected by hyperthermia and heat shock proteins? Int. J. Hyperth. 2005, 21, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Urriola, P.E.; Luo, Z.h.; Rambo, Z.J.; Wilson, M.E.; Torrison, J.L.; Shurson, G.C.; Chen, C. Metabolomics revealed diurnal heat stress and zinc supplementation-induced changes in amino acid, lipid, and microbial metabolism. Physiol. Rep. 2016, 4, e12676. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhao, W.; Wang, L.; Guo, F.; Song, D.; Zhang, Q.; Zhang, D.; Fan, Y.; Wang, J. Integration of metabolomic and transcriptomic profiles to identify biomarkers in serum of lung cancer. J. Cell. Biochem. 2019, 120, 11981–11989. [Google Scholar] [CrossRef] [PubMed]
- Wheelock, J.; Rhoads, R.; VanBaale, M.; Sanders, S.; Baumgard, L. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef]
- Kamiya, M.; Kamiya, Y.; Tanaka, M.; Oki, T.; Nishiba, Y.; Shioya, S. Effects of high ambient temperature and restricted feed intake on urinary and plasma 3-methylhistidine in lactating Holstein cows. Anim. Sci. J. 2006, 77, 201–207. [Google Scholar] [CrossRef]
- Azad, M.; Kikusato, M.; Maekawa, T.; Shirakawa, H.; Toyomizu, M. Metabolic characteristics and oxidative damage to skeletal muscle in broiler chickens exposed to chronic heat stress. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 155, 401–406. [Google Scholar] [CrossRef]
- Kellaway, R.; Colditz, P. The effect of heat stress on growth and nitrogen metabolism in Friesian and F1 Brahman× Friesian heifers. Aust. J. Agric. Res. 1975, 26, 615–622. [Google Scholar] [CrossRef]
- Srikandakumar, A.; Johnson, E.; Mahgoub, O. Effect of heat stress on respiratory rate, rectal temperature and blood chemistry in Omani and Australian Merino sheep. Small Rumin. Res. 2003, 49, 193–198. [Google Scholar] [CrossRef]
- Pearce, S.; Gabler, N.; Ross, J.; Escobar, J.; Patience, J.; Rhoads, R.; Baumgard, L. The effects of heat stress and plane of nutrition on metabolism in growing pigs. J. Anim. Sci. 2013, 91, 2108–2118. [Google Scholar] [CrossRef] [Green Version]
- Pearce, S. The Effects of Heat Stress and Nutritional Status on Metabolism and Intestinal Integrity in Growing Pigs. Master’s Thesis, Iowa State University, Ames, IA, USA, 2011. [Google Scholar]
- Brockman, R. Pancreatic and adrenal hormonal regulation of metabolism. In Control of Digestion and Metabolism in Ruminants; Prentice-Hall: Upper Saddle River, NJ, USA, 1986. [Google Scholar]
- Bender, A.; Hajieva, P.; Moosmann, B. Adaptive antioxidant methionine accumulation in respiratory chain complexes explains the use of a deviant genetic code in mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 16496–16501. [Google Scholar] [CrossRef] [Green Version]
- Lawler, J.M.; Barnes, W.S.; Wu, G.; Song, W.; Demaree, S. Direct antioxidant properties of creatine. Biochem. Biophys. Res. Commun. 2002, 290, 47–52. [Google Scholar] [CrossRef] [PubMed]
- BV, S.K.; Ajeet, K.; Meena, K. Effect of heat stress in tropical livestock and different strategies for its amelioration. J. Stress Physiol. Biochem. 2011, 7, 45–54. [Google Scholar]
- Hall, G.; Lucke, J.; Lovell, R.; Lister, D. Porcine malignant hyperthermia. VII: Hepatic metabolism. Br. J. Anaesth. 1980, 52, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wu, Z.; Lin, G.; Hu, S.; Wang, B.; Dai, Z.; Wu, G. Glycine stimulates protein synthesis and inhibits oxidative stress in pig small intestinal epithelial cells. J. Nutr. 2014, 144, 1540–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-T.; Feng, L.; Jiang, W.-D.; Liu, Y.; Jiang, J.; Li, S.-H.; Zhou, X.-Q. Oxidative stress parameters and anti-apoptotic response to hydroxyl radicals in fish erythrocytes: protective effects of glutamine, alanine, citrulline and proline. Aquat. Toxicol. 2013, 126, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.J.; Kovacs-Nolan, J.A.; Yang, C.; Archbold, T.; Fan, M.Z.; Mine, Y. l-Tryptophan exhibits therapeutic function in a porcine model of dextran sodium sulfate (DSS)-induced colitis. J. Nutr. Biochem. 2010, 21, 468–475. [Google Scholar] [CrossRef]
- Yao, K.; Fang, J.; Yin, Y.; Feng, Z.-M.; Tang, Z.-R.; Wu, G. Tryptophan metabolism in animals: important roles in nutrition and health. Front. Biosci. (Schol Ed) 2011, 3, 286–297. [Google Scholar]
- Zhu, M.; Du, J.; Chen, S.; Liu, A.; Holmberg, L.; Chen, Y.; Zhang, C.; Tang, C.; Jin, H. L-cystathionine inhibits the mitochondria-mediated macrophage apoptosis induced by oxidized low density lipoprotein. Int. J. Mol. Sci. 2014, 15, 23059–23073. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Ma, X.; Luo, X.; Zhang, Y.; He, Y.; Dai, Z.; Yang, Y.; Wu, G.; Wu, Z. L-glutamine attenuates apoptosis in porcine enterocytes by regulating glutathione-related redox homeostasis. J. Nutr. 2018, 148, 526–534. [Google Scholar] [CrossRef]
- Baird, C.H.; Niederlechner, S.; Beck, R.; Kallweit, A.R.; Wischmeyer, P.E. L-Threonine induces heat shock protein expression and decreases apoptosis in heat-stressed intestinal epithelial cells. Nutrition 2013, 29, 1404–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Material | Percentage |
|---|---|
| Corn | 55.8 |
| Soybean meal product | 24.4 |
| Wheat Bran | 9 |
| Soybean Hull | 3 |
| Molasses | 3 |
| Soybean Oil | 2 |
| Limestone | 1.1 |
| Lysine | 0.4 |
| Salt | 0.4 |
| Globik SW | 0.3 |
| TCP | 0.4 |
| Methionine-50 | 0.2 |
| Nutrient | |
| Calcium | 0.63 |
| Total Phosphorus | 0.5 |
| Crude Protein | 18 |
| Crude Fat | 4.9 |
| Crude Fiber | 4.6 |
| Crude Ash | 4.4 |
| DRY MATTER | 87.5 |
| Arginine | 1.16 |
| Lysine | 1.37 |
| Methionine + Cysteine | 0.7 |
| D.Energy | 3450 Kcal/kg |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srikanth, K.; Park, J.-E.; Ji, S.Y.; Kim, K.H.; Lee, Y.K.; Kumar, H.; Kim, M.; Baek, Y.C.; Kim, H.; Jang, G.-W.; et al. Genome-Wide Transcriptome and Metabolome Analyses Provide Novel Insights and Suggest a Sex-Specific Response to Heat Stress in Pigs. Genes 2020, 11, 540. https://doi.org/10.3390/genes11050540
Srikanth K, Park J-E, Ji SY, Kim KH, Lee YK, Kumar H, Kim M, Baek YC, Kim H, Jang G-W, et al. Genome-Wide Transcriptome and Metabolome Analyses Provide Novel Insights and Suggest a Sex-Specific Response to Heat Stress in Pigs. Genes. 2020; 11(5):540. https://doi.org/10.3390/genes11050540
Chicago/Turabian StyleSrikanth, Krishnamoorthy, Jong-Eun Park, Sang Yun Ji, Ki Hyun Kim, Yoo Kyung Lee, Himansu Kumar, Minji Kim, Youl Chang Baek, Hana Kim, Gul-Won Jang, and et al. 2020. "Genome-Wide Transcriptome and Metabolome Analyses Provide Novel Insights and Suggest a Sex-Specific Response to Heat Stress in Pigs" Genes 11, no. 5: 540. https://doi.org/10.3390/genes11050540