1. Introduction
This study addresses the physiology of caudal photoreceptor in crayfish, as model preparation for the analysis of neuronal control mechanisms of circadian rhythmicity and photoreception by light-sensitive non-visual neurons [
1,
2]. Natural fluctuations of light and darkness modulate the physiology and behavior of crustacean decapods. Illumination during the day varies ~9 logarithmic intensity units [
3]. Aquatic organisms, for instance, receive a proportion of blue and green light which depends on the distance from the surface water [
4]. Freshwater crayfish are a suitable model to study the functional role of three photo-sensitivity systems: (1) in the retina, (2) non-visual photoreceptors in the supraesophageal ganglion (brain), and (3) caudal photoreceptors in the sixth abdominal ganglion (AG) [
1,
5,
6,
7].
This study focuses on caudal photoreceptor light responsiveness from both left and right (CPR-L and CPR-R, respectively) anatomic sides. The CPR is a primary photosensitive neuron, one for each side of the sixth AG [
8,
9]. The axon of the CPR projects from this ganglion to the brain [
10]. This photosensitive neuron discharges spontaneous action potentials in darkness and a phasic-tonic activity in direct response to higher illumination levels. It also acts as an interneuron that receives synaptically mechanosensory input from the crayfish tail [
8,
9]. In some crustaceans from marine and freshwater sites, these photosensitive neurons also show similarities in their light responsiveness [
11]. Furthermore, the firing rate of spikes from the CPR can be related to properties of a pacemaker [
8,
12]. In crayfish, prior studies have suggested that the CPR also has a functional role in the circadian system [
1,
2,
7].
The underlying phototransduction occurs in CPR dendrites [
9,
13]; other results have shown only partial knowledge regarding this phototransduction mechanism [
14,
15]. Additional studies have suggested the specificity for color light sensitivity of the CPR from diverse species of freshwater crayfish, performed at different temperatures. For example, the CPR did not respond to red light at 17 to 19 °C [
16]. Stimulation with green (546 nm) monochromatic light caused a higher discharge of action potentials from the CPR at 20 to 25 °C [
17]. Simultaneous recordings from both caudal photoreceptors, by stimulating with green light (502 nm), led to a slightly faster response for CPR-L than -R, at 15 to 18 °C [
18]. Other results have illustrated light-induced activity from the CPR by using a cold white light lamp (light emitting diode), which emitted light with two peaks: blue (446 nm) and green (557 nm) [
19]. Kingston and Cronin [
20] identified two visual opsin proteins in the crayfish
P. clarkii. The short-wavelength-sensitive (SWS) opsin from blue light (440 nm), and the long-wavelength-sensitive (LWS) opsin from green light (530 nm). Both opsins are located in the sixth abdominal ganglion, and hence it has been postulated that these opsins participate in photoreception from the CPR [
20,
21]. The absorption properties of the light wavelength of the photopigments depend on their opsins [
22,
23].
Left–right asymmetry is a characteristic of the brain from vertebrate species and some invertebrate species [
24], which arises from embryogenesis in the organism [
25,
26,
27]. Crustacean decapods showed the left and right asymmetry in some anatomical regions [
28,
29,
30]. Besides, some functional properties are asymmetric, as examples, lateral movement towards escape direction in shrimp [
29], and the performance of limbs during aggressive behavior of crayfish [
30]. The sixth AG develops from two embryonic neuromeres in the crayfish
Procambarus clarkii [
31,
32]. The CPR-axons are in the ventral of cord, one fiber into Wiersma area 82 of the left hemicord, and another fiber in region 79 on the right hemicord, of the cord map of the crayfish [
33]. Moreover, the CPR response is asymmetric from lateralized water movements, the ipsilateral CPR is excited, and the contralateral CPR is inhibited [
9]. The CPRs seem to code this directionality, and it is likely that they contribute to a turning response in crayfish [
10]. A recent study suggests that the left and right caudal photoreceptors show asymmetry in spontaneous action potential discharge in the darkness, as well as with respect to their white light-induced response. In the dark, most (70%) of the CPR-L showed an activity higher that CPR-R. Furthermore, a pulse of white light (700 Lux, 4 s) causes in most (70%) of the CPR- R showed a higher response than the CPR-L. In both conditions, the difference was significant. Moreover, a differential effect of temperature on CPRs activity from the crayfish
Cherax quadricarinatus was also noted [
34].
This study focusses on the crayfish caudal photoreceptor as a model to obtain further understanding of the photoreceptor functions. Responsiveness to blue and green monochromatic light pulses from both CPR-L and CPR-R are determined because these extra-retinal photoreceptors possess both photopigments. The effects of temperature on these processes are also analyzed.
2. Materials and Methods
2.1. Animals
Adult freshwater crayfish Cherax quadricarinatus (n = 12) of weights 30–40 g, and carapace of lengths 11–13 cm from the rostrum to telson were used for analyses. Animals were acquired from a local provider and maintained in the laboratory in aerated water containers for two weeks prior to experiments, with 12:12-h light–dark cycles, using white light of 560 Lux intensity. Animals had free access to vegetables and dried fish as food.
