Effects of Emerging Infectious Diseases on Amphibians: A Review of Experimental Studies

 , , , , , , and
, , , , , , and
Abstract
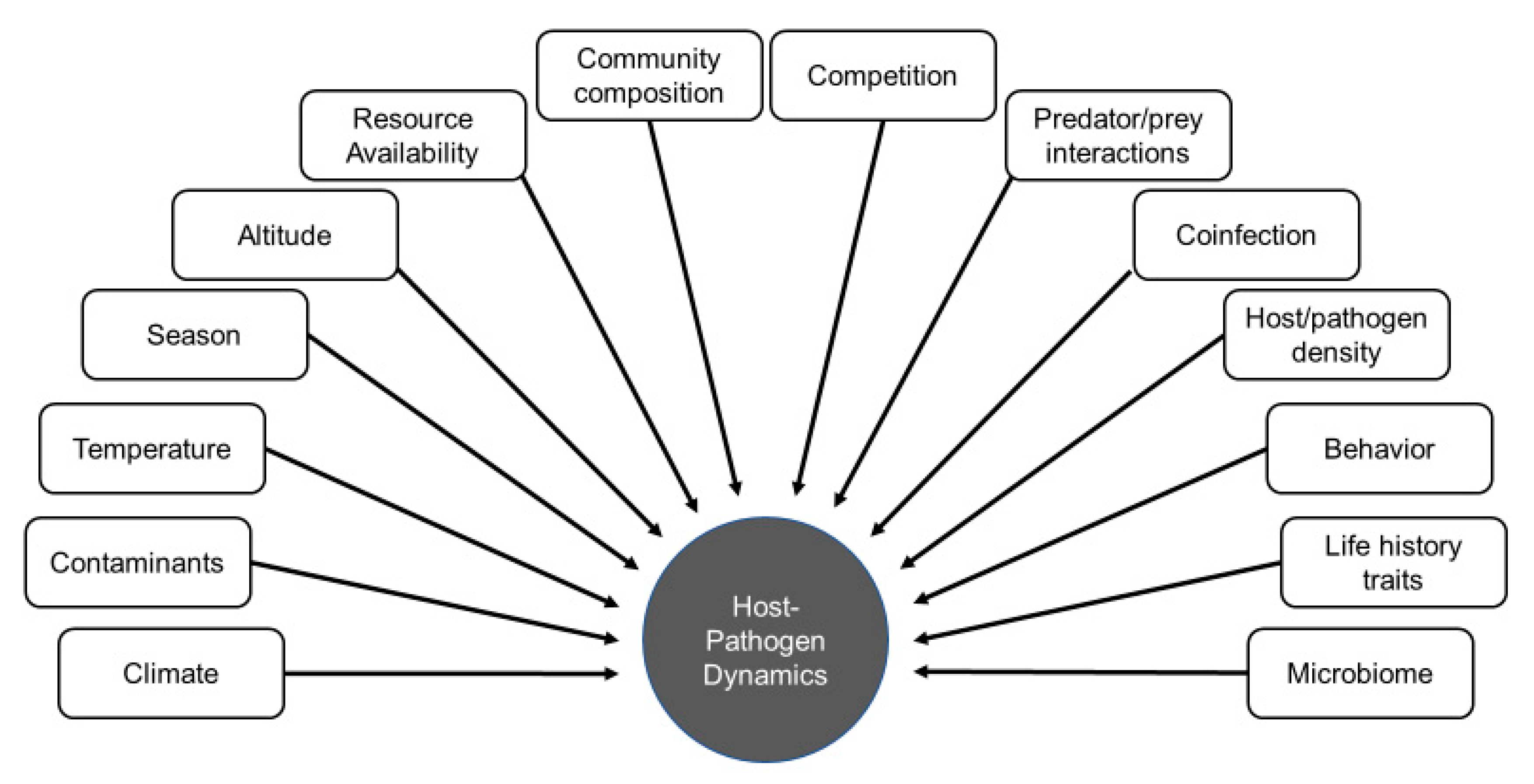
:1. Introduction
2. Methods
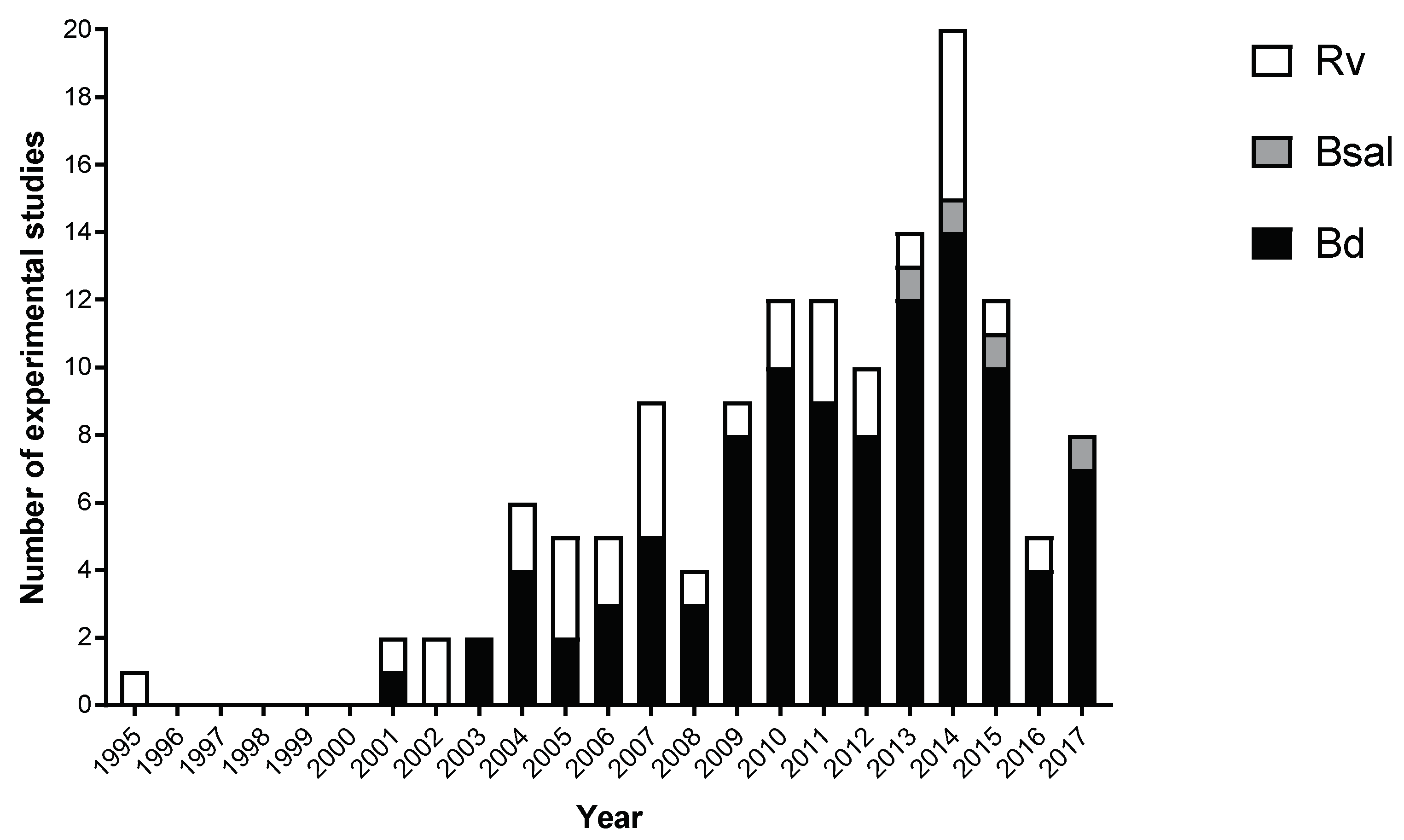
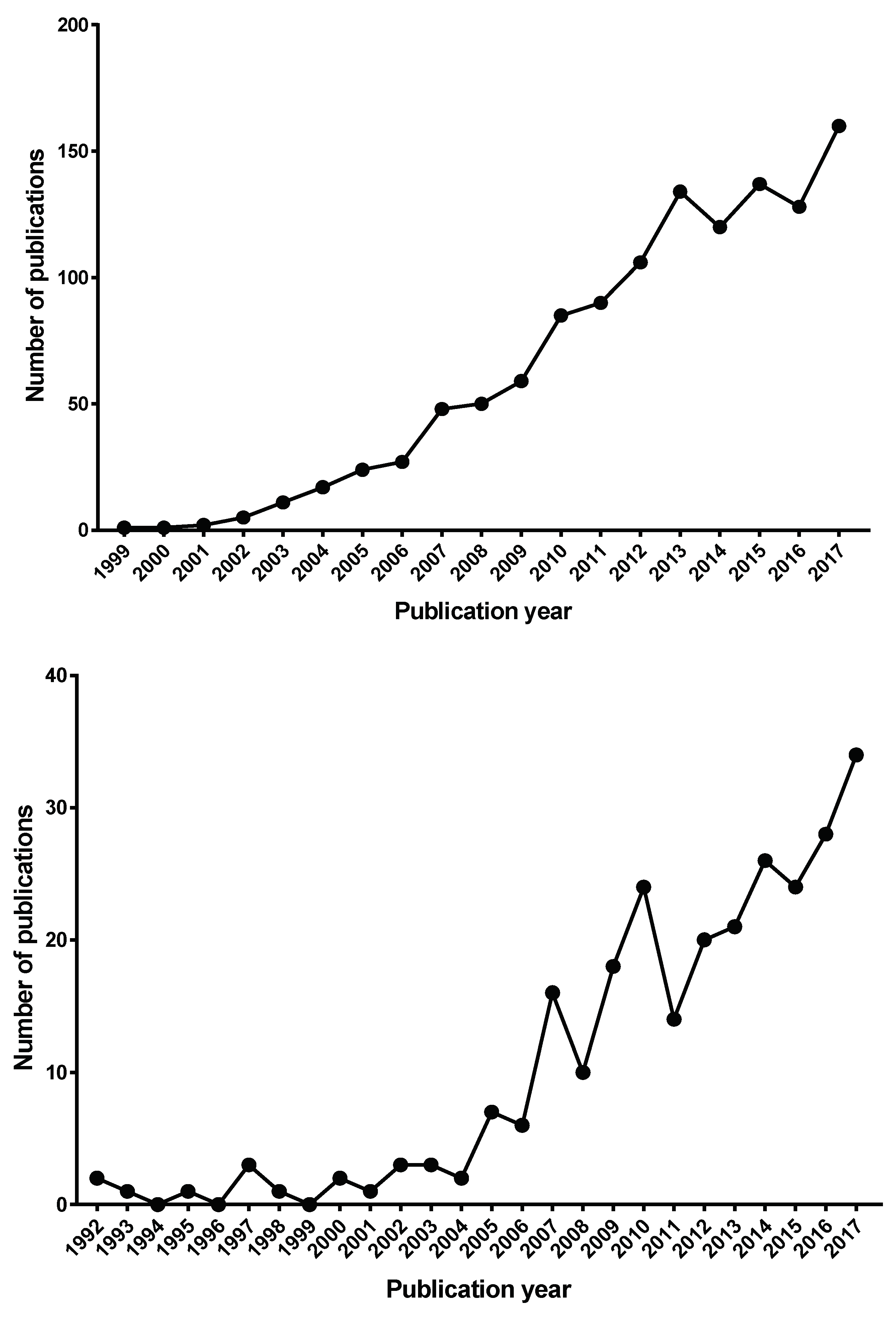
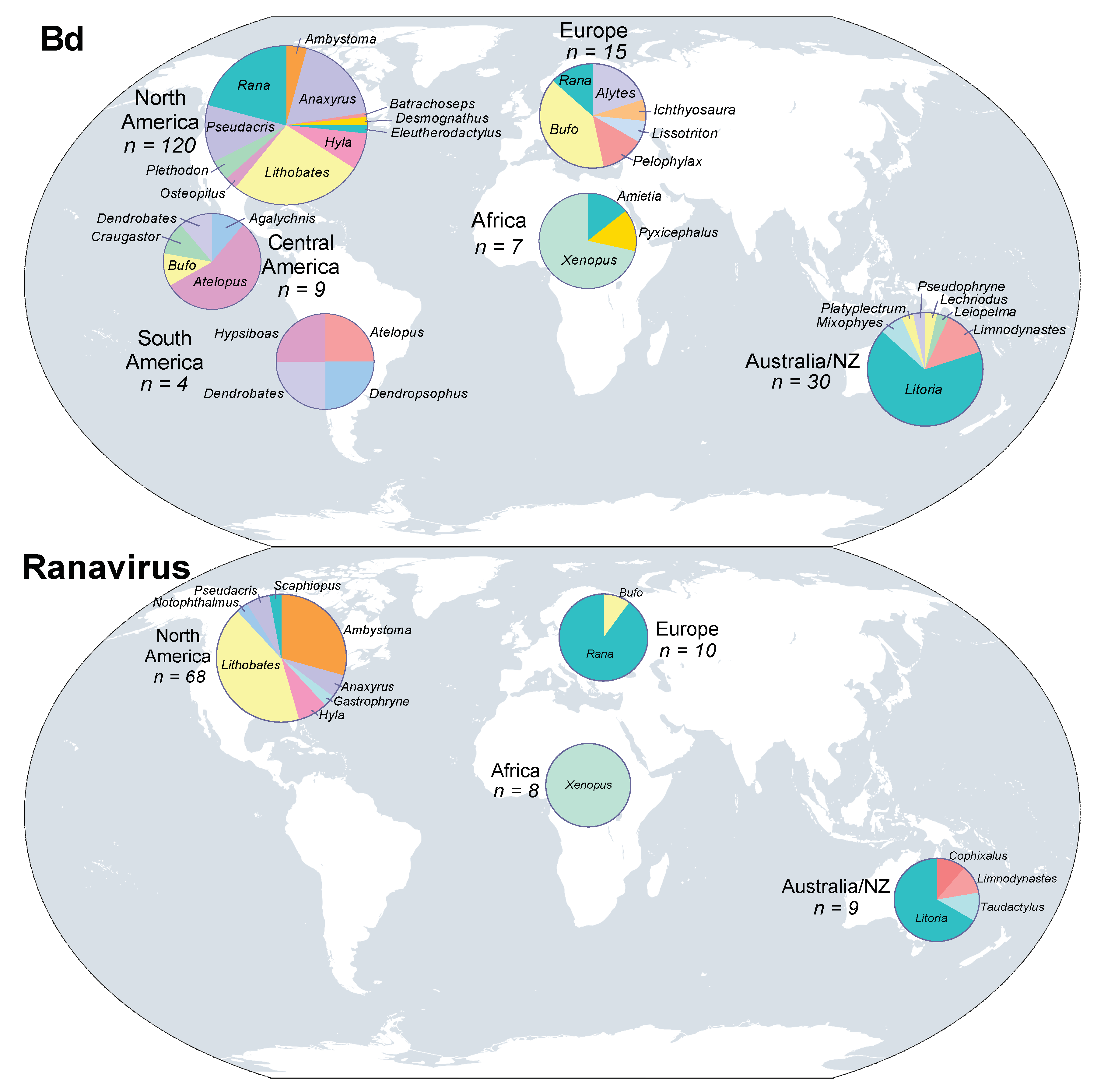
3. Results
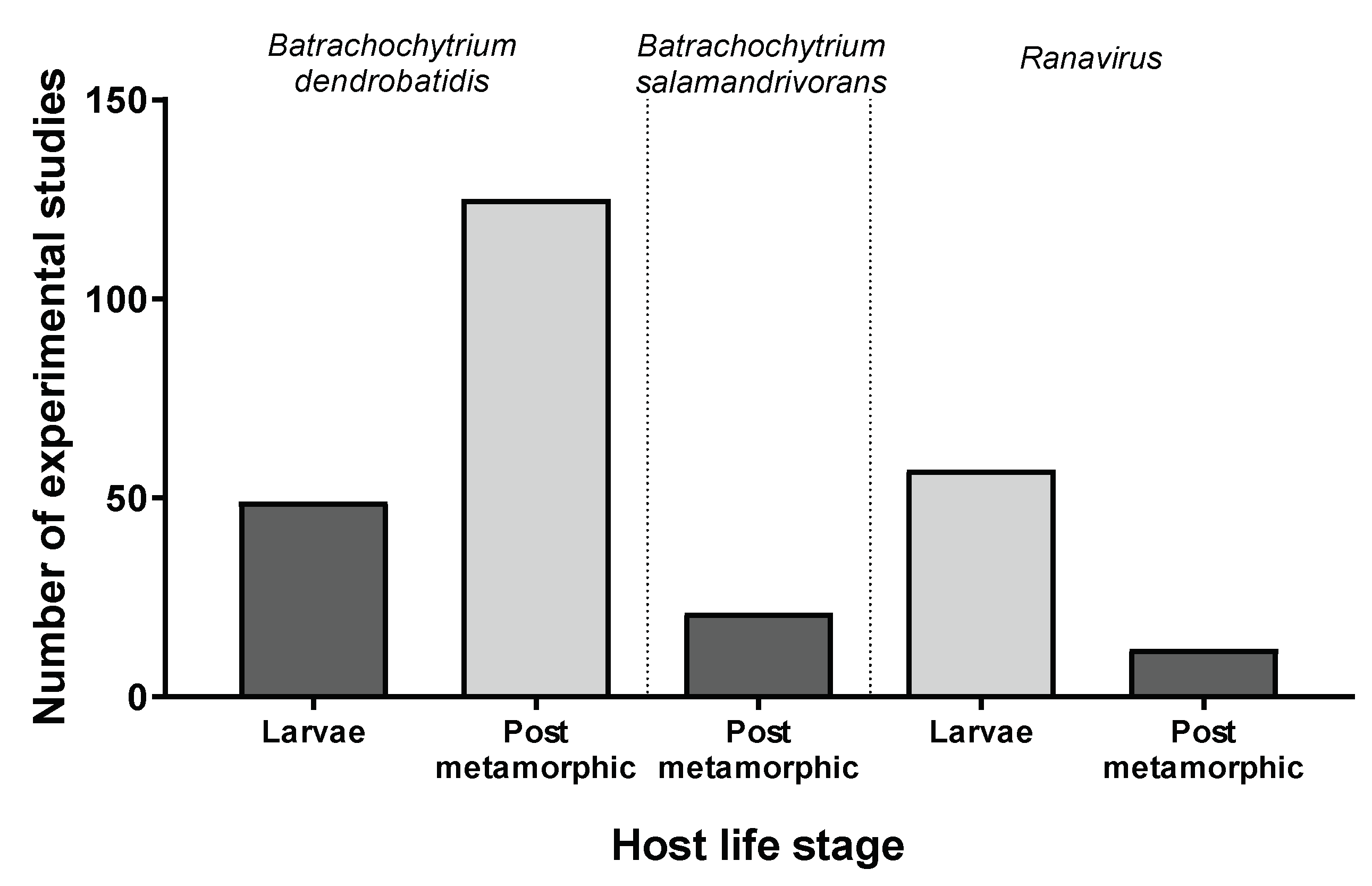
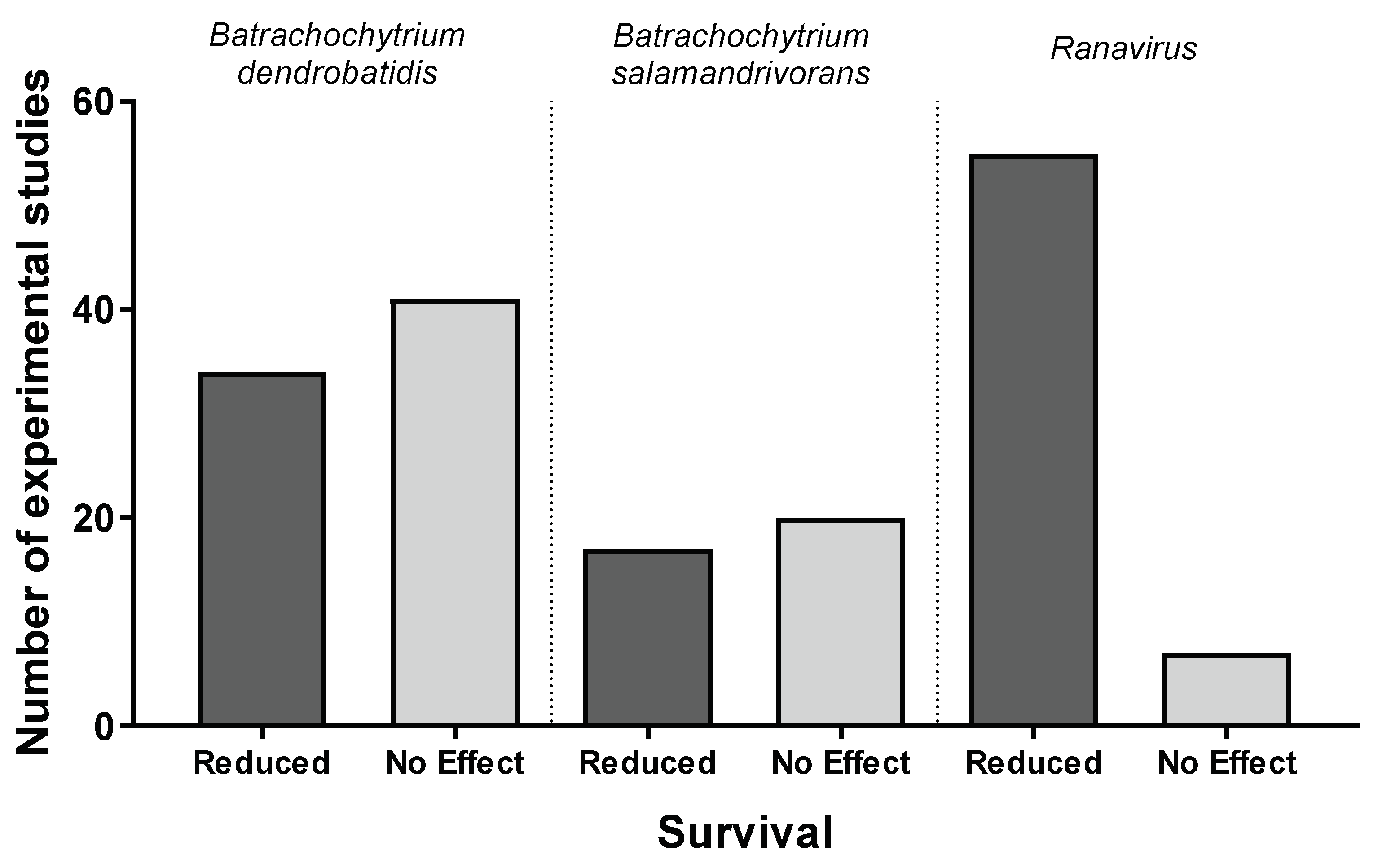
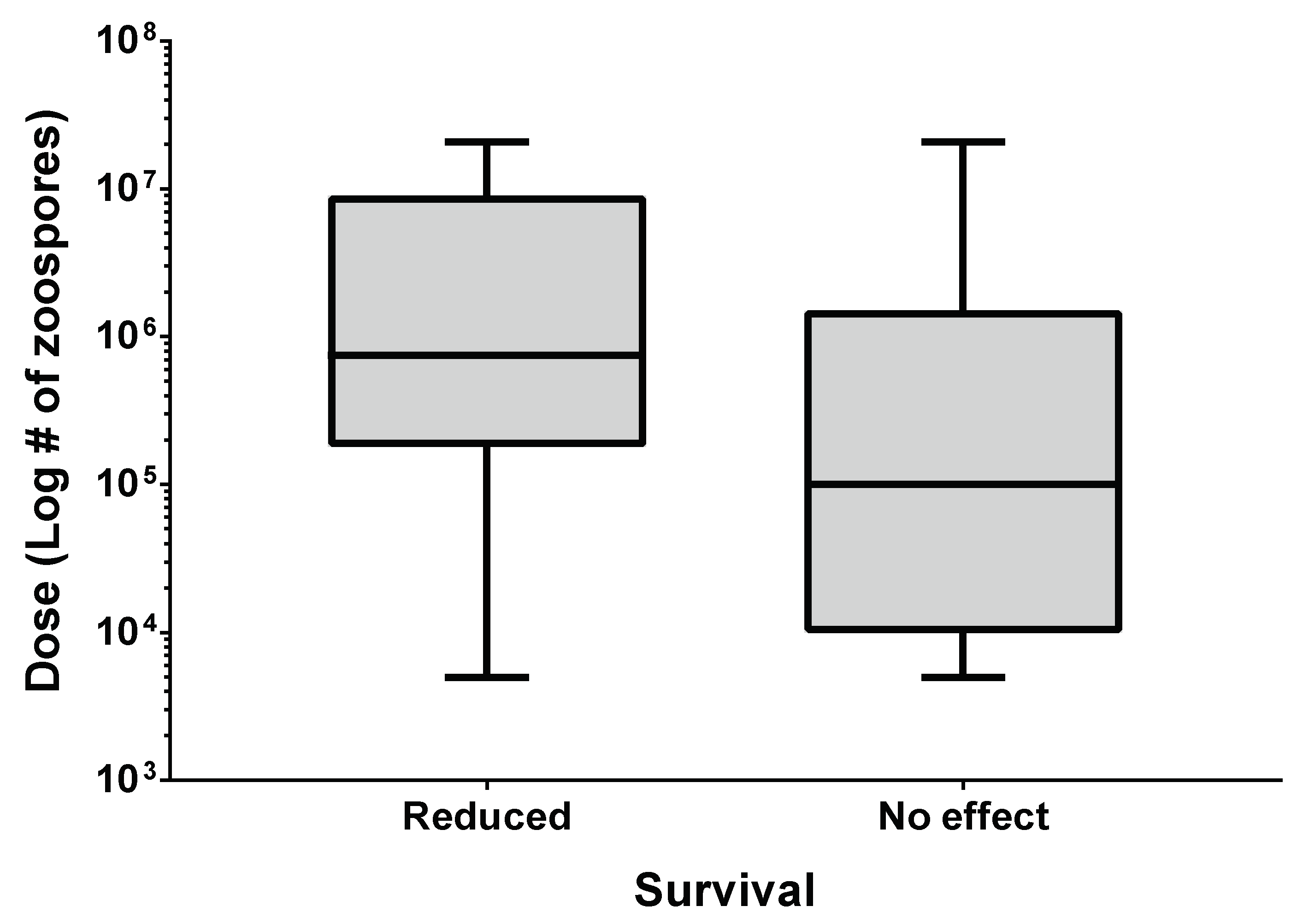
3.1. Batrachochytrium dendrobatidis
3.2. Batrachochytrium salamandrivorans
3.3. Ranavirus
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Pimm, S.L.; Russell, G.J.; Gittleman, J.L.; Brooks, T.M. The future of biodiversity. Science 1995, 269, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.O. The effects of complex social life on evolution and biodiversity. Oikos 1992, 63, 13–18. [Google Scholar] [CrossRef]
- Alroy, J. Current extinction rates of reptiles and amphibians. Proc. Natl. Acad. Sci. USA 2015, 112, 13003–13008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, A.R.; Han, B.A.; Relyea, R.A.; Johnson, P.T.J.; Buck, J.C.; Gervasi, S.S.; Kats, L.B. The complexity of amphibian population declines: Understanding the role of cofactors in driving amphibian losses. Ann. N. Y. Acad. Sci. 2011, 1223, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Alford, R.A.; Richards, S.J. Global amphibian declines: A Problem in applied ecology. Annu. Rev. Ecol. Syst. 1999, 30, 133–165. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Romansic, J.M.; Kiesecker, J.M.; Hatch, A.C. Ultraviolet radiation, toxic chemicals and amphibian population declines. Divers. Distrib. 2003, 9, 123–140. [Google Scholar] [CrossRef]
- Muths, E.; Scherer, R.D.; Corn, P.S.; Lambert, B.A. Estimation of temporary emigration in male toads. Ecology 2006, 87, 1048–1056. [Google Scholar] [CrossRef]
- Daszak, P.; Berger, L.; Cunningham, A.A.; Hyatt, A.D.; Green, D.E.; Speare, R. Emerging infectious diseases and amphibian population declines. Emerg. Infect. Dis. 1999, 5, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife—Threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Bosch, J.; Yin, Z.; Stead, D.A.; Walker, J.; Selway, L.; Brown, A.J.P.; Walker, L.A.; Gow, N.A.R.; Stajich, J.E.; et al. Proteomic and phenotypic profiling of the amphibian pathogen Batrachochytrium dendrobatidis shows that genotype is linked to virulence. Mol. Ecol. 2009, 18, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Olson, D.H.; Aanensen, D.M.; Ronnenberg, K.L.; Powell, C.I.; Walker, S.F.; Bielby, J.; Garner, T.W.J.; Weaver, G.; The Bd Mapping Group; Fisher, M.C. Mapping the global emergence of Batrachochytrium dendrobatidis, the amphibian chytrid fungus. PLoS ONE 2013, 8, e56802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skerratt, L.; Berger, L.; Speare, R.; Cashins, S.; McDonald, K.; Phillott, A.; Hines, H.; Kenyon, N. Spread of chytridiomycosis has caused the rapid global decline and extinction of frogs. EcoHealth 2007, 4, 125–134. [Google Scholar] [CrossRef]
- Martel, A.; Spitzen-van der Sluijs, A.; Blooi, M.; Bert, W.; Ducatelle, R.; Fisher, M.C.; Woeltjes, A.; Bosman, W.; Chiers, K.; Bossuyt, F.; et al. Batrachochytrium salamandrivorans sp. nov. causes lethal chytridiomycosis in amphibians. Proc. Natl. Acad. Sci. USA 2013, 110, 15325–15329. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G.; Hyatt, A.; Miyazaki, T.; Williams, T. Family iridoviridae: Poor viral relations no longer. In Lesser Known Large dsDNA Viruses; Van Etten, J.L., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 123–170. ISBN 978-3-540-68618-7. [Google Scholar]
- Kik, M.; Martel, A.; der Sluijs, A.S.; Pasmans, F.; Wohlsein, P.; Gröne, A.; Rijks, J.M. Ranavirus-associated mass mortality in wild amphibians, The Netherlands, 2010: A first report. Vet. J. 2011, 190, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Miaud, C.; Dejean, T.; Savard, K.; Millery-Vigues, A.; Valentini, A.; Curt Grand Gaudin, N.; Garner, T.W.J. Invasive North American bullfrogs transmit lethal fungus Batrachochytrium dendrobatidis infections to native amphibian host species. Biol. Invasions 2016, 18, 2299–2308. [Google Scholar] [CrossRef]
- Teacher, A.G.F.; Cunningham, A.A.; Garner, T.W.J. Assessing the long-term impact of Ranavirus infection in wild common frog populations. Anim. Conserv. 2010, 13, 514–522. [Google Scholar] [CrossRef]
- Green, D.E.; Converse, K.A.; Schrader, A.K. Epizootiology of sixty-four amphibian morbidity and mortality events in the USA, 1996–2001. Ann. N. Y. Acad. Sci. 2002, 969, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, A.R. Chicken Little or Nero’s Fiddle? A perspective on declining amphibian populations. Herpetologica 1994, 50, 85–97. [Google Scholar]
- Cunningham, A.A.; Langton, T.E.; Bennet, P.M.; Lewin, J.F.; Drury, S.N.; Gough, R.E.; Macgregor, S.K. Pathological and microbiological findings from incidents of unusual mortality of the common frog Rana temporaria. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1539–1557. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; Lunde, K.B.; Thurman, E.M.; Ritchie, E.G.; Wray, S.N.; Sutherland, D.R.; Kapfer, J.M.; Frest, T.J.; Bowerman, J.; Blaustein, A.R. Parasite (Ribeiroia ondatrae) infection linked to amphibian malformations in the western United States. Ecol. Monogr. 2002, 72, 151–168. [Google Scholar] [CrossRef]
- Worthylake, K.M.; Hovingh, P. Mass mortality of salamanders (Ambystoma tigrinum) by bacteria (Acinetobacter) in an oligotrophic seepage mountain lake. Great Basin Nat. 1989, 49, 364–372. [Google Scholar] [CrossRef]
- Blaustein, A.; Alford, R.; Harris, R. The value of well-designed experiments in studying diseases with special reference to amphibians. EcoHealth 2009, 6, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Longcore, J.E.; Pessier, A.P.; Nichols, D.K. Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia 1999, 91, 219–227. [Google Scholar] [CrossRef]
- Berger, L.; Speare, R.; Daszak, P.; Green, D.E.; Cunningham, A.A.; Goggin, C.L.; Slocombe, R.; Ragan, M.A.; Hyatt, A.D.; McDonald, K.R.; et al. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc. Natl. Acad. Sci. USA 1998, 95, 9031–9036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lips, K.R. Decline of a tropical montane amphibian fauna. Conserv. Biol. 1998, 12, 106–117. [Google Scholar] [CrossRef]
- McCallum, H. Inconclusiveness of chytridiomycosis as the agent in widespread frog declines. Conserv. Biol. 2005, 19, 1421–1430. [Google Scholar] [CrossRef]
- O’Hanlon, S.J.; Rieux, A.; Farrer, R.A.; Rosa, G.M.; Waldman, B.; Bataille, A.; Kosch, T.A.; Murray, K.A.; Brankovics, B.; Fumagalli, M.; et al. Recent Asian origin of chytrid fungi causing global amphibian declines. Science 2018, 360, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.; Marantelli, G.; Skerratt, L.F.; Speare, R. Virulence of the amphibian chytrid fungus Batrachochytrium dendrobatidis varies with the strain. Dis. Aquat. Org. 2005, 68, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, S.E.; Calhoun, A.J.K.; Longcore, J.E.; Levy, M.G. Transmission of Batrachochytrium dendrobatidis to wood frogs (Lithobates sylvaticus) via a bullfrog (L. catesbeianus) vector. J. Wildl. Dis. 2012, 48, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.K.; Lamirande, E.W.; Pessier, A.P.; Longcore, J.E. Experimental transmission of cutaneous chytridiomycosis in dendrobatid frogs. J. Wildl. Dis. 2001, 37, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Voyles, J.; Young, S.; Berger, L.; Campbell, C.; Voyles, W.F.; Dinudom, A.; Cook, D.; Webb, R.; Alford, R.A.; Skerratt, L.F.; et al. Pathogenesis of chytridiomycosis, a cause of catastrophic amphibian declines. Science 2009, 326, 582–585. [Google Scholar] [CrossRef] [PubMed]
- Voyles, J.; Berger, L.; Young, S.; Speare, R.; Webb, R.; Warner, J.; Rudd, D.; Campbell, R.; Skerratt, L.F. Electrolyte depletion and osmotic imbalance in amphibians with chytridiomycosis. Dis. Aquat. Org. 2007, 77, 113–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitzen-van der Sluijs, A.; Martel, A.; Asselberghs, J.; Bales, E.K.; Beukema, W.; Bletz, M.C.; Dalbeck, L.; Goverse, E.; Kerres, A.; Kinet, T.; et al. Expanding distribution of lethal amphibian fungus Batrachochytrium salamandrivorans in Europe. Emerg. Infect. Dis. 2016, 22, 1286–1288. [Google Scholar] [CrossRef] [PubMed]
- Spitzen-van der Sluijs, A.; Martel, A.; Hallman, C.; Bosman, W.; Garner, T.W.J.; Van Rooij, P.; Jooris, R.; Haesebrouck, F.; Pasmans, F. Environmental determinants of recent endemism of Batrachochytrium dendrobatidis infections in amphibian assemblages in the absence of disease outbreaks. Conserv. Biol. 2014, 28, 1302–1311. [Google Scholar] [CrossRef] [PubMed]
- Sabino-Pinto, J.; Bletz, M.; Hendrix, R.; Perl, R.G.B.; Martel, A.; Pasmans, F.; Lötters, S.; Mutschmann, F.; Schmeller, D.S.; Schmidt, B.R.; et al. First detection of the emerging fungal pathogen in Germany. Amphib. Reptil. 2015, 36, 411–416. [Google Scholar] [CrossRef]
- Martel, A.; Blooi, M.; Adriaensen, C.; Van Rooij, P.; Beukema, W.; Fisher, M.C.; Farrer, R.A.; Schmidt, B.R.; Tobler, U.; Goka, K.; et al. Recent introduction of a chytrid fungus endangers Western Palearctic salamanders. Science 2014, 346, 630–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bales, E.K.; Hyman, O.J.; Loudon, A.H.; Harris, R.N.; Lipps, G.; Chapman, E.; Roblee, K.; Kleopfer, J.D.; Terrell, K.A. Pathogenic chytrid fungus Batrachochytrium dendrobatidis, but not B. salamandrivorans, detected on eastern hellbenders. PLoS ONE 2015, 10, e0116405. [Google Scholar] [CrossRef] [PubMed]
- Muletz, C.; Caruso, N.M.; Fleischer, R.C.; McDiarmid, R.W.; Lips, K.R. Unexpected rarity of the pathogen Batrachochytrium dendrobatidis in Appalachian plethodon salamanders: 1957–2011. PLoS ONE 2014, 9, e103728. [Google Scholar] [CrossRef] [PubMed]
- Iwanowicz, D.D.; Schill, W.B.; Olson, D.H.; Adams, M.J.; Densmore, C.; Conman, R.; Adams, C.; Figiel, J.; Anderson, C.W.; Blaustein, A.R.; et al. Potential concerns with analytical methods used for detection of Batrachochytrium salamandrivorans from archived DNA of amphibian swab samples, Oregon, USA. Herpetol. Rev. 2017, 48, 352–355. [Google Scholar]
- Zhu, W.; Bai, C.; Wang, S.; Soto-Azat, C.; Li, X.; Liu, X.; Li, Y. Retrospective survey of museum specimens reveals historically widespread presence of Batrachochytrium dendrobatidis in China. EcoHealth 2014, 11, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Grant, E.H.C.; Muths, E.L.; Katz, R.A.; Canessa, S.; Adams, M.J.; Ballard, J.R.; Berger, L.; Briggs, C.J.; Coleman, J.; Gray, M.J.; et al. Salamander Chytrid Fungus (Batrachochytrium salamandrivorans) in the United States—Developing Research, Monitoring, and Management Strategies; Open-File Report; USGS: Reston, VA, USA, 2016; p. 26.
- Gray, M.J.; Lewis, J.P.; Nanjappa, P.; Klocke, B.; Pasmans, F.; Martel, A.; Stephen, C.; Parra Olea, G.; Smith, S.A.; Sacerdote-Velat, A.; et al. Batrachochytrium salamandrivorans: The North American response and a call for action. PLoS Pathog. 2015, 11, e1005251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richgels, K.L.D.; Russell, R.E.; Adams, M.J.; White, C.L.; Grant, E.H.C. Spatial variation in risk and consequence of Batrachochytrium salamandrivorans introduction in the USA. R. Soc. Open Sci. 2016, 3, 150616. [Google Scholar] [CrossRef] [PubMed]
- Yap, T.A.; Koo, M.S.; Ambrose, R.F.; Wake, D.B.; Vredenburg, V.T. Averting a North American biodiversity crisis. Science 2015, 349, 481–482. [Google Scholar] [CrossRef] [PubMed]
- Chinchar, V.G. Ranaviruses (family Iridoviridae): Emerging cold-blooded killers. Arch. Virol. 2002, 147, 447–470. [Google Scholar] [CrossRef] [PubMed]
- Granoff, A.; Came, P.E.; Rafferty, K.A. The isolation and properties of viruses from Rana pipiens: Their possible relationship to the renal adenocarcinoma of the leopard frog*. Ann. N. Y. Acad. Sci. 1965, 126, 237–255. [Google Scholar] [CrossRef] [PubMed]
- Lesbarrères, D.; Balseiro, A.; Brunner, J.; Chinchar, V.G.; Duffus, A.; Kerby, J.; Miller, D.L.; Robert, J.; Schock, D.M.; Waltzek, T.; et al. Ranavirus: Past, present and future. Biol. Lett. 2012, 8, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Schock, D.M.; Bollinger, T.K.; Gregory Chinchar, V.; Jancovich, J.K.; Collins, J.P. Experimental evidence that amphibian ranaviruses are multi-host pathogens. Copeia 2008, 2008, 133–143. [Google Scholar] [CrossRef]
- Storfer, A.; Alfaro, M.E.; Ridenhour, B.J.; Jancovich, J.K.; Mech, S.G.; Parris, M.J.; Collins, J.P. Phylogenetic concordance analysis shows an emerging pathogen is novel and endemic. Ecol. Lett. 2007, 10, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.L.; Schock, D.M.; Collins, J.P. Transmission dynamics of the amphibian ranavirus Ambystoma tigrinum virus. Dis. Aquat. Org. 2007, 77, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Harp, E.M.; Petranka, J.W. Ranavirus in wood frogs (Rana sylvatica): Potential sources of transmission within and between ponds. J. Wildl. Dis. 2006, 42, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Robert, J.; George, E.; De Jesús Andino, F.; Chen, G. Waterborne infectivity of the Ranavirus frog virus 3 in Xenopus laevis. Virology 2011, 417, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Greer, A.L.; Berrill, M.; Wilson, P.J. Five amphibian mortality events associated with ranavirus infection in south central Ontario, Canada. Dis. Aquat. Org. 2005, 67, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffus, A.L.J.; Pauli, B.D.; Wozney, K.; Brunetti, C.R.; Berrill, M. Frog Virus 3-Like Infections in aquatic amphibian communities. J. Wildl. Dis. 2008, 44, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.J.; Miller, D.L.; Hoverman, J.T. Ecology and pathology of amphibian ranaviruses. Dis. Aquat. Org. 2009, 87, 243–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenes, R.; Miller, D.L.; Waltzek, T.B.; Wilkes, R.P.; Tucker, J.L.; Chaney, J.C.; Hardman, R.H.; Brand, M.D.; Huether, R.R.; Gray, M.J. Susceptibility of fish and turtles to three ranaviruses isolated from different ectothermic vertebrate classes. J. Aquat. Anim. Health 2014, 26, 118–126. [Google Scholar] [CrossRef] [PubMed]
- North, A.C.; Hodgson, D.J.; Price, S.J.; Griffiths, A.G.F. Anthropogenic and ecological drivers of amphibian disease (Ranavirosis). PLoS ONE 2015, 10, e0127037. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.; Barbosa-Solomieu, V.; Chinchar, V.G. A decade of advances in iridovirus research. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2005; Volume 65, pp. 173–248. ISBN 0065-3527. [Google Scholar]
- Bollinger, T.K.; Mao, J.; Schock, D.; Brigham, R.M.; Chinchar, V.G. Pathology, isolation, and preliminary molecular characterization of a novel iridovirus from tiger salamanders in Saskatchewan. J. Wildl. Dis. 1999, 35, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Tweedell, K.; Granoff, A. viruses and renal carcinoma of Rana pipiens. Effect of frog virus 3 on developing frog embryos and larvae. J. Natl. Cancer Inst. 1968, 40, 407–410. [Google Scholar] [PubMed]
- Docherty, D.E.; Meteyer, C.U.; Wang, J.; Mao, J.; Case, S.T.; Chinchar, V.G. Diagnostic and molecular evaluation of three iridovirus-associated salamander mortality events. J. Wildl. Dis. 2003, 39, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.L.; Gray, M.J. Amphibian decline and mass mortality: The value of visualizing ranavirus in tissue sections. Vet. J. 2010, 186, 133–134. [Google Scholar] [CrossRef] [PubMed]
- Andino, F.D.J.; Jones, L.; Maggirwar, S.B.; Robert, J. Frog Virus 3 dissemination in the brain of tadpoles, but not in adult Xenopus, involves blood brain barrier dysfunction. Sci. Rep. 2016, 6, 22508. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.; Richards, K.; Collins, J. Dose and host characteristics influence virulence of ranavirus infections. Oecologia 2005, 144, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Groner, M.L.; Rollins-Smith, L.A.; Reinert, L.K.; Hempel, J.; Bier, M.E.; Relyea, R.A. Interactive effects of competition and predator cues on immune responses of leopard frogs at metamorphosis. J. Exp. Biol. 2014, 217, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Parris, M.J.; Reese, E.; Storfer, A. Antipredator behavior of chytridiomycosis-infected northern leopard frog (Rana pipiens) tadpoles. Can. J. Zool. 2006, 84, 58–65. [Google Scholar] [CrossRef]
- Rachowicz, L.J.; Briggs, C.J. Quantifying the disease transmission function: Effects of density on Batrachochytrium dendrobatidis transmission in the mountain yellow-legged frog Rana muscosa. J. Anim. Ecol. 2007, 76, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Romansic, J.M.; Johnson, P.T.; Searle, C.L.; Johnson, J.E.; Tunstall, T.S.; Han, B.A.; Rohr, J.R.; Blaustein, A.R. Individual and combined effects of multiple pathogens on Pacific treefrogs. Oecologia 2011, 166, 1029–1041. [Google Scholar] [CrossRef] [PubMed]
- Andre, S.E.; Parker, J.; Briggs, C.J. Effect of temperature on host response to Batrachochytrium dendrobatidis infection in the mountain yellow-legged frog (Rana muscosa). J. Wildl. Dis. 2008, 44, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.; Speare, R.; Hines, H.B.; Marantelli, G.; Hyatt, A.D.; McDonald, K.R.; Skerratt, L.F.; Olsen, V.; Clarke, J.; Gillespie, G.; et al. Effect of season and temperature on mortality in amphibians due to chytridiomycosis. Aust. Vet. J. 2004, 82, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodhams, D.C.; Alford, R.A.; Marantelli, G. Emerging disease of amphibians cured by elevated body temperature. Dis. Aquat. Org. 2003, 55, 65–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustamante, H.M.; Livo, L.J.; Carey, C. Effects of temperature and hydric environment on survival of the Panamanian golden frog infected with a pathogenic chytrid fungus. Integr. Zool. 2010, 5, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Garner, T.W.J.; Walker, S.; Bosch, J.; Leech, S.; Marcus Rowcliffe, J.; Cunningham, A.A.; Fisher, M.C. Life history tradeoffs influence mortality associated with the amphibian pathogen Batrachochytrium dendrobatidis. Oikos 2009, 118, 783–791. [Google Scholar] [CrossRef]
- Gervasi, S.; Gondhalekar, C.; Olson, D.H.; Blaustein, A.R. Host identity matters in the amphibian Batrachochytrium dendrobatidis system: Fine-scale patterns of variation in responses to a multi-host pathogen. PLoS ONE 2013, 8, e54490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garner, T.W.J.; Rowcliffe, J.M.; Fisher, M. Climate change, chytridiomycosis or condition: An experimental test of amphibian survival. Glob. Chang. Biol. 2011, 17, 667–675. [Google Scholar] [CrossRef]
- Márquez, M.; Nava-González, F.; Sánchez, D.; Calcagno, M.; Lampo, M. Immmunological clearance of Batrachochytrium dendrobatidis infection at a pathogen-optimal temperature in the hylid frog Hypsiboas crepitans. EcoHealth 2010, 7, 380–388. [Google Scholar] [CrossRef] [PubMed]
- McMahon, T.A.; Sears, B.F.; Venesky, M.D.; Bessler, S.M.; Brown, J.M.; Deutsch, K.; Halstead, N.T.; Lentz, G.; Tenouri, N.; Young, S.; et al. Amphibians acquire resistance to live and dead fungus overcoming fungal immunosuppression. Nature 2014, 511, 224–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, S.B. An aquatic disease on a terrestrial salamander: Individual and population level effects of the amphibian chytrid fungus, Batrachochytrium dendrobatidis, on Batrachoseps attenuatus (Plethodontidae). Copeia 2009, 2009, 653–660. [Google Scholar] [CrossRef]
- Farrer, R.A.; Weinert, L.A.; Bielby, J.; Garner, T.W.J.; Balloux, F.; Clare, F.; Bosch, J.; Cunningham, A.A.; Weldon, C.; du Preez, L.H.; et al. Multiple emergences of genetically diverse amphibian-infecting chytrids include a globalized hypervirulent recombinant lineage. Proc. Natl. Acad. Sci. USA 2011, 108, 18732–18736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahl, M.K.; Longcore, J.E.; Houlahan, J.E. Varying responses of Northeastern North American amphibians to the chytrid pathogen Batrachochytrium dendrobatidis. Conserv. Biol. 2012, 26, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Briggs, C.J.; Daszak, P. The ecology and impact of chytridiomycosis: An emerging disease of amphibians. Trends Ecol. Evol. 2010, 25, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Retallick, R.W.R.; Miera, V. Strain differences in the amphibian chytrid Batrachochytrium dendrobatidis and non-permanent, sub-lethal effects of infection. Dis. Aquat. Org. 2007, 75, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Brannelly, L.A.; Chatfield, M.W.H.; Richards-Zawacki, C.L. Field and laboratory studies of the susceptibility of the green treefrog (Hyla cinerea) to Batrachochytrium dendrobatidis Infection. PLoS ONE 2012, 7, e38473. [Google Scholar] [CrossRef] [PubMed]
- Padgett-Flohr, G.E.; Hayes, M.P. Assessment of the vulnerability of the Oregon spottedfrog (Rana pretiosa) to the amphibian chytrid fungus (Batrachochytrium dendrobatidis). Herpetol. Conserv. Biol. 2011, 6, 99–106. [Google Scholar]
- Dang, T.D.; Searle, C.L.; Blaustein, A.R. Virulence variation among strains of the emerging infectious fungus Batrachochytrium dendrobatidis (Bd) in multiple amphibian host species. Dis. Aquat. Org. 2017, 124, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Piovia-Scott, J.; Pope, K.; Joy Worth, S.; Rosenblum, E.B.; Poorten, T.; Refsnider, J.; Rollins-Smith, L.A.; Reinert, L.K.; Wells, H.L.; Rejmanek, D.; et al. Correlates of virulence in a frog-killing fungal pathogen: Evidence from a California amphibian decline. ISME J. 2015, 9, 1570–1578. [Google Scholar] [CrossRef] [PubMed]
- Doddington, B.J.; Bosch, J.; Oliver, J.A.; Grassly, N.C.; Garcia, G.; Schmidt, B.R.; Garner, T.W.J.; Fisher, M.C. Context-dependent amphibian host population response to an invading pathogen. Ecology 2013, 94, 1795–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langhammer, P.F.; Burrowes, P.A.; Lips, K.R.; Bryant, A.B.; Collins, J.P. Susceptibility to the amphibian chytrid fungus varies with ontogeny in the direct-developing frog, Eleutherodactylus coqui. J. Wildl. Dis. 2014, 50, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, E.B.; James, T.Y.; Zamudio, K.R.; Poorten, T.J.; Ilut, D.; Rodriguez, D.; Eastman, J.M.; Richards-Hrdlicka, K.; Joneson, S.; Jenkinson, T.S.; et al. Complex history of the amphibian-killing chytrid fungus revealed with genome resequencing data. Proc. Natl. Acad. Sci. USA 2013, 110, 9385–9390. [Google Scholar] [CrossRef] [PubMed]
- Voyles, J.; Johnson, L.R.; Briggs, C.J.; Cashins, S.D.; Alford, R.A.; Berger, L.; Skerratt, L.F.; Speare, R.; Rosenblum, E.B. Experimental evolution alters the rate and temporal pattern of population growth in Batrachochytrium dendrobatidis, a lethal fungal pathogen of amphibians. Ecol. Evol. 2014, 4, 3633–3641. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, S.S.; Stephens, P.R.; Hua, J.; Searle, C.L.; Xie, G.Y.; Urbina, J.; Olson, D.H.; Bancroft, B.A.; Weis, V.; Hammond, J.I.; et al. Linking ecology and epidemiology to understand predictors of multi-host responses to an emerging pathogen, the amphibian chytrid fungus. PLoS ONE 2017, 12, e0167882. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, A.R.; Romansic, J.M.; Scheessele, E.A.; Han, B.A.; Pessier, A.P.; Longcore, J.E. Interspecific variation in susceptibility of frog tadpoles to the pathogenic fungus Batrachochytrium dendrobatidis. Conserv. Biol. 2005, 19, 1460–1468. [Google Scholar] [CrossRef]
- Carey, C.; Bruzgul, J.E.; Livo, L.J.; Walling, M.L.; Kuehl, K.A.; Dixon, B.F.; Pessier, A.P.; Alford, R.A.; Rogers, K.B. Experimental exposures of boreal toads (Bufo boreas) to a pathogenic chytrid fungus (Batrachochytrium dendrobatidis). EcoHealth 2006, 3, 5–21. [Google Scholar] [CrossRef]
- Searle, C.L.; Gervasi, S.S.; Hua, J.; Hammond, J.I.; Relyea, R.A.; Olson, D.H.; Blaustein, A.R. Differential host susceptibility to Batrachochytrium dendrobatidis, an emerging amphibian pathogen. Conserv. Biol. 2011, 25, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Reeder, N.M.; Pessier, A.P.; Vredenburg, V.T. A reservoir species for the emerging amphibian pathogen Batrachochytrium dendrobatidis thrives in a landscape decimated by disease. PLoS ONE 2012, 7, e33567. [Google Scholar] [CrossRef] [PubMed]
- Gabor, C.R.; Fisher, M.C.; Bosch, J. A Non-invasive stress assay shows that tadpole populations infected with Batrachochytrium dendrobatidis have elevated corticosterone levels. PLoS ONE 2013, 8, e56054. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.D.; Steffen, J.E.; Reinert, L.K.; Cobine, P.A.; Appel, A.; Rollins-Smith, L.; Mendonça, M.T. Host stress response is important for the pathogenesis of the deadly amphibian disease, chytridiomycosis, in Litoria caerulea. PLoS ONE 2013, 8, e62146. [Google Scholar] [CrossRef] [PubMed]
- Searle, C.L.; Belden, L.K.; Du, P.; Blaustein, A.R. Stress and chytridiomycosis: Exogenous exposure to corticosterone does not alter amphibian susceptibility to a fungal pathogen. J. Exp. Zool. 2014, 321, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warne, R.W.; Crespi, E.J.; Brunner, J.L. Escape from the pond: Stress and developmental responses to ranavirus infection in wood frog tadpoles. Funct. Ecol. 2011, 25, 139–146. [Google Scholar] [CrossRef]
- Bancroft, B.A.; Han, B.A.; Searle, C.L.; Biga, L.M.; Olson, D.H.; Kats, L.B.; Lawler, J.J.; Blaustein, A.R. Species-level correlates of susceptibility to the pathogenic amphibian fungus Batrachochytrium dendrobatidis in the United States. Biodivers. Conserv. 2011, 20, 1911–1920. [Google Scholar] [CrossRef]
- Rowley, J.J.L.; Alford, R.A. Behaviour of Australian rainforest stream frogs may affect the transmission of chytridiomycosis. Dis. Aquat. Org. 2007, 77, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.A.; Bradley, P.W.; Blaustein, A.R. Ancient behaviors of larval amphibians in response to an emerging fungal pathogen, Batrachochytrium dendrobatidis. Behav. Ecol. Sociobiol. 2008, 63, 241–250. [Google Scholar] [CrossRef]
- Venesky, M.D.; Kerby, J.L.; Storfer, A.; Parris, M.J. Can differences in host behavior drive patterns of disease prevalence in tadpoles? PLoS ONE 2011, 6, e24991. [Google Scholar] [CrossRef] [PubMed]
- Lefcort, H.; Blaustein, A.R. Disease, predator avoidance, and vulnerability to predation in tadpoles. Oikos 1995, 74, 469–474. [Google Scholar] [CrossRef]
- Lefcort, H.; Eiger, S.M. Antipredatory behaviour of feverish tadpoles: Implications for pathogen transmission. Behaviour 1993, 126, 13–27. [Google Scholar] [CrossRef]
- Richards-Zawacki, C.L. Thermoregulatory behaviour affects prevalence of chytrid fungal infection in a wild population of Panamanian golden frogs. Proc. R. Soc. Lond. B Biol. Sci. 2010, 277, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlaepfer, M.; Sredl, M.; Rosen, P.; Ryan, M. High prevalence of Batrachochytrium dendrobatidis in wild populations of lowland leopard frogs Rana yavapaiensis in Arizona. EcoHealth 2007, 4, 421–427. [Google Scholar] [CrossRef]
- Han, B.A.; Kerby, J.L.; Searle, C.L.; Storfer, A.; Blaustein, A.R. Host species composition influences infection severity among amphibians in the absence of spillover transmission. Ecol. Evol. 2015, 5, 1432–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.A.; Searle, C.L.; Blaustein, A.R. Effects of an infectious fungus, Batrachochytrium dendrobatidis, on amphibian predator-prey interactions. PLoS ONE 2011, 6, e16675. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Santaliestra, M.E.; Rittenhouse, T.A.G.; Cary, T.L.; Karasov, W.H. Interspecific and postmetamorphic variation in susceptibility of three North American anurans to Batrachochytrium dendrobatidis. J. Herpetol. 2013, 47, 286–292. [Google Scholar] [CrossRef]
- Hanlon, S.M.; Lynch, K.J.; Kerby, J.; Parris, M.J. Batrachochytrium dendrobatidis exposure effects on foraging efficiencies and body size in anuran tadpoles. Dis. Aquat. Org. 2015, 112, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Parris, M.J. Hybrid response to pathogen infection in interspecific crosses between two amphibian species (Anura: Ranidae). Evol. Ecol. Res. 2004, 6, 457–471. [Google Scholar]
- Venesky, M.D.; Parris, M.J.; Storfer, A. Impacts of Batrachochytrium dendrobatidis infection on tadpole foraging performance. EcoHealth 2009, 6, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Venesky, M.D.; Wassersug, R.J.; Parris, M.J. Fungal pathogen changes the feeding kinematics of larval anurans. J. Parasitol. 2010, 96, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Hess, A.; McAllister, C.; DeMarchi, J.; Zidek, M.; Murone, J.; Venesky, M.D. Salamanders increase their feeding activity when infected with the pathogenic chytrid fungus Batrachochytrium dendrobatidis. Dis. Aquat. Org. 2015, 116, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Kuris, A.M.; Blaustein, A.R.; Alio, J.J. Hosts as islands. Am. Nat. 1980, 116, 570–586. [Google Scholar] [CrossRef]
- Gabor, C.R.; Fisher, M.C.; Bosch, J. Elevated corticosterone levels and changes in amphibian behavior are associated with Batrachochytrium dendrobatidis (Bd) infection and Bd lineage. PLoS ONE 2015, 10, e0122685. [Google Scholar] [CrossRef] [PubMed]
- Pearman, P.B.; Garner, T.W.J.; Straub, M.; Greber, U.F. Response of the Italian agile frog (Rana Latastei) to a Ranavirus, Frog Virus 3: A Model for Viral Emergence in Naïve Populations. J. Wildl. Dis. 2004, 40, 660–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, A.A.; Hyatt, A.D.; Russell, P.; Bennett, P. Emerging epidemic diseases of frogs in Britain are dependent on the source of ranavirus agent and the route of exposure. Epidemiol. Infect. 2007, 135, 1200–1212. [Google Scholar] [CrossRef] [PubMed]
- Hoverman, J.T.; Gray, M.J.; Miller, D.L. Anuran susceptibilities to ranaviruses: Role of species identity, exposure route, and a novel virus isolate. Dis. Aquat. Org. 2010, 89, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Jancovich, J.K.; Davidson, E.W.; Morado, J.F.; Jacobs, B.L.; Collins, J.P. Isolation of a lethal virus from the endangered tiger salamander Ambystoma tigrinum stebbinsi. Dis. Aquat. Org. 1997, 31, 161–167. [Google Scholar] [CrossRef]
- Jancovich, J.K.; Davidson, E.W.; Seiler, A.; Jacobs, B.L.; Collins, J.P. Transmission of the Ambystoma tigrinum virus to alternative hosts. Dis. Aquat. Org. 2001, 46, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Echaubard, P.; Leduc, J.; Pauli, B.; Chinchar, V.G.; Robert, J.; Lesbarrères, D. Environmental dependency of amphibian–ranavirus genotypic interactions: Evolutionary perspectives on infectious diseases. Evol. Appl. 2014, 7, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Rojas, S.; Richards, K.; Jancovich, J.K.; Davidson, E.W. Davidson Influence of temperature on Ranavirus infection in larval salamanders Ambystoma tigrinum. Dis. Aquat. Org. 2005, 63, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Gray, M.; Storfer, A. Ecopathology of ranaviruses Infecting amphibians. Viruses 2011, 3, 2351–2373. [Google Scholar] [CrossRef] [PubMed]
- Hoverman, J.T.; Gray, M.J.; Haislip, N.A.; Miller, D.L. Phylogeny, life history, and ecology contribute to differences in amphibian susceptibility to ranaviruses. EcoHealth 2011, 8, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Schock, D.M.; Bollinger, T.K.; Collins, J.P. Mortality rates differ among amphibian populations exposed to three strains of a lethal ranavirus. EcoHealth 2009, 6, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Andino, F.D.J.; Chen, G.; Li, Z.; Grayfer, L.; Robert, J. Susceptibility of Xenopus laevis tadpoles to infection by the ranavirus Frog-Virus 3 correlates with a reduced and delayed innate immune response in comparison with adult frogs. Virology 2012, 432, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Brenes, R.; Gray, M.J.; Waltzek, T.B.; Wilkes, R.P.; Miller, D.L. Transmission of ranavirus between ectothermic vertebrate hosts. PLoS ONE 2014, 9, e92476. [Google Scholar] [CrossRef] [PubMed]
- Haislip, N.A.; Gray, M.J.; Hoverman, J.T.; Miller, D.L. Development and disease: How susceptibility to an emerging pathogen changes through anuran development. PLoS ONE 2011, 6, e22307. [Google Scholar] [CrossRef] [PubMed]
- Rollins-Smith, L.A. Metamorphosis and the amphibian immune system. Immunol. Rev. 1998, 166, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Maniero, G.D.; Morales, H.; Gantress, J.; Robert, J. Generation of a long-lasting, protective, and neutralizing antibody response to the ranavirus FV3 by the frog Xenopus. Dev. Comp. Immunol. 2006, 30, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Kiesecker, J.M.; Blaustein, A.R.; Belden, L.K. Complex causes of amphibian population declines. Nature 2001, 410, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Rollins-Smith, L.A.; Ramsey, J.P.; Pask, J.D.; Reinert, L.K.; Woodhams, D.C. Amphibian immune defenses against chytridiomycosis: Impacts of changing environments. Integr. Comp. Biol. 2011, 51, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, A.R.; Wake, D.B.; Sousa, W.P. Amphibian declines: Judging stability, persistence, and susceptibility of populations to local and global extinctions. Conserv. Biol. 1994, 8, 60–71. [Google Scholar] [CrossRef]
- Williamson, C.E.; Madronich, S.; Lal, A.; Zepp, R.G.; Lucas, R.M.; Overholt, E.P.; Rose, K.C.; Schladow, S.G.; Lee-Taylor, J. Climate change-induced increases in precipitation are reducing the potential for solar ultraviolet radiation to inactivate pathogens in surface waters. Sci. Rep. 2017, 7, 13033. [Google Scholar] [CrossRef] [PubMed]
- Overholt, E.P.; Hall, S.R.; Williamson, C.E.; Meikle, C.K.; Duffy, M.A.; Cáceres, C.E. Solar radiation decreases parasitism in Daphnia. Ecol. Lett. 2012, 15, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Garcia, T.S.; Romansic, J.M.; Blaustein, A.R. Survival of three species of anuran metamorphs exposed to UV-B radiation and the pathogenic fungus Batrachochytrium dendrobatidis. Dis. Aquat. Org. 2006, 72, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Searle, C.; Belden, L.; Bancroft, B.; Han, B.; Biga, L.; Blaustein, A. Experimental examination of the effects of ultraviolet-B radiation in combination with other stressors on frog larvae. Oecologia 2010, 162, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Santaliestra, M.E.; Fisher, M.C.; Fernández-Beaskoetxea, S.; Fernández-Benéitez, M.J.; Bosch, J. Ambient ultraviolet b radiation and prevalence of infection by Batrachochytrium dendrobatidis in two amphibian species. Conserv. Biol. 2011, 25, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Sanchez-Tomé, E.; Fernández-Loras, A.; Oliver, J.A.; Fisher, M.C.; Garner, T.W.J. Successful elimination of a lethal wildlife infectious disease in nature. Biol. Lett. 2015, 11. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Carrascal, L.M.; Duran, L.; Walker, S.; Fisher, M.C. Climate change and outbreaks of amphibian chytridiomycosis in a montane area of Central Spain; is there a link? Proc. R. Soc. Lond. B Biol. Sci. 2007, 274, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, P.T.; Richardson, J.M.L.; Govindarajulu, P.; Anholt, B.R. Higher temperature variability increases the impact of Batrachochytrium dendrobatidis and shifts interspecific interactions in tadpole mesocosms. Ecol. Evol. 2012, 2, 2450–2459. [Google Scholar] [CrossRef] [PubMed]
- Raffel, T.R.; Halstead, N.T.; McMahon, T.A.; Davis, A.K.; Rohr, J.R. Temperature variability and moisture synergistically interact to exacerbate an epizootic disease. Proc. Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Rumschlag, S.L.; Boone, M.D.; Fellers, G. The effects of the amphibian chytrid fungus, insecticide exposure, and temperature on larval anuran development and survival. Environ. Toxicol. Chem. 2014, 33, 2545–2550. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.J.; St-Hilaire, S.; Corn, P.S. Temperature, hydric environment, and prior pathogen exposure alter the experimental severity of chytridiomycosis in boreal toads. Dis. Aquat. Org. 2011, 95, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, G.Y.; Olson, D.H.; Blaustein, A.R. Projecting the Global Distribution of the emerging amphibian fungal pathogen, Batrachochytrium dendrobatidis, based on IPCC climate futures. PLoS ONE 2016, 11, e0160746. [Google Scholar] [CrossRef] [PubMed]
- Relyea, R.A.; Edwards, K. What doesn’t kill you makes you sluggish: How sublethal pesticides alter predator–prey interactions. Copeia 2010, 2010, 558–567. [Google Scholar] [CrossRef]
- Relyea, R.A.; Jones, D.K. The toxicity of Roundup Original Max® to 13 species of larval amphibians. Environ. Toxicol. Chem. 2009, 28, 2004–2008. [Google Scholar] [CrossRef] [PubMed]
- Davidson, C. Declining Downwind: Amphibian population declines in California and historical pesticide use. Ecol. Appl. 2004, 14, 1892–1902. [Google Scholar] [CrossRef]
- Hayes, T.B.; Case, P.; Chui, S.; Chung, D.; Haeffele, C.; Haston, K.; Lee, M.; Mai, V.P.; Marjuoa, Y.; Parker, J.; et al. Pesticide mixtures, endocrine disruption, and amphibian declines: Are we underestimating the impact? Environ. Health Perspect. 2006, 114, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Relyea, R.A.; Diecks, N. An unforeseen chain of events: Lethal effects of pesticides on frogs at sublethal concentrations. Ecol. Appl. 2008, 18, 1728–1742. [Google Scholar] [CrossRef] [PubMed]
- Christin, M.-S.; Gendron, A.D.; Brousseau, P.; Ménard, L.; Marcogliese, D.J.; Cyr, D.; Ruby, S.; Fournier, M. Effects of agricultural pesticides on the immune system of Rana pipiens and on its resistance to parasitic infection. Environ. Toxicol. Chem. 2003, 22, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Gilbertson, M.-K.; Haffner, G.D.; Drouillard, K.G.; Albert, A.; Dixon, B. Immunosuppression in the northern leopard frog (Rana pipiens) induced by pesticide exposure. Environ. Toxicol. Chem. 2003, 22, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.K.; Williams, E.S.; Mills, K.W. Effects of malathion on disease susceptibility in woodhouse’s toads. J. Wildl. Dis. 1999, 35, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Miiller, T.; Kerby, J.L. The interactive effect of an emerging infectious disease and an emerging contaminant on Woodhouse’s toad (Anaxyrus woodhousii) tadpoles. Environ. Toxicol. Chem. 2013, 32, 2003–2008. [Google Scholar] [CrossRef] [PubMed]
- Buck, J.C.; Hua, J.; Brogan, W.R., III; Dang, T.D.; Urbina, J.; Bendis, R.J.; Stoler, A.B.; Blaustein, A.R.; Relyea, R.A. Effects of pesticide mixtures on host-pathogen dynamics of the amphibian chytrid fungus. PLoS ONE 2015, 10, e0132832. [Google Scholar] [CrossRef] [PubMed]
- Buck, J.C.; Scheessele, E.A.; Relyea, R.A.; Blaustein, A.R. The effects of multiple stressors on wetland communities: Pesticides, pathogens and competing amphibians. Freshw. Biol. 2012, 57, 61–73. [Google Scholar] [CrossRef]
- Davidson, C.; Benard, M.F.; Shaffer, H.B.; Parker, J.M.; O’Leary, C.; Conlon, J.M.; Rollins-Smith, L.A. Effects of chytrid and carbaryl exposure on survival, growth and skin peptide defenses in foothill yellow-legged frogs. Environ. Sci. Technol. 2007, 41, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Edge, C.B.; Gahl, M.K.; Thompson, D.G.; Houlahan, J.E. Laboratory and field exposure of two species of juvenile amphibians to a glyphosate-based herbicide and Batrachochytrium dendrobatidis. Sci. Total Environ. 2013, 444, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Hanlon, S.M.; Parris, M.J. The interactive effects of chytrid fungus, pesticides, and exposure timing on gray treefrog (Hyla versicolor) larvae. Environ. Toxicol. Chem. 2014, 33, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.K.; Dang, T.D.; Urbina, J.; Bendis, R.J.; Buck, J.C.; Cothran, R.D.; Blaustein, A.R.; Relyea, R.A. Effect of simultaneous amphibian exposure to pesticides and an emerging fungal pathogen, Batrachochytrium dendrobatidis. Environ. Sci. Technol. 2017, 51, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Kleinhenz, P.; Boone, M.D.; Fellers, G. Effects of the amphibian chytrid fungus and four insecticides on pacific treefrogs (Pseudacris regilla). J. Herpetol. 2012, 46, 625–631. [Google Scholar] [CrossRef]
- McMahon, T.A.; Brannelly, L.A.; Chatfield, M.W.H.; Johnson, P.T.J.; Joseph, M.B.; McKenzie, V.J.; Richards-Zawacki, C.L.; Venesky, M.D.; Rohr, J.R. Chytrid fungus Batrachochytrium dendrobatidis has nonamphibian hosts and releases chemicals that cause pathology in the absence of infection. Proc. Natl. Acad. Sci. USA 2013, 110, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Paetow, L.J.; Daniel McLaughlin, J.; Cue, R.I.; Pauli, B.D.; Marcogliese, D.J. Effects of herbicides and the chytrid fungus Batrachochytrium dendrobatidis on the health of post-metamorphic northern leopard frogs (Lithobates pipiens). Ecotoxicol. Environ. Saf. 2012, 80, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.S.; Rumschlag, S.L.; Boone, M.D. Effects of amphibian chytrid fungus exposure on American toads in the presence of an insecticide. Environ. Toxicol. Chem. 2014, 33, 2541–2544. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; Raffel, T.R.; Halstead, N.T.; McMahon, T.A.; Johnson, S.A.; Boughton, R.K.; Martin, L.B. Early-life exposure to a herbicide has enduring effects on pathogen-induced mortality. Proc. R. Soc. Lond. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef]
- Forson, D.; Storfer, A. Effects of atrazine and iridovirus infection on survival and life-history traits of the long-toed salamander (Ambystoma macrodactylum). Environ. Toxicol. Chem. 2006, 25, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Kerby, J.L.; Storfer, A. Combined effects of atrazine and chlorpyrifos on susceptibility of the tiger salamander to Ambystoma tigrinum Virus. EcoHealth 2009, 6, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Stehle, S.; Schulz, R. Agricultural insecticides threaten surface waters at the global scale. Proc. Natl. Acad. Sci. USA 2015, 112, 5750–5755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Civitello, D.J.; Cohen, J.; Fatima, H.; Halstead, N.T.; Liriano, J.; McMahon, T.A.; Ortega, C.N.; Sauer, E.L.; Sehgal, T.; Young, S.; et al. Biodiversity inhibits parasites: Broad evidence for the dilution effect. Proc. Natl. Acad. Sci. USA 2015, 112, 8667–8671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafferty, K.D. Biodiversity loss decreases parasite diversity: Theory and patterns. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 2814–2827. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Keesing, F. Biodiversity and Disease Risk: The Case of Lyme Disease. Conserv. Biol. 2000, 14, 722–728. [Google Scholar] [CrossRef]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 7039–7044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, P.T.J.; Preston, D.L.; Hoverman, J.T.; Richgels, K.L.D. Biodiversity decreases disease through predictable changes in host community competence. Nature 2013, 494, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Venesky, M.D.; Liu, X.; Sauer, E.L.; Rohr, J.R. Linking manipulative experiments to field data to test the dilution effect. J. Anim. Ecol. 2014, 83, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Venesky, M.D.; Hanlon, S.M.; Lynch, K.; Parris, M.J.; Rohr, J.R. Optimal digestion theory does not predict the effect of pathogens on intestinal plasticity. Biol. Lett. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Buck, J.C.; Truong, L.; Blaustein, A.R. Predation by zooplankton on Batrachochytrium dendrobatidis: Biological control of the deadly amphibian chytrid fungus? Biodivers. Conserv. 2011, 20, 3549–3553. [Google Scholar] [CrossRef]
- Searle, C.L.; Mendelson, J.R.; Green, L.E.; Duffy, M.A. Daphnia predation on the amphibian chytrid fungus and its impacts on disease risk in tadpoles. Ecol. Evol. 2013, 3, 4129–4138. [Google Scholar] [CrossRef] [PubMed]
- Schmeller, D.S.; Blooi, M.; Martel, A.; Garner, T.W.J.; Fisher, M.C.; Azemar, F.; Clare, F.C.; Leclerc, C.; Jäger, L.; Guevara-Nieto, M.; et al. Microscopic aquatic predators strongly affect infection dynamics of a globally emerged pathogen. Curr. Biol. 2014, 24, 176–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, M.A.; Hall, S.R.; Tessier, A.J.; Huebner, M. Selective predators and their parasitized prey: Are epidemics in zooplankton under top-down control? Limnol. Oceanogr. 2005, 50, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Lafferty, K.D. Fishing for lobsters indirectly increases epidemics in sea urchins. Ecol. Appl. 2004, 14, 1566–1573. [Google Scholar] [CrossRef]
- Packer, C.; Holt, R.D.; Hudson, P.J.; Lafferty, K.D.; Dobson, A.P. Keeping the herds healthy and alert: Implications of predator control for infectious disease. Ecol. Lett. 2003, 6, 797–802. [Google Scholar] [CrossRef]
- Groner, M.L.; Relyea, R.A. Predators reduce Batrachochytrium dendrobatidis infection loads in their prey. Freshw. Biol. 2015, 60, 1699–1704. [Google Scholar] [CrossRef]
- Groner, M.L.; Buck, J.C.; Gervasi, S.; Blaustein, A.R.; Reinert, L.K.; Rollins-Smith, L.A.; Bier, M.E.; Hempel, J.; Relyea, R.A. Larval exposure to predator cues alters immune function and response to a fungal pathogen in post-metamorphic wood frogs. Ecol. Appl. 2013, 23, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Kerby, J.L.; Hart, A.J.; Storfer, A. Combined Effects of Virus, Pesticide, and Predator Cue on the Larval Tiger Salamander (Ambystoma tigrinum). EcoHealth 2011, 8, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Haislip, N.A.; Hoverman, J.T.; Miller, D.L.; Gray, M.J. Natural stressors and disease risk: Does the threat of predation increase amphibian susceptibility to ranavirus? Can. J. Zool. 2012, 90, 893–902. [Google Scholar] [CrossRef]
- Wilbur, H.M. Density-dependent aspects of growth and metamorphosis in Bufo americanus. Ecology 1977, 58, 196–200. [Google Scholar] [CrossRef]
- Parris, M.J.; Cornelius, T.O. Fungal pathogen causes competitive and developmental stress in larval amphibian communities. Ecology 2004, 85, 3385–3395. [Google Scholar] [CrossRef]
- Bielby, J.; Fisher, M.C.; Clare, F.C.; Rosa, G.M.; Garner, T.W.J. Host species vary in infection probability, sub-lethal effects, and costs of immune response when exposed to an amphibian parasite. Sci. Rep. 2015, 5, 10828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeve, B.C.; Crespi, E.J.; Whipps, C.M.; Brunner, J.L. Natural stressors and ranavirus susceptibility in larval wood frogs (Rana sylvatica). EcoHealth 2013, 10, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.; Beaty, L.; Guitard, A.; Russell, D. Heterogeneities in the infection process drive ranavirus transmission. Ecology 2017, 98, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Ezenwa, V.O.; Jolles, A.E. From Host immunity to pathogen invasion: The effects of helminth coinfection on the dynamics of microparasites. Integr. Comp. Biol. 2011, 51, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Kik, M.; Stege, M.; Boonyarittichaikij, R.; van Asten, A. Concurrent ranavirus and Batrachochytrium dendrobatidis infection in captive frogs (Phyllobates and Dendrobates species), The Netherlands, 2012: A first report. Vet. J. 2012, 194, 247–249. [Google Scholar] [CrossRef] [PubMed]
- Warne, R.W.; LaBumbard, B.; LaGrange, S.; Vredenburg, V.T.; Catenazzi, A. Co-Infection by chytrid fungus and ranaviruses in wild and harvested frogs in the tropical andes. PLoS ONE 2016, 11, e0145864. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, S.M.; Geerdes, E.; Chacon, I.; Ballestero, R.E.; Jimenez, R.R.; Donnelly, M.A.; Kerby, J.L. Infection and co-infection by the amphibian chytrid fungus and ranavirus in wild Costa Rican frogs. Dis. Aquat. Org. 2013, 104, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuerthner, V.P.; Hua, J.; Hoverman, J.T. The benefits of coinfection: Trematodes alter disease outcomes associated with virus infection. J. Anim. Ecol. 2017, 86, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Kuris, A.; Blaustein, A. Ectoparasitic mites on rodents: Application of the island biogeography theory. Science 1977, 195, 596–598. [Google Scholar] [CrossRef] [PubMed]
- ASA Amphibian Red List Authority. Amphibian Survival Alliance. Available online: http://www.amphibians.org/redlist (accessed on 13 November 2017).
- Bradley, P.W.; Gervasi, S.S.; Hua, J.; Cothran, R.D.; Relyea, R.A.; Olson, D.H.; Blaustein, A.R. Differences in sensitivity to the fungal pathogen Batrachochytrium dendrobatidis among amphibian populations. Conserv. Biol. 2015, 29, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Pearman, P.B.; Garner, T.W.J. Susceptibility of Italian agile frog populations to an emerging strain of Ranavirus parallels population genetic diversity. Ecol. Lett. 2005, 8, 401–408. [Google Scholar] [CrossRef]
- Rachowicz, L.J.; Vredenburg, V.T. Transmission of Batrachochytrium dendrobatidis within and between amphibian life stages. Dis. Aquat. Org. 2004, 61, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Hairston, N.G. Ecological Experiments: Purpose, Design and Execution; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Underwood, A.J. Experiments in Ecology: Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Hoverman, J.; Mihaljevic, J.; Richgels, K.; Kerby, J.; Johnson, P. Widespread co-occurrence of virulent pathogens within california amphibian communities. EcoHealth 2012, 9, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; Sutherland, D.R. Amphibian deformities and Ribeiroia infection: An emerging helminthiasis. Trends Parasitol. 2003, 19, 332–335. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Wake, D.B. Declining amphibian populations—A global phenomenon. Trends Ecol. Evol. 1990, 5, 203–204. [Google Scholar] [CrossRef]
- Jenkins, S.H. How Science Works: Evaluating Evidence in Biology and Medicine; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of species diversity on disease risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, A.R.; Tunstall, T.; DiRenzo, G.V.; Hughey, M.C.; Rebollar, E.A.; Belden, L.K.; Harris, R.N.; Ibáñez, R.; Lips, K.R.; Zamudio, K.R. more than skin deep: Functional genomic basis for resistance to amphibian chytridiomycosis. Genome Biol. Evol. 2015, 7, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Tobler, U.; Schmidt, B.R. Within- and among-population variation in chytridiomycosis-induced mortality in the toad Alytes obstetricans. PLoS ONE 2010, 5, e10927. [Google Scholar] [CrossRef]
- Geiger, C.C.; Bregnard, C.; Maluenda, E.; Voordouw, M.J.; Schmidt, B.R. Antifungal treatment of wild amphibian populations caused a transient reduction in the prevalence of the fungal pathogen, Batrachochytrium dendrobatidis. Sci. Rep. 2017, 7, 5956. [Google Scholar] [CrossRef] [PubMed]
- Padgett-Flohr, G.E. Pathogenicity of Batrachochytrium dendrobatidis in two threatened California amphibians: Rana draytonii and Ambystoma californiense. Herpetol. Conserv. Biol. 2008, 3, 182–191. [Google Scholar]
- Davidson, E.W.; Parris, M.; Collins, J.P.; Longcore, J.E.; Pessier, A.P.; Brunner, J.; Beaupre, S.J. Pathogenicity and transmission of chytridiomycosis in tiger salamanders (Ambystoma tigrinum). Copeia 2003, 2003, 601–607. [Google Scholar] [CrossRef]
- Peterson, A.C.; McKenzie, V.J. Investigating differences across host species and scales to explain the distribution of the amphibian pathogen Batrachochytrium dendrobatidis. PLoS ONE 2014, 9, e107441. [Google Scholar] [CrossRef] [PubMed]
- Antwis, R.; Weldon, C. Amphibian skin defences show variation in ability to inhibit growth of Batrachochytrium dendrobatidis from the global panzootic lineage. Microbiology 2017, 163, 1835–1838. [Google Scholar] [CrossRef] [PubMed]
- Karavlan, S.A.; Venesky, M.D. Thermoregulatory behavior of Anaxyrus americanus in response to infection with Batrachochytrium dendrobatidis. Copeia 2016, 104, 746–751. [Google Scholar] [CrossRef]
- Poorten, T.J.; Rosenblum, E.B. Comparative study of host response to chytridiomycosis in a susceptible and a resistant toad species. Mol. Ecol. 2016, 25, 5663–5679. [Google Scholar] [CrossRef] [PubMed]
- Barnhart, K.; Forman, M.E.; Umile, T.P.; Kueneman, J.; McKenzie, V.; Salinas, I.; Minbiole, K.P.C.; Woodhams, D.C. Identification of Bufadienolides from the boreal toad, Anaxyrus boreas, active against a fungal pathogen. Microb. Ecol. 2017, 74, 990–1000. [Google Scholar] [CrossRef] [PubMed]
- Marcum, R.; St-Hilaire, S.; Murphy, P.; Rodnick, K. Effects of Batrachochytrium dendrobatidis infection on ion concentrations in the boreal toad Anaxyrus (Bufo) boreas boreas. Dis. Aquat. Org. 2010, 91, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Voyles, J.; Woodhams, D.C.; Saenz, V.; Byrne, A.Q.; Perez, R.; Rios-Sotelo, G.; Ryan, M.J.; Bletz, M.C.; Sobell, F.A.; McLetchie, S.; et al. Shifts in disease dynamics in a tropical amphibian assemblage are not due to pathogen attenuation. Science 2018, 359, 1517–1519. [Google Scholar] [CrossRef] [PubMed]
- DiRenzo, G.V.; Langhammer, P.F.; Zamudio, K.R.; Lips, K.R. Fungal infection intensity and zoospore output of Atelopus zeteki, a potential acute chytrid supershedder. PLoS ONE 2014, 9, e93356. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A.R.; Savage, A.E.; DiRenzo, G.V.; Langhammer, P.; Lips, K.R.; Zamudio, K.R. Fighting a Losing Battle: Vigorous immune response countered by pathogen suppression of host defenses in the chytridiomycosis-susceptible frog Atelopus zeteki. G3: Genes|Genomes|Genet. 2014, 4, 1275–1289. [Google Scholar] [CrossRef] [PubMed]
- Becker, M.H.; Harris, R.N.; Minbiole, K.P.C.; Schwantes, C.R.; Rollins-Smith, L.A.; Reinert, L.K.; Brucker, R.M.; Domangue, R.J.; Gratwicke, B. Towards a better understanding of the use of probiotics for preventing chytridiomycosis in panamanian golden frogs. EcoHealth 2011, 8, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Villarroel, L.; Garcia, F.; Nava-Gonzalez, F.; Lampo, M. Susceptibility of the endangered frog Dendropsophus meridensis to the pathogenic fungus Batrachochytrium dendrobatidis. Dis. Aquat. Org. 2013, 107, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, V.M.; Rothermel, B.B.; Pessier, A.P. Experimental infection of North American plethodontid salamanders with the fungus Batrachochytrium dendrobatidis. Dis. Aquat. Org. 2009, 84, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chinnadurai, S.K.; Cooper, D.; Dombrowski, D.S.; Poore, M.F.; Levy, M.G. Experimental infection of native north carolina salamanders with Batrachochytrium dendrobatidis. J. Wildl. Dis. 2009, 45, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Parris, M.J.; Baud, D.R.; Quattro, J.M. Interactive effects of a heavy metal and chytridiomycosis on gray treefrog larvae (Hyla chrysoscelis). Copeia 2004, 2004, 344–350. [Google Scholar] [CrossRef]
- Gaietto, K.M.; Rumschlag, S.L.; Boone, M.D. Effects of pesticide exposure and the amphibian chytrid fungus on gray treefrog (Hyla chrysoscelis) metamorphosis. Environ. Toxicol. Chem. 2014, 33, 2358–2362. [Google Scholar] [CrossRef] [PubMed]
- Miaud, C.; Pozet, F.; Gaudin, N.C.G.; Martel, A.; Pasmans, F.; Labrut, S. Ranavirus causes mass die-offs of alpine amphibians in the Southwestern Alps, France. J. Wildl. Dis. 2016, 52, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Ohmer, M.E.B.; Cramp, R.L.; Russo, C.J.M.; White, C.R.; Franklin, C.E. Skin sloughing in susceptible and resistant amphibians regulates infection with a fungal pathogen. Sci. Rep. 2017, 7, 3529. [Google Scholar] [CrossRef] [PubMed]
- Shaw, S.D.; Bishop, P.J.; Berger, L.; Skerratt, L.F.; Garland, S.; Gleeson, D.M.; Haigh, A.; Herbert, S.; Speare, R. Experimental infection of self-cured Leiopelma archeyi with the amphibian chytrid Batrachochytrium dendrobatidis. Dis. Aquat. Org. 2010, 92, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, M.P.; Clulow, J.; Mahony, M.J. Host species determines whether infection load increases beyond disease-causing thresholds following exposure to the amphibian chytrid fungus. Anim. Conserv. 2010, 13, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Woodhams, D.C.; Ardipradja, K.; Alford, R.A.; Marantelli, G.; Reinert, L.K.; Rollins-Smith, L.A. Resistance to chytridiomycosis varies among amphibian species and is correlated with skin peptide defenses. Anim. Conserv. 2007, 10, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Cheatsazan, H.; de Almedia, A.P.L. G.; Russell, A.F.; Bonneaud, C. Experimental evidence for a cost of resistance to the fungal pathogen, Batrachochytrium dendrobatidis, for the palmate newt, Lissotriton helveticus. BMC Ecol. 2013, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Walke, J.B.; Becker, M.H.; Loftus, S.C.; House, L.L.; Teotonio, T.L.; Minbiole, K.P.C.; Belden, L.K. Community structure and function of amphibian skin microbes: An experiment with bullfrogs exposed to a chytrid fungus. PLoS ONE 2015, 10, e0139848. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, M.W.H.; Brannelly, L.A.; Robak, M.J.; Freeborn, L.; Lailvaux, S.P.; Richards-Zawacki, C.L. Fitness consequences of infection by Batrachochytrium dendrobatidis in northern leopard frogs (Lithobates pipiens). EcoHealth 2013, 10, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Venesky, M.D.; Wilcoxen, T.E.; Rensel, M.A.; Rollins-Smith, L.; Kerby, J.L.; Parris, M.J. Dietary protein restriction impairs growth, immunity, and disease resistance in southern leopard frog tadpoles. Oecologia 2012, 169, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Holden, W.M.; Reinert, L.K.; Hanlon, S.M.; Parris, M.J.; Rollins-Smith, L.A. Development of antimicrobial peptide defenses of southern leopard frogs, Rana sphenocephala, against the pathogenic chytrid fungus, Batrachochytrium dendrobatidis. Dev. Comp. Immunol. 2015, 48, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Savage, A.E.; Zamudio, K.R. MHC genotypes associate with resistance to a frog-killing fungus. Proc. Natl. Acad. Sci. USA 2011, 108, 16705–16710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cashins, S.D.; Grogan, L.F.; McFadden, M.; Hunter, D.; Harlow, P.S.; Berger, L.; Skerratt, L.F. Prior infection does not improve survival against the amphibian disease chytridiomycosis. PLoS ONE 2013, 8, e56747. [Google Scholar] [CrossRef] [PubMed]
- Ohmer, M.E.B.; Cramp, R.L.; White, C.R.; Franklin, C.E. Skin sloughing rate increases with chytrid fungus infection load in a susceptible amphibian. Funct. Ecol. 2015, 29, 674–682. [Google Scholar] [CrossRef]
- Young, S.; Whitehorn, P.; Berger, L.; Skerratt, L.F.; Speare, R.; Garland, S.; Webb, R. Defects in host immune function in tree frogs with chronic chytridiomycosis. PLoS ONE 2014, 9, e107284. [Google Scholar] [CrossRef] [PubMed]
- Greenspan, S.E.; Bower, D.S.; Webb, R.J.; Berger, L.; Rudd, D.; Schwarzkopf, L.; Alford, R.A. White blood cell profiles in amphibians help to explain disease susceptibility following temperature shifts. Dev. Comp. Immunol. 2017, 77, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Carver, