1. Introduction
Different coat colors in the European rabbit (
Oryctolagus cuniculus) have been selected throughout domestication and are nowadays fixed in specific breeds. Among the various phenotypic traits, coat color dilution corresponds to an altered distribution of eumelanin and pheomelanin pigments in skin and hair [
1]. A similar coat color dilution phenotype has been observed in other mammals such as mice. Mutations within proteins encoding the melanosome transport complex were described in the myosin VA (
Myo5a) [
2], Ras-related protein (
Rab27a) [
3] and melanophilin (
Mlph) [
4] genes. In humans, patients suffering from Griscelli Syndrome (GS) exhibit a pigment dilution in their hair and skin, which, depending on the specific genetic variant, may or may not be accompanied by other important symptoms. Interestingly, mutations within
MYO5A [
5],
RAB27A [
6] and
MLPH [
7,
8] are respectively responsible for GS1 (OMIM #214450), GS2 (OMIM #607624) and GS3 (OMIM #609227). Only GS3 is characterized by hypomelanosis with no immunologic or neurologic manifestations.
Candidate gene approaches focused on these specific genes were performed in several species presenting a dilution-like coat color phenotype. The mode of inheritance of the dilution phenotype is autosomal recessive [
1]. Mutations within the
MLPH gene were identified in mice [
4], cat [
9] (OMIA 000206-9685), dog [
10,
11,
12] (OMIA 000031-9615), chicken [
13] (OMIA 001445-9031), quail [
14] (OMIA 001445-93934), American mink [
15] (OMIA 001438-452646) and cattle [
16] (OMIA 001438-452646). In rabbits, although variants have also been highlighted within the
MLPH gene, two published studies have suggested different variants as the genuine causal mutation [
17,
18]. An alternatively spliced
MLPH transcript isoform corresponding to two-exon skipping was suggested as the causal molecular mechanism for the coat color phenotype [
17]. The c.111-5C>A variant, located within intron 2 in an acceptor site for splicing, was reported as the most likely variant leading to this exon skipping [
17]. This skipping of exons 3 and 4 caused a frameshift leading to a change of two amino acids followed by a premature stop codon [
17]. A second variant, c.585delG, corresponding to a 1-bp deletion in exon 6 of the
MLPH gene has also been identified in various breeds with a dilution of their coat [
17,
18]. This variant also led to a frameshift and an altered amino acid sequence with a premature stop codon. Both variants were identified in the first study and are associated with coat color dilution in several breeds (Netherland Dwarf, Loh, Lionhead Dwarf and Blue Vienna) [
17]. The authors suggested a higher relevance of the c.111-5C>A variant although an effect of the c.585delG variant on the dilution phenotype was suggested for individuals who were not homozygous for mutated allele at c.111-5C>A [
17]. However, only the c.585delG variant was highlighted and analyzed by Fontanesi et al. [
18]. An association signal with the coat color dilution was obtained in various breeds (
n = 7) including Blue Vienna and Castor Rex [
18].