2.2. Extracellular Recordings
Electrophysiological experiments were performed following a previously described procedure [
34]. Briefly, we isolated the abdominal
ganglia chain from the crayfish under ice-cold conditions, with a modified physiological saline Van Harreveld (VH) solution [
35], (205 mM NaCl, 5.4 mM KCl, 2.6 mM MgCl
2, 13.5 mM CaCl
2, and 10 mM HEPES), and pH to 7.4. We dissected in the isolated sixth AG some nerve bundles between the fifth and sixth AG using a stereoscopic microscope. Then, the biological preparations were transferred to the recording chamber mounted on a microscope (SMZ800, Nikon, Tokyo, Japan) with VH saline solution perfusion (~1 mL/min) in the dark. The sixth AG was typically maintained at room temperature 24 (±1) °C. For some experiments, these ganglia were stabilized at 14 (±1) °C by supplying water from a temperature-regulated bath. As shown in
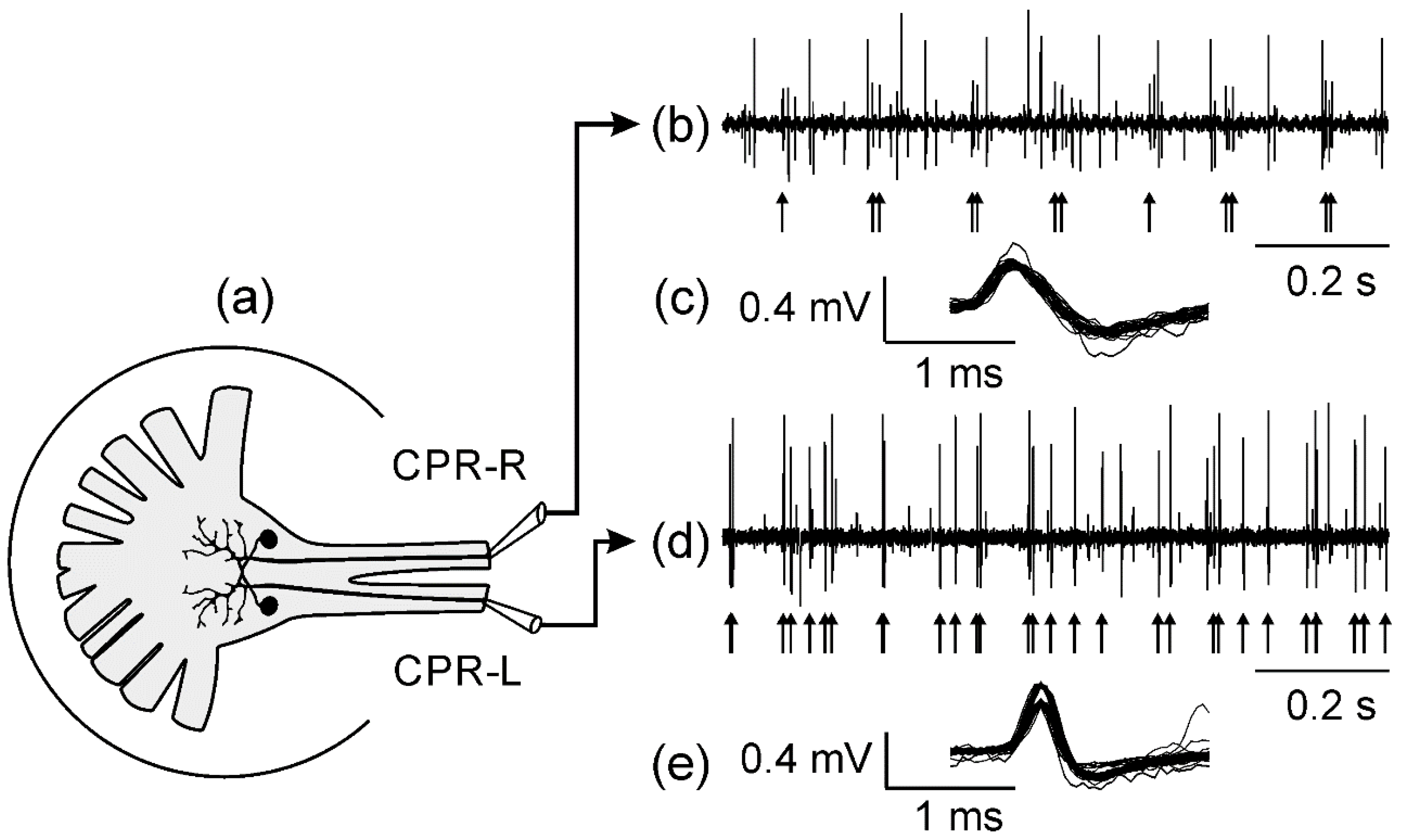
Figure 1a, extracellular recordings were performed with suction electrodes filled with the saline solution of VH, with these electrodes being positioned on the right and left hemicord, respectively, and the reference electrode (Ag-AgCl) placed in the bath solution. We acquired signal recordings by using AC amplifiers (EX1, Dagan MN, USA) filtered at 30 Hz to 10 kHz with a band-pass filter system, and displayed on an oscilloscope (TDS460, Tektronix, OR, USA).
As described earlier, the CPR photoresponse is dependent on light intensity and the time of day [
1]. We performed the experiments from 11:00 a.m. to 2:00 p.m. We triggered light pulses using a photostimulator (PS33, Grass, RI, USA), which produced white incandescent light; by using color filters (Grass) for blue (λmax = 485 nm) and green light (λmax = 530 nm). The blue light intensity ranged from 13.96 to 14.71 (log photons/cm
2/s), and for green light 14.37 to 15.35 (log photons/cm
2/s). We calibrated the wavelength and light intensity output using a spectroradiometer (Acton Research Corporation, CA, USA) and a light meter (Coherent, model FieldMax II, CA, USA), respectively. Electrophysiological recordings were sent to a computer using Spike2 software and a Micro-1401 AD board (CED, Cambridge, UK). The sorting spikes were made off-line by the Spike2 software, which studied differences in waveform, amplitude, and duration for separating spikes from different putative neurons, and their temporal firing rate in response to the light stimulus.
2.3. Statistical Analysis
The CPRs showed spontaneous activity, determined as the firing rate of action potentials per second (FRSAD) and here described with mean, median values, and interquartile range (IQR). CPR response to monochromatic light pulses was expressed as the variation in the frequency of action potentials per second (ΔF (impulses/s)), subtracting the mean value of its spontaneous activity in the dark (see Equation (1)).
The CPR photoresponse was determined for the following 10 s with respect to light pulses, with means and standard deviations being calculated, as well as median values and their IQR. We analyzed CPR firing rate spikes and the parameters: amplitude, time to peak, and duration of the recorded action potentials. We analyzed data normality using Shapiro–Wilk test (S–W-test) or Kolmogorov–Smirnov test (K–S-test). To compare the differences from data of the CPR-L and the CPR-R, we employed the independent two-sample t-test, with equal variance not assumed (Welch correction). For those parameters that did not follow a normal distribution, we used nonparametric statistical test. Thus, we compared the firing rate of the spontaneous activity (impulses/s), and the photoresponse ΔF (impulses/s), between left and right CPR activities for two samples, according to the Mann–Whitney U-test. Statistical significance was determined as p < 0.05, using Origin software (OriginLab, Northampton, MA, USA).
4. Discussion
This research is our first attempt to analyze the responsiveness of left and right caudal photoreceptors caused by blue and green monochromatic light pulses. This study provides robust data to demonstrate the different responses in the CPR neurons located in the left and right side of the last ganglion of an isolated nerve cord including the intensities of light, colors of light, and temperature, by extracellular recordings of action potentials. Our results support the presence of the SWS, and the LWS opsins in the CPRs, which showed sensitivities to blue and green light, respectively [
20,
21]. In addition, the current results provide support for the functional asymmetries described recently as a novel property for both CPR-L and -R when comparing their responses induced by white light pulses in the same species of crayfish
C. quadricarinatus [
34]. At room temperature of 24 (±1) °C, we found a significant difference in the amplitude, time to peak, and duration in action potentials extracellularly recorded from CPR-L and -R in most of the comparisons (
Table 1,
Table 2,
Table 3,
Table 4,
Table 5 and
Table 6). Some factors could contribute to these differences: the position within the connective tract connective of the CPR-L-axon is in Wiersma area 82, which is more superficial with respect at to the position of the CPR-R-axon that is in area 79 of the cord map of the crayfish [
33,
34]. The waveform of the extracellular action potential varies with electrode position relative to the recorded cell [
36], the amplitude of the action potential is function on the diameter of the axon [
37] and neurons with many dendrites will produce large amplitude spikes [
38]. Even more, there is an inverse relationship between the impulse amplitude and duration: the higher the impulse amplitude, the shorter the impulse duration [
39].
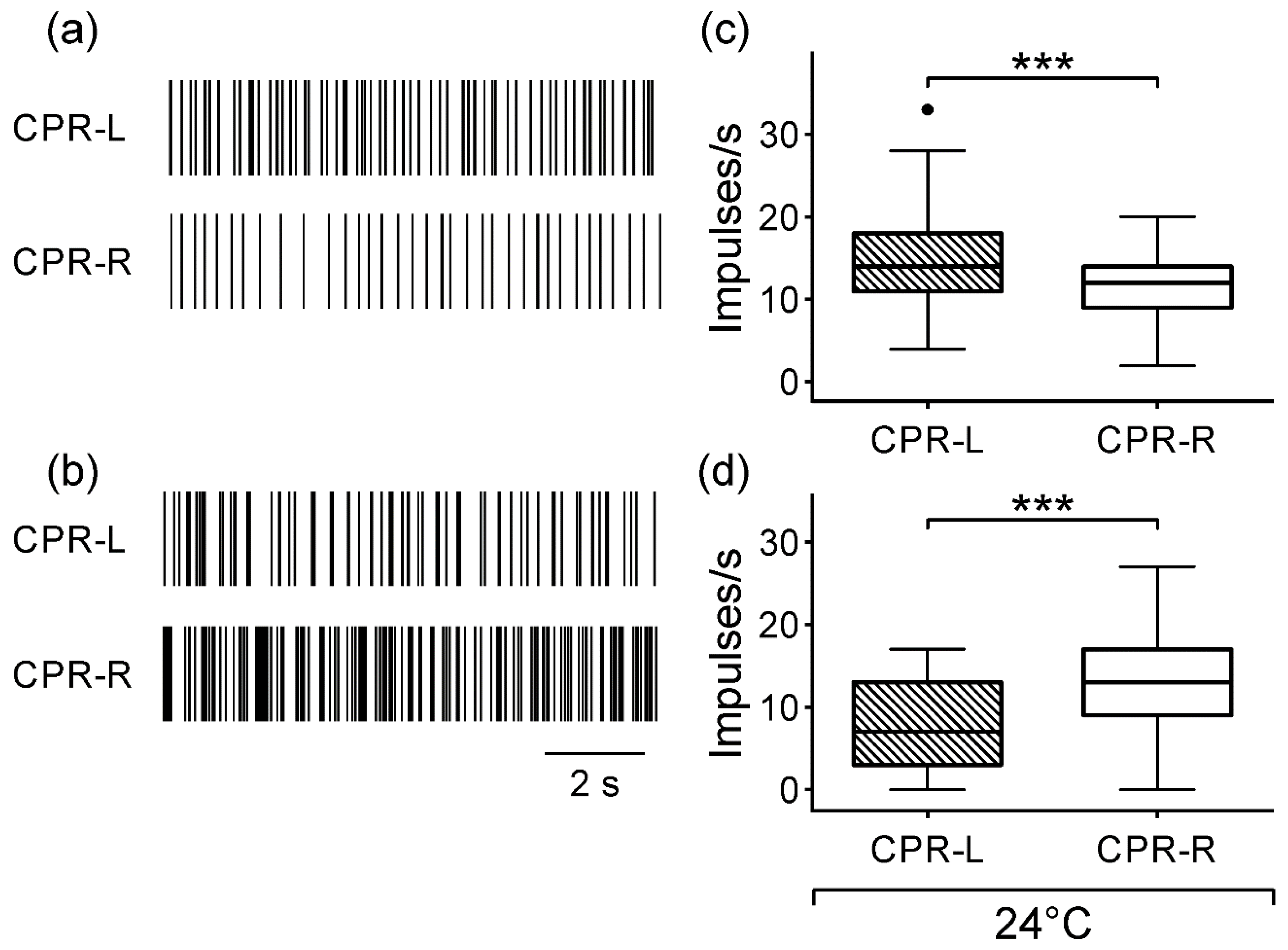
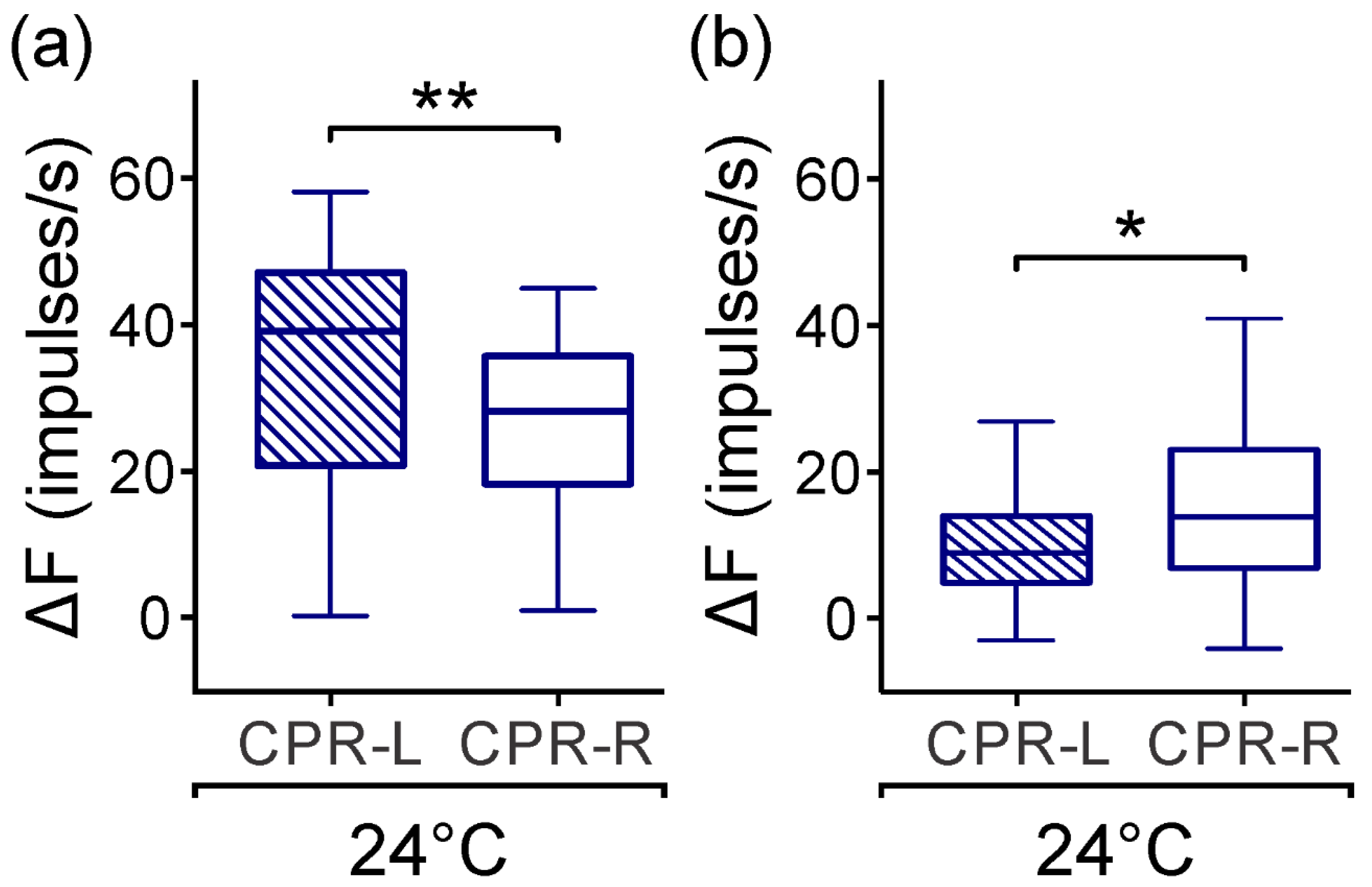
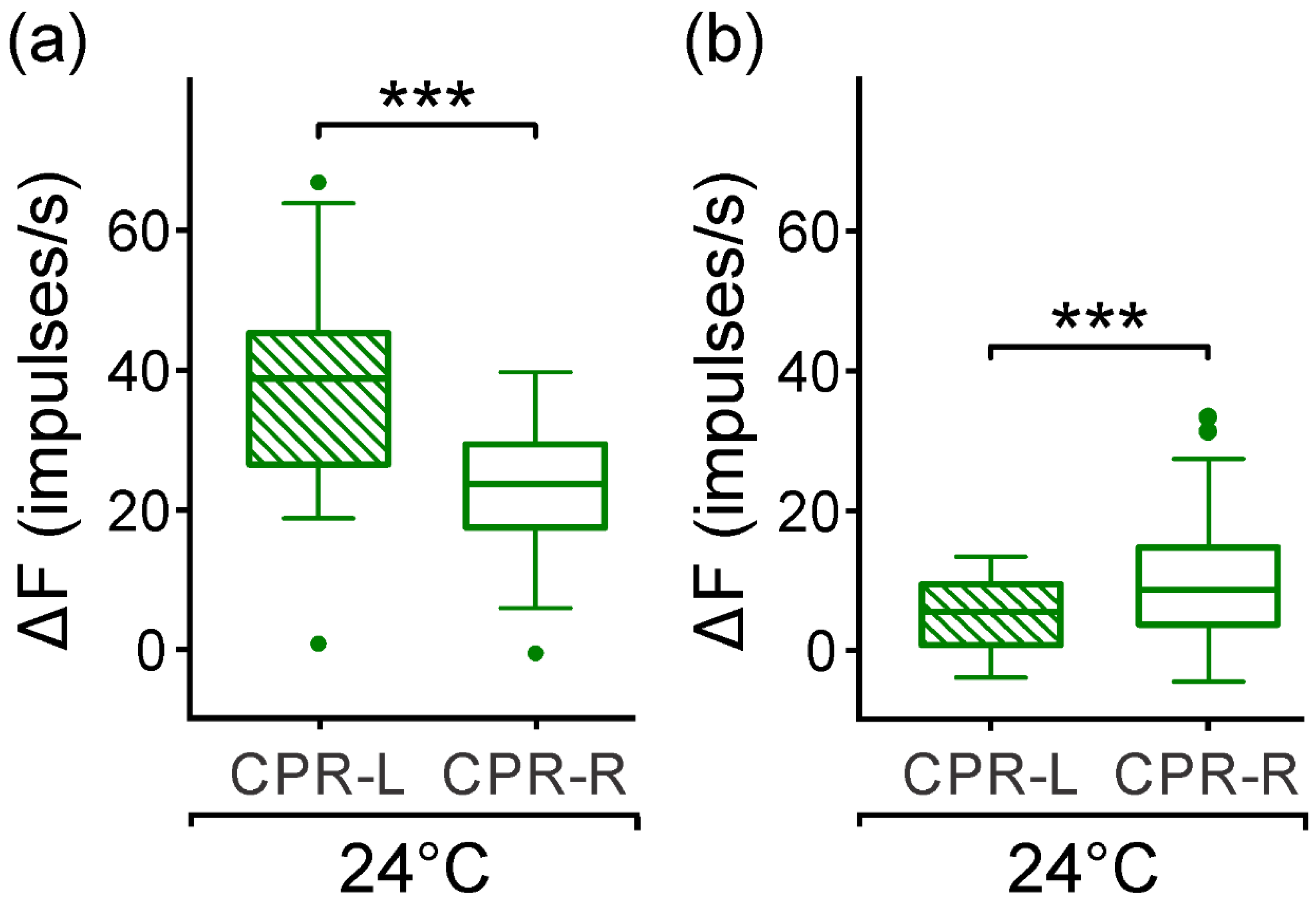
Moreover, both left and right CPRs in the isolated ganglion showed a spontaneous asymmetric activity in darkness (
Figure 1). In a subset, the CPR-L showed a median firing rate 14% higher than CPR-R (
Figure 2c), and in another subgroup, the CPRs showed an opposed asymmetry of 46% (
Figure 2d). At this temperature, we may consider that the endogenous pacemaker activity displays a regular spontaneous firing rate and responds to synaptic inputs to the nervous system in the crayfish [
40].
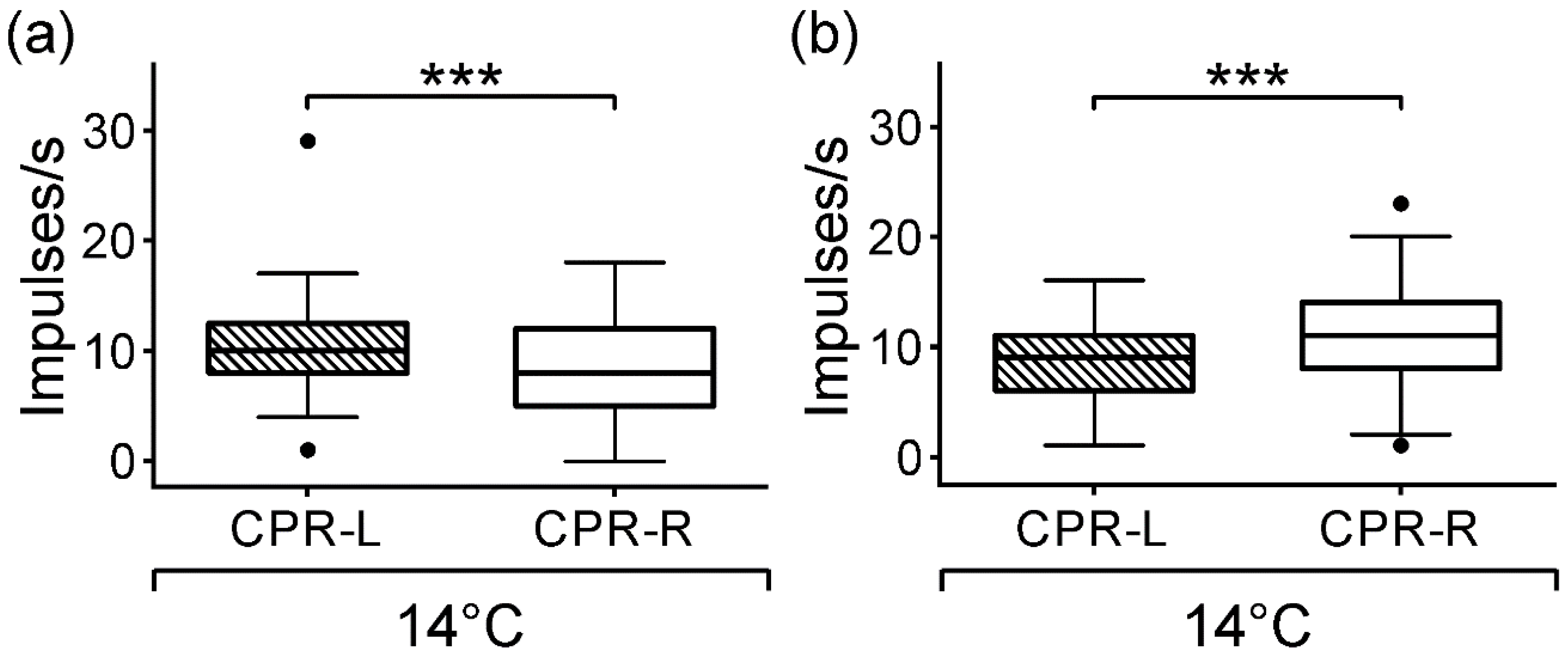
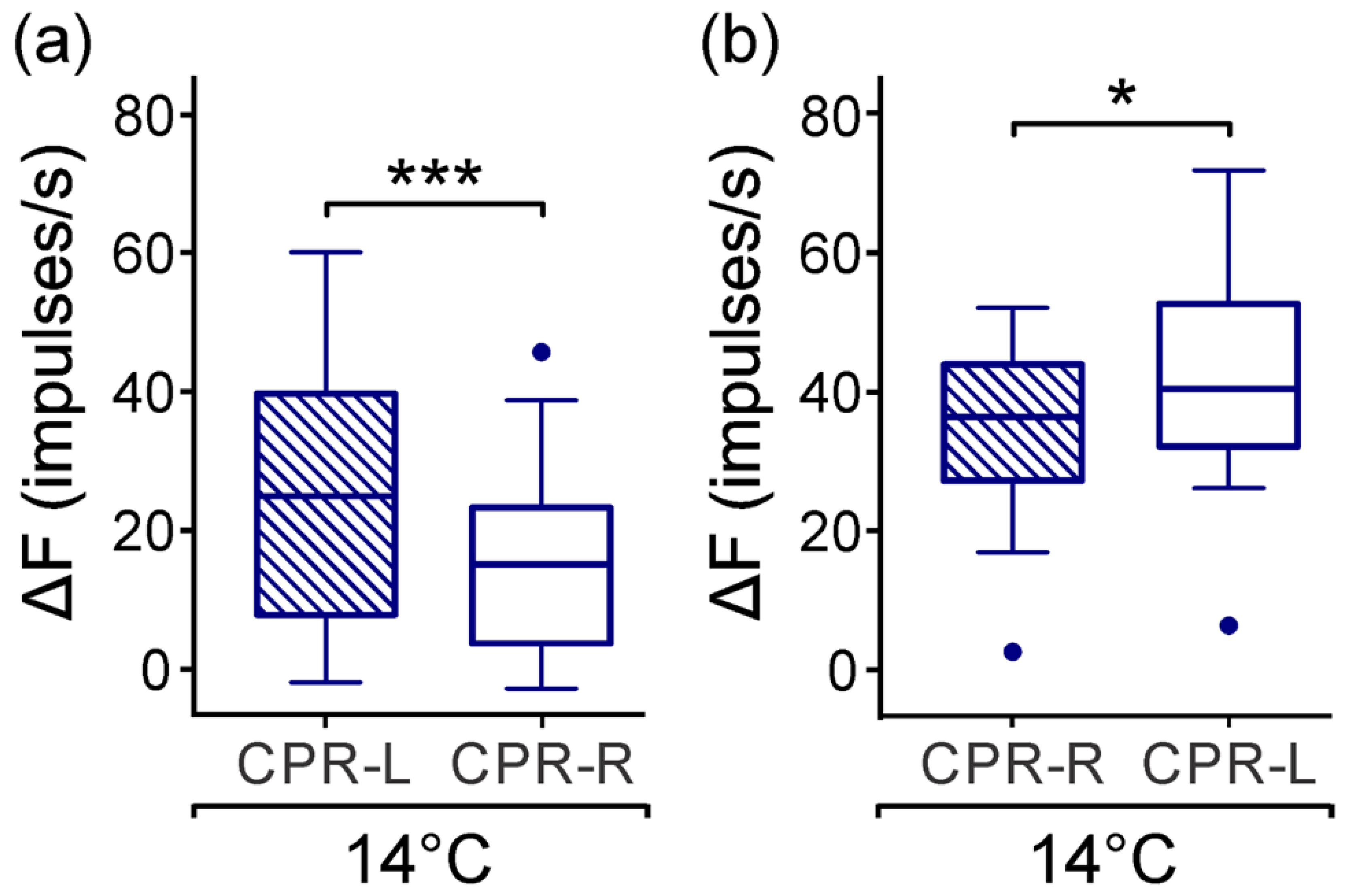
In earlier studies, the effect the temperature on the CPRs was observed, by incubating the ganglion in a cold saline solution. A quiescent spontaneous activity was displayed, and in this manner, the light-induced activity appears robust [
8,
40]. In fact, we found that at a temperature of 14 (±1) °C, the CPRs diminished the firing rate in darkness and maintained significant asymmetric activities (
Figure 3). Moreover, CPR showed an abrupt increase in the inter-spike intervals by reducing the bath temperature to 13.5 °C, and this homoclinic bifurcation suggested a thermally sensitive property from this primary photosensitive neuron [
41].
In fact, the CPRs receive inhibitory and excitatory synaptic inputs [
8,
13,
16]. Also, various substances postulated as neurotransmitters or modulators differentially influence the spontaneous activity and photoresponse from the CPR [
2]. This asymmetric activity in darkness can be a balanced state of inhibitory and excitatory effects of these modulators in the firing rate from both left and right caudal photoreceptors. As suggested, synaptic activity regulates the fluctuations from neuronal networks of the spinal cord, hippocampus, and in the leech ganglia [
42,
43].
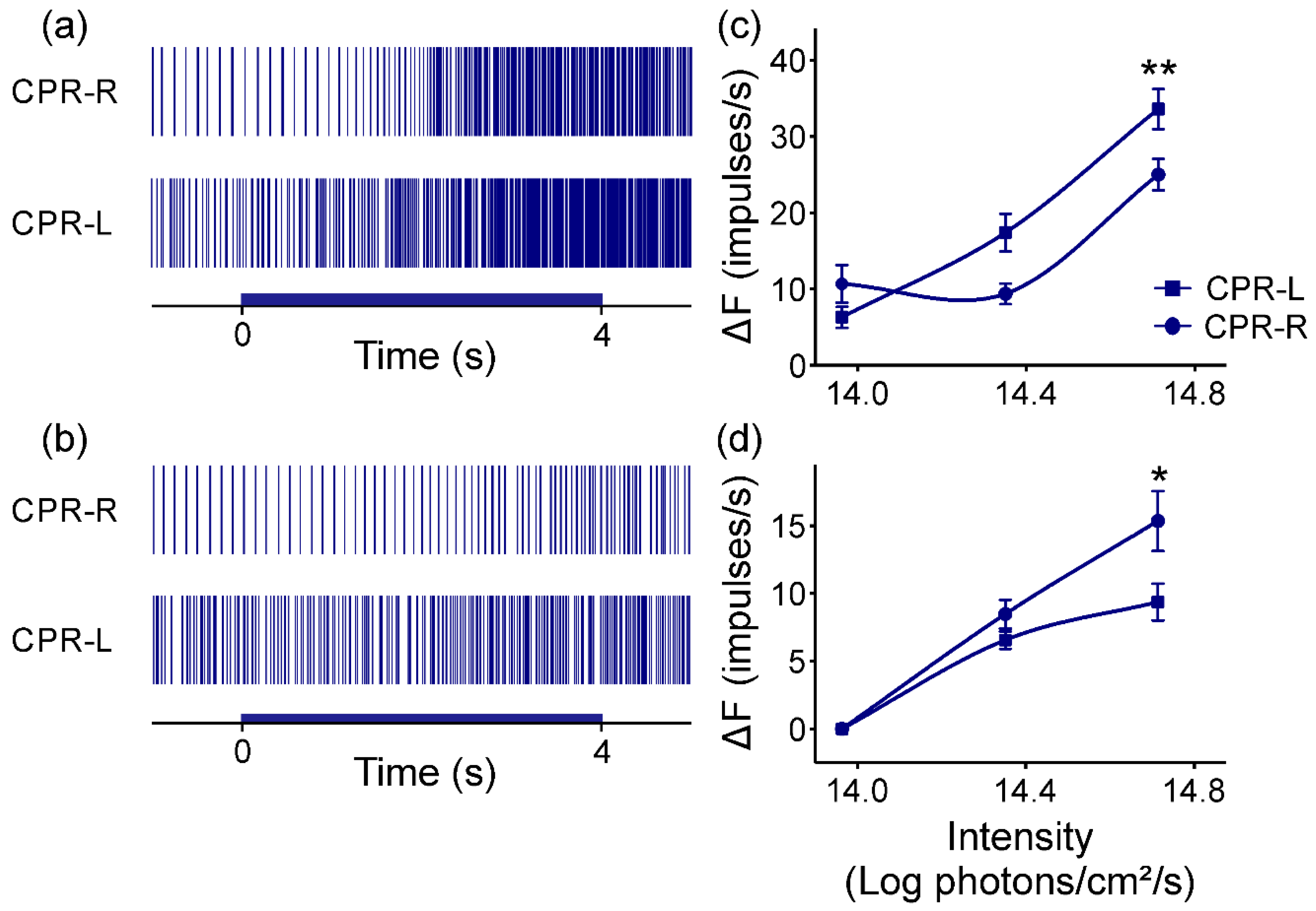
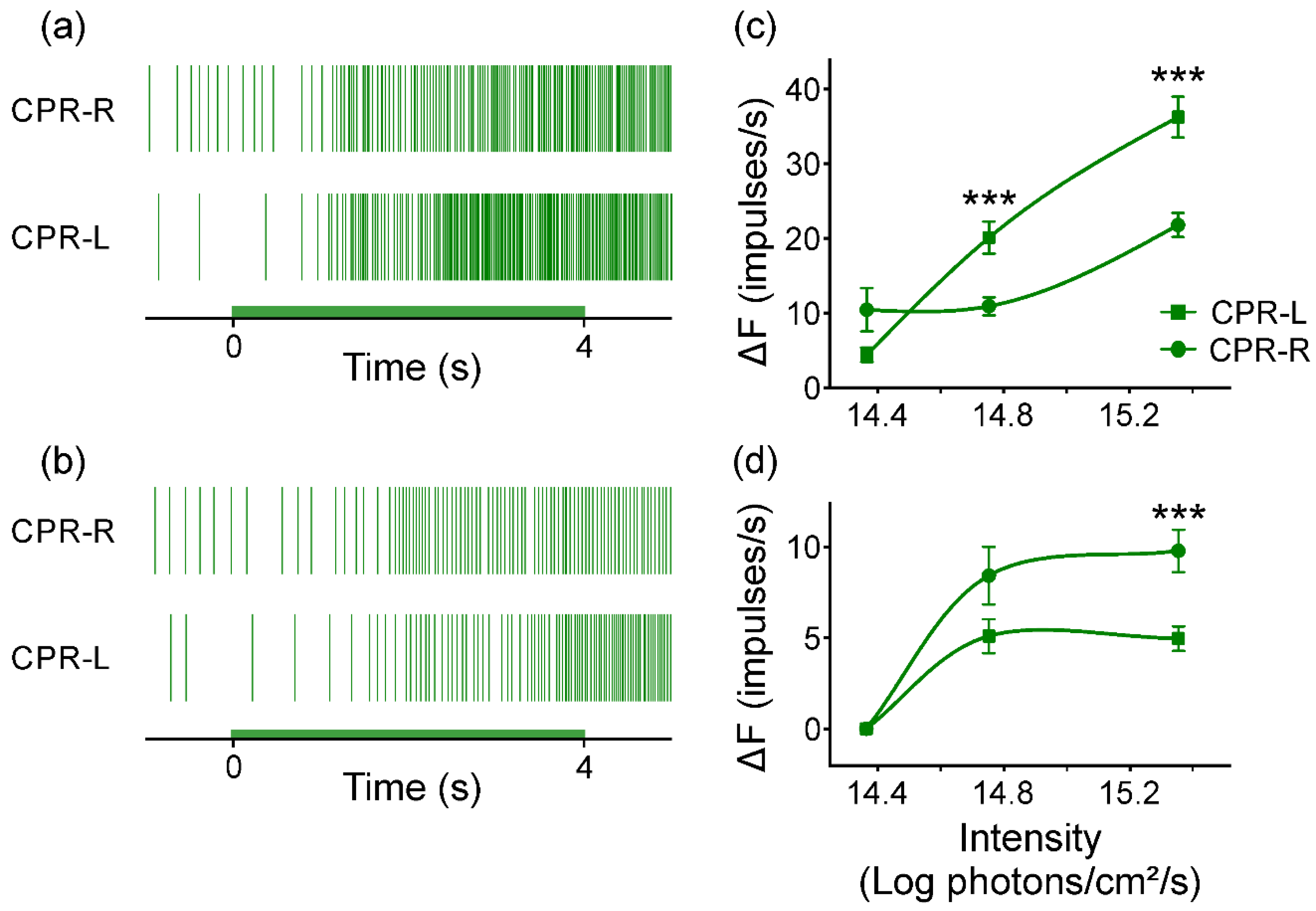
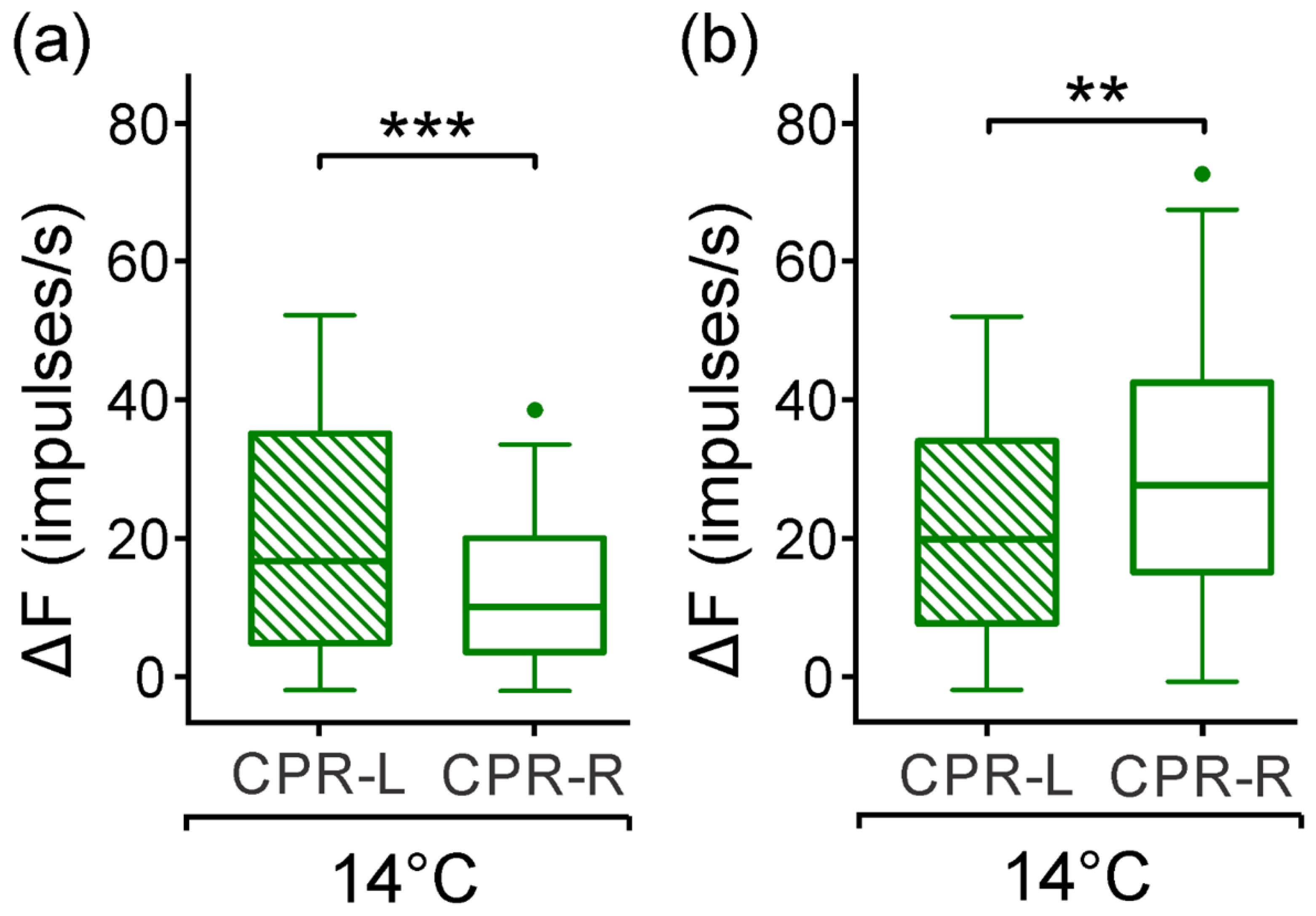
The caudal photoreceptors display both monochromatic light sensitivities after dark adaptation at two temperatures. At 24 (±1) °C, their responses depend on light intensity at a range of 1.4 logarithmic intensity units. There was approximately 0.5 logarithmic intensity units more responsive to blue light than to green (
Figure 4 and
Figure 7), with latencies and time to peaks diminishing depending on light intensities. The differential photoresponse observed between both right and left photosensitive neurons for both monochromatic light pulses could be attributed to a distinct molecular expression of photopigments SWS and LWS. Battelle [
44] reviewed the physiology of opsins with low expression levels in another extraocular photoreceptor model. Lowering the temperature to 14 (±1) °C for the CPRs was also found to cause a robust photoresponse to both monochromatic light pulses and allowed asymmetry with respect to their light-induced activity (
Figure 6 and
Figure 8) to be more easily observed. Interestingly, the crayfish
P. clarkii acclimated at a lower temperature, and the photoresponses induced by white light and recorded at 15° C from the CPR were higher than in caudal photoreceptors from the CPR from animals acclimated to 25 °C [
45]. These asymmetric photoresponses here reported from the CPRs resemble those observed with white light pulses [
33]. We carried out our studies at 14 to 24 °C. The crayfish
C. quadricarinatus are found in the native environment at temperature ranges of 12 to −33 °C [
46]. Results from other neuronal models suggest multiple mechanisms to achieve thermal compensation in crustaceans [
47,
48]. However, the possible mechanisms for the effects of temperature on the CPR responses caused by both monochromatic light wavelengths are still unknown and remain for future analyses. Serpico and Frasnelli [
49] suggested that the asymmetrical function of the nervous system plays a part in learning and memory both in vertebrates and invertebrates. In the
Caenorhabditis elegants model, left–right asymmetry is occurring in single (paired) neurons in the taste sensitivity respond differently to taste cues [
26]. Moreover, monochromatic light modulates circadian rhythms through the non-visual photoreceptor pathways in different biological models [
50,
51,
52]. As aforementioned, the caudal photoreceptor acts as a circadian pacemaker in the crayfish [
1,
2,
7]. We suggest as a topic of future studies the effect of both monochromatic light wavelengths on CPRs and the circadian rhythms of crayfish.
Renart and Machens [
53] review the studies on variability: it is reduced if there is greater control of internal variables. That is, the uncontrolled variables induce a greater variability. The results of our investigation show high variability in the activity of the caudal photoreceptors, probably because we have used young animals without distinction of sex. For this study, we control age, height, and weight; but not sex. For future studies, it is suggested to explore the factorial effect of sex and age of the animal on the caudal photoreceptors activity, and their influence on the locomotor behavior.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}