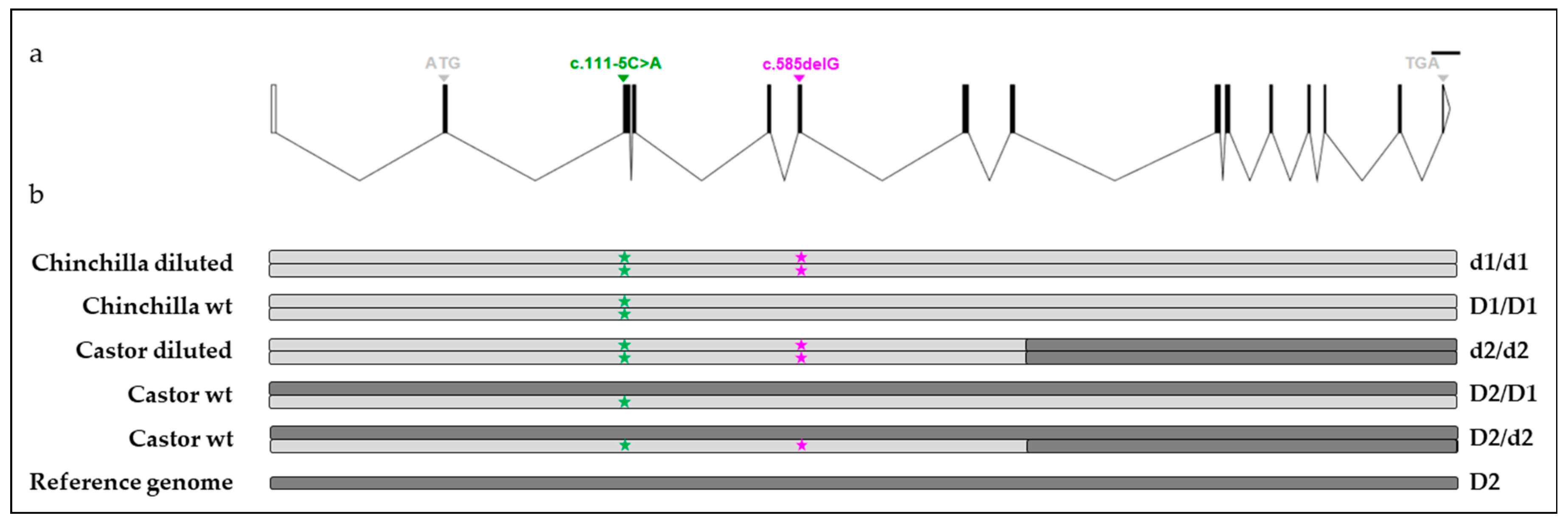
To better understand the dilution phenotype in rabbits and to likely discriminate, which of the two previously reported variants is the true causal variant, we analyzed their segregation with the dilution phenotype in Castor and Chinchilla breeds already known to have the trait of interest [
18]. Once highlighting the genetic implication of the c.585delG variant, we evaluated its impact on
MLPH transcripts in order to understand the molecular mechanisms involved in coat color dilution in rabbits.
2. Materials and Methods
2.1. Animals
The coat color dilution phenotype was observed in Castor and Chinchilla breeds (
Figure 1). Both breeds are selected for fur production; each includes two lines. The wild type (wt) Castor line has a normal black-brown coat color with a yellow agouti band. The diluted Castor line was derived from the wt Castor line by selecting the blue coat color phenotype. Similarly, the wt Chinchilla line carries a normal black-brown coat color with the Ch allele at the C locus and the other line (diluted Chinchilla) was derived from wt Chinchilla with the ash coat color phenotype. Nowadays, the four lines represent four distinct populations since they are selected independently.
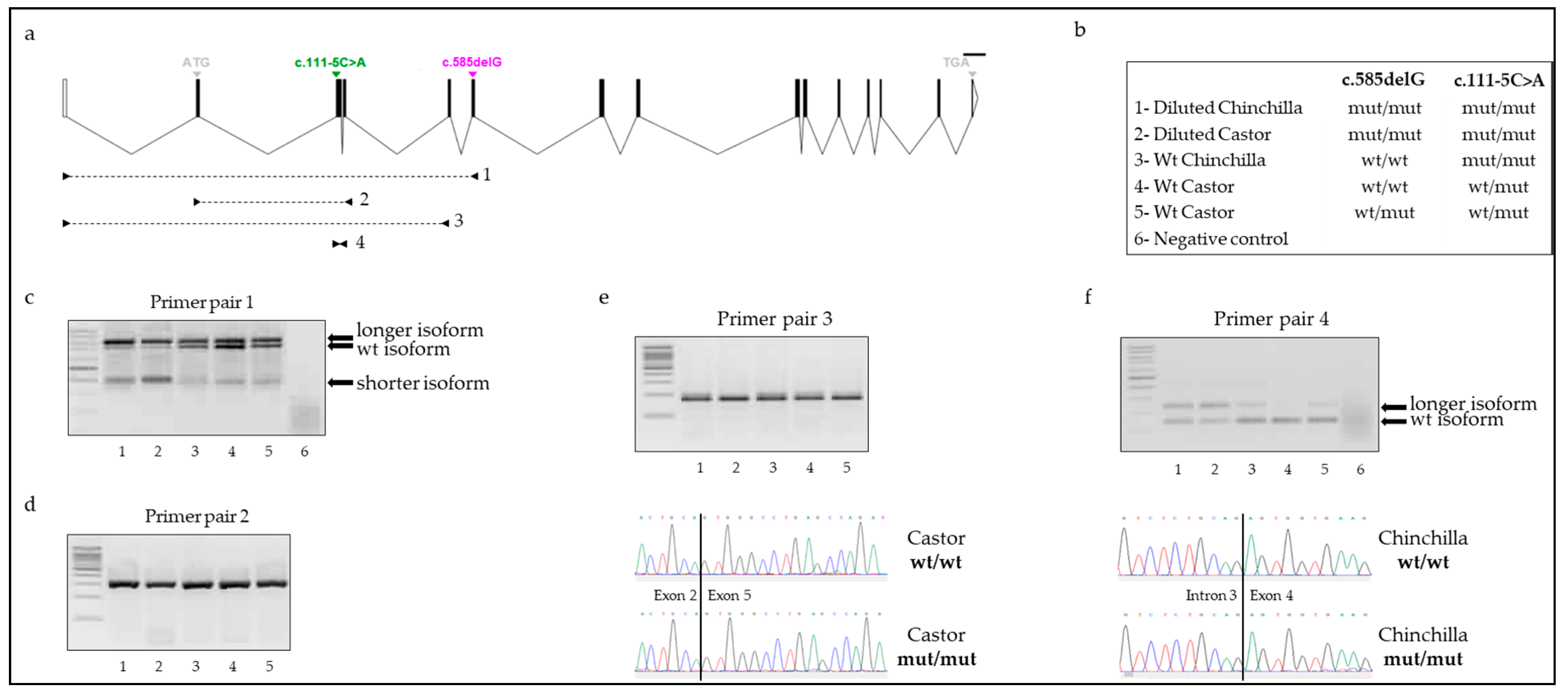
A first set of 74 rabbits including 50 Castor (37 wt vs. 13 diluted) and 24 Chinchilla (12 wt vs. 12 diluted) was used for genotyping on the genomic DNA (gDNA) and complementary DNA (cDNA) level. Moreover, 5 animals out of 74 were entirely sequenced for the MLPH gene. Among the first set, 43 rabbits were selected based on their phenotype and their genotype at the c.585delG variant to investigate MLPH transcripts. Within this selection, 17 individuals were diluted and homozygous mut/mut (8 Castor and 9 Chinchilla); 8 Castor wt animals were heterozygous wt/mut, and 18 rabbits were wt and homozygous wt/wt (8 Castor and 9 Chinchilla). Unrelated individuals were chosen.
Table S1 integrates the full information concerning animals used in this article.
2.2. Samples
All procedures were conducted in accordance with the Guide for the Care and Use of Agricultural Animals in Research and Teaching.
Skin samples were collected in 2012; no ethical code was needed at that time in France. Skin samples were collected at 3 months of age for all rabbits. Skin punch biopsies (28 mm2) were taken from the breast by an expert technician. Genomic DNA and total RNA were extracted from the same biopsy using the AllPrep DNA/RNA Mini kit according to the manufacturer’s protocol (Qiagen, Venlo, Netherlands). RNA samples were DNAse-treated to avoid gDNA contamination and diluted at 50 ng/µL in RNAse-free water. RNA (1 µg) was reverse-transcribed using Superscript II reverse transcriptase (Invitrogen, Cergy-Pontoise, France) and random primers (Nonamers, Sigma-Aldrich, St. Louis, MO, USA). Synthetised cDNA was used for real-time quantitative PCR.
2.3. Amplification of MLPH gDNA and cDNA for Sequencing
Nineteen PCR primers pairs [
18] were used for sequencing
MLPH gDNA. Briefly, PCR was carried out using a 2720 thermal cycler (Life Technologies, Carlsbad, CA, USA) in a 22-µL reaction volume containing 20 ng gDNA, 0.5 U GoTaq DNA polymerase (Promega, Madison, WI, USA), 1X GoTaq PCR buffer, 0.2 mm dNTPs, 0.4 µm of each primer and 1.5 mm MgCl
2. The PCR profile was as follows: 5 min at 95 °C, 35 amplification cycles of 30 s at 95 °C, 30 s at 62 °C or 64 °C, 30 s at 72 °C, 10 min at 72 °C.
Five additional primers pairs were used (
Table S2) and were designed on the rabbit
MLPH gene sequence (ENSOCUG00000016496) with Primer3plus software (
http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plusAbout.cgi). Two Long Range PCRs were carried out using a 2720 thermal cycler (Life Technologies) in a 23-µL reaction volume containing 100 ng gDNA, 1X GoTaq Long PCR Master Mix (Promega), 0.2 mm dNTPs, 0.4 µm of each primer and 1.5 mm MgCl
2. The PCR profile was as follows: 2 min at 94 °C, 35 amplification cycles of 20 s at 94 °C, 5 min at 68 °C, 10 min at 72 °C. Three other pairs were used for sequencing
MLPH cDNA. PCR was carried out using a 2720 thermal cycler (Life Technologies) with the GC-Rich PCR System (Roche, Basel, Switzerland) in a 50-µL reaction volume containing 5 µL cDNA dilution in 1/5, 2 U GC-rich enzyme, 1X buffer, 0.2 mm dNTPs, 0.4 µm of each primer and 1.5 mm MgCl
2. The PCR profile was as follows: 5 min at 95 °C, 45 amplification cycles of 30 s at 95 °C, 1 min at 60 °C or 62 °C, 30 s at 72 °C, 10 min at 72 °C.
Sequencing of PCR products was performed using an ABI Big Dye terminator v3.1 sequencing kit (Life Technologies) as recommended by the manufacturer’s protocol. Briefly, excess of PCR primers was removed via the treatment of PCR products with 10 U Exonuclease (New England Biolabs, Ipswich, MA, USA) and 0.5 U Thermo Sensitive Alkaline Phosphatase (Promega) for 45 min at 37 °C and 30 min at 80 °C. Purified PCR amplicons were then sequenced using the ABI Big Dye terminator v3.1 sequencing kit (Life Technologies,) and purified again with the Sephadex™ G-50 Superfine (GE Healthcare, Chicago, IL, USA) before being analysed on an automated ABI 3730 capillary sequencer (Life Technologies). Electropherograms were analysed with ChromasPro software (Technelysium, South Brisbane, Australia).
2.4. Genotyping Analyses
2.4.1. Genotyping of the c.585delG Variant on gDNA
Allele specific PCR was used for genotyping the c.585delG variant. Primers used (
Table S2) were designed on the rabbit
MLPH gene sequence (ENSOCUG00000016496) with Primer3plus software (
http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plusAbout.cgi). PCR was carried out using a 2720 thermal cycler (Life Technologies) in a 12-µL reaction volume containing 20 ng gDNA, 0.5 U GoTaq DNA polymerase (Promega), 1X GoTaq PCR buffer, 0.2 mm dNTPs, 0.4 µm of each primer and 1.5 mm MgCl
2. The PCR profile was as follows: 5 min at 95 °C, 35 amplification cycles of 30 s at 95 °C, 30 s at 66 °C for the mutated allele or 68 °C for the wild type allele, 30 s at 72 °C, 10 min at 72 °C. PCR products were electrophoresed on 2% agarose gel and visualized with bromide of ethidium.
2.4.2. Genotyping of the c.111-5C>A Variant on gDNA
2.4.3. Genotyping of the c.585delG Variant on cDNA
The c.585delG variant was analyzed on cDNA using Restriction Fragment Length Polymorphism (RFLP). The primers used (
Table S2) were designed on the rabbit
MLPH gene sequence (ENSOCUG00000016496) with Primer3plus software (
http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plusAbout.cgi). PCR was carried out using a 2720 thermal cycler (Life Technologies) with the GC-Rich PCR System (Roche) in a 25-µL reaction volume containing 2.5 µL cDNA dilution in 1/5, 1 U GC-rich enzyme, 1X buffer, 0.2 mm dNTPs, 0.4 µm of each primer and 1.5 mm MgCl
2. The PCR profile was as follows: 5 min at 95 °C, 45 amplification cycles of 30 s at 95 °C, 30 s at 56 °C, 30 s at 72 °C, 10 min at 72 °C. Digestion was performed with SmlI enzyme (New England Biolabs) in a 12-µL reaction volume containing 4 µL PCR product, 1 U enzyme, 1X buffer at 55 °C overnight. The digestion products were electrophoresed on 2% agarose gel and were visualized with bromide of ethidium. In animals that were homozygous wt/wt for the c.585delG variant, a 173-bp undigested fragment was expected, while in rabbits homozygous mut/mut, digestion by SmlI was suggested, leading to two fragments of sizes 123 and 50 bp.
2.5. Stability and Accumulation of MLPH Transcripts
Primers are listed in
Table S2. Real-time quantitative PCR analysis was carried out with a LightCycler
® 480 System using the SYBR Green I Master (Roche Life Science, Indianapolis, IN, USA) as recommended by the manufacturer. Briefly, 3 µL of diluted cDNA (1/5) were mixed with 0.15 µM of each primer and 1X Master volume in a final volume of 10 µL, and a standard qPCR amplification was then performed as follows: 45 cycles of 15 s at 95 °C, 15 s at 60 °C and 15 s at 72 °C. Each qPCR reaction was done in duplicate.
A final melting curve analysis was systematically performed for each primer pair to check for the presence of only one PCR product peak, and each amplicon was sequenced to verify the amplification specificity (as explained previously). The baseline adjustment method of the LightCycler 480 software (Roche Diagnostic, Rotkreuz, Switzerland) was used to determine the Ct (Cycle threshold) in each reaction. Primer efficiency curves were generated using five serial dilutions of cDNA (two-fold dilution from 1:5 to 1:80) on the abscissa and the corresponding Ct on the ordinate. The slope of the log-linear phase reflects the amplification efficiency (E) derived from the formula E = e(−1/slope). For quantification analysis, the Ct of target gene was compared with the mean of the expression of the internal reference gene HPRT encoding a ubiquitous hypoxanthine guanine phosphoribosyl transferase according to the ratio R = 100 × (EReferenceCt Reference/ETargetCt Target). Results were expressed as the quantify relative to the mean of the housekeeping gene expression. The significance of results was tested using a t-test for the comparison of the genotypic clades.
4. Discussion
Dilution of coat color is a trait of interest in many species. Variants within the melanophilin (
MLPH) gene have been identified in dogs [
10,
11,
12], mink [
15] and cattle [
16]. In humans,
MLPH variants are responsible for GS3 (OMIM #609227) [
7,
8]. In rabbits, two different variants were suggested as potentially causal for the dilution phenotype. The first described variant [
17], c.111-5C>A, was located within the second intron and resulted in a two-exon skipping
MLPH transcript. This skipping of exons 3 and 4 caused a frameshift leading to a change of two amino acids followed by a premature stop codon at the fortieth amino acid of the protein [
17]. The second variant [
17,
18], c.585delG, corresponded to a 1-bp deletion within the sixth exon of the
MLPH gene, leading to a premature STOP codon at the 360th amino acid of the protein. To discriminate between the two variants and evaluate the effect on the stability and accumulation of
MLPH transcripts, we performed both genetic and functional studies focused on
MLPH in rabbits from the Castor and Chinchilla breeds in which the dilution phenotype segregated.
We managed to sequence the whole
MLPH sequence from exons 1 to 15. This wt
MLPH sequence obtained from both Castor and Chinchilla animals was similar to NM_001297485.1 [
17] obtained from a black Loh rabbit. The rabbit MLPH protein seemed shorter with 562 amino acids (NP_001284414) compared to both human and mouse proteins consisting of 600 (NP_077006) and 590 (NP_443748) amino acids, respectively. While the beginning (amino acids 1 to 210) and the end (amino acids 460 to 560) of the protein are homologous to human and mice proteins, the middle of the protein is quite different in rabbit. Indeed, the Myosin VA binding domain appeared very different across species compared to Rab27A and Actin binding domains that seemed very well conserved among rabbit, human and mice.
The genetic analysis performed in Castor and Chinchilla breeds likely excluded the c.111-5C>A variant [
17] as the causal mutation of the dilution phenotype. Indeed, some animals which were homozygous mut/mut at this variant and homozygous wt/wt or heterozygous at the c.585delG variant had a wt phenotype which is incompatible with the postulated causal role of this variant in an autosomal recessive mode of inheritance with full penetrance of the coat color dilution [
1]. Moreover, only the c.585delG variant perfectly discriminated wt (D1) from diluted (d1) haplotypes in the Chinchilla breed as observed in [
18] between the most frequent wt haplotype and diluted haplotype obtained from the
MLPH sequencing of 18 rabbits of different coat color. While a few Castor individuals were analyzed by Fontanesi et al. [
18], neither Castor nor Chinchilla rabbits were genotyped by Lehner et al. [
17]. Different variants within the
MLPH gene associated with the dilution phenotype would be one hypothesis explaining the discrepancy between our results and those of [
17]. This was observed in sheep for various
BMP15 mutations that were all responsible for sterility in homozygous ewes [
19,
20,
21,
22,
23] or in cattle for mutations in the myostatin gene (
MSTN), which were all associated with hypermuscularity [
24]. However, all diluted haplotypes identified from six different rabbits including one Chinchilla, one Castor, one Vienna Blue [
18], one Checkered Giant [
18] and two F1 Checkered Giant [
18] were similar for all common positions. These results strongly suggested that in the investigated rabbit breeds a single
MLPH variant influences coat color dilution instead of different variants within the
MLPH gene, each specific to a breed. The 1-bp deletion in exon 6 of the
MLPH gene might be the causal variant for the dilution phenotype in Castor and Chinchilla breeds. The same variant in both populations suggested a common origin with likely an introgression of Chinchilla into Castor given the shorter size of the Castor haplotype carrying the deletion. To go into further details about the origin of the rabbit dilute allele, it would be interesting to determine the shortest diluted haplotype segregating in breeds of rabbits with a dilution of their coat color.
Fancy breeders suggest the existence of two types of dilution: one that gives a darker grey (or blue), as observed in 2 rabbits [
17], and another one that gives a lighter grey (or blue) colour. While both variants (c.111-5C>A and c.585delG variant) were identified in [
17], authors suggested the c.111-5C>A as the most probable candidate due to the
p-value of the trend test and the early position in the protein of a premature stop codon although the c.585delG variant showed the best association signal from both genotypic and allelic tests [
17]. However, none of the two variants segregated perfectly with the dilution phenotype in their various populations. They finally suggested a combination of both variants to influence coat color dilution. In Castor and Chinchilla breeds, the c.111-5C>A variant and the c.585delG variant were in total linkage disequilibrium in diluted animals since all diluted rabbits of both breeds were homozygous mut/mut at both variant positions. The identification of other individuals mut/mut at the c.585delG variant position but homozygous wt/wt or heterozygous at the c.111-5C>A variant position might provide new insights whether c.111-5C>A has a modifying effect in the Castor and Chinchilla breeds. However, only one dilution phenotype was observed in our study, likely rejecting the involvement of the
MLPH gene in the degree of dilution (dark vs. light dilution). The most probable hypothesis would be that the degree of dilution might result from interactions between the 1-bp deletion at the c.585delG variant, responsible for the coat color dilution, and other coat color loci.
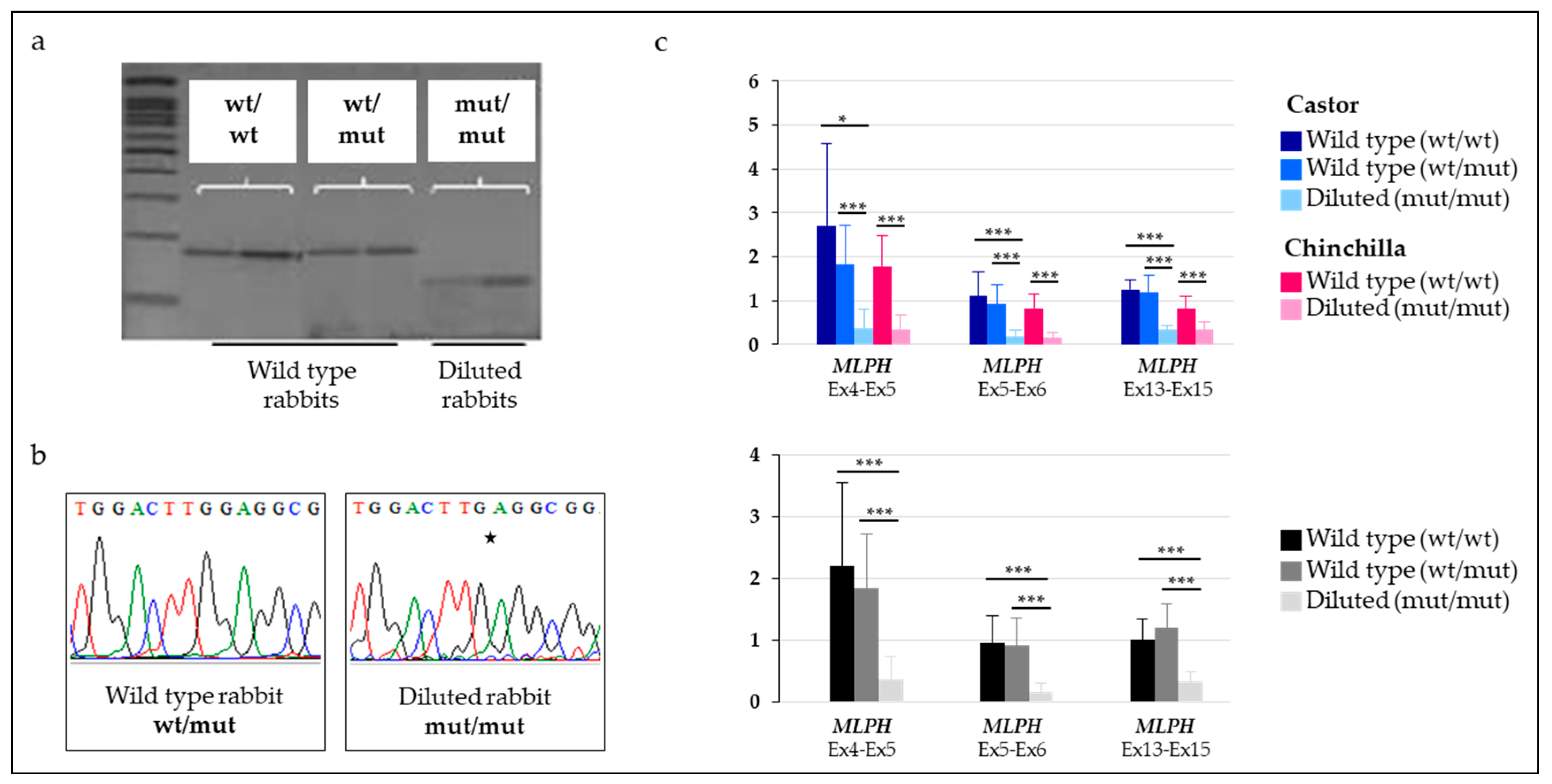
Knowledge of the molecular mechanisms involved in the dilution phenotype is limited. Assuming that the c.585delG variant was the causal mutation affecting the trait of interest, we followed-up by analyzing the impact of the deletion on MLPH transcripts. Our data suggested a production of various abnormal MLPH transcripts in both wt and diluted rabbits. Three distinct isoforms of MLPH were identified in skin samples of animals: the wild type MLPH transcript, a shorter MLPH transcript corresponding to the two-exon skipping isoform and a longer MLPH transcript. Sequencing of this latest isoform identified a transcript with retention of intron 3 between exons 3 and 4. However, no variant was detected in the splice donor splicing or splice acceptor splicing site, which might potentially explain the retention. In both human and mouse species, various retained intron MLPH isoforms (ENST00000477457.1, ENST00000495439.5, ENSMUST00000136220.1) do exist, involving introns much longer than the 92-bp of intron 3 of rabbit. Our results are thus concordant with what was observed in other mammals. The qualitative presence of the exon skipping and retained intron isoforms did not seem to differ between wt and diluted rabbits. Nevertheless, the quantity seemed variable with a stronger amplification of both shorter and longer MLPH isoforms in individuals with a dilution of their coat color and homozygous mut/mut at both c.111-5C>A and c.585delG variants. Our data did not allow us to conclude about the direct or additional effect of the two-exon skipping on the coat color dilution or c.111-5C>A as the putative causal variant for this exon skipping since we did not investigate individuals, which were wt/wt for the c.111-5C>A variant. An in-depth analysis performed on more animals representative of the nine groups of genotypes at both variants would help to better appreciate the relative quantification of each isoform.
Finally, we showed that
MLPH transcripts carrying the deletion were undetectable in rabbits heterozygous at the c.585delG variant while this isoform was detectable in homozygous mut/mut individuals. As expected, this result strongly suggested that the deleted
MLPH isoform, containing a premature stop codon, went through the nonsense mediated mRNA decay (NMD) process and might be degraded. This was shown, in a proteomics experiment, for a human
MLPH isoform (accession numbers ENST00000432475.5 and H7C052). A way to test this hypothesis would be to measure the quantity of key factors of this molecular mechanism such as up-frameshift (UPF) members [
25]. In consequence, we demonstrated that the overall expression of
MLPH transcripts, including the different isoforms, was drastically decreased in animals carrying the 1-bp deletion at the c.585delG variant and with a coat color dilution. Our results are in concordance with those obtained for other species including cats [
9] and dogs [
11]. Variants located within the
MLPH gene and associated with a dilution phenotype were predicted to alter the splicing or the protein. In dogs, quantitative PCR showed that diluted animals had only about 25% of the
MLPH transcript compared with wild type animals [
11].
Together, our results bring new insights into the dilution of coat color in rabbits. The c.585delG variant remains as the most likely causative variant for coat color dilution in multiple rabbit breeds. This frameshift deletion may drive the produced MLPH transcript to degradation and prevent a full-length functional MLPH protein. Additional MLPH variants including the c.111-5C>A variant may also lead to abnormal MLPH isoforms (containing premature stop codon) such as exon skipping and intron retention transcripts. The production of a non-functional MLPH protein from the various MLPH isoforms might contribute to modifying effects on coat color by affecting melanosome transport within pigmented tissues.






{kind=link}
{kind=link}
{kind=link}
{kind=link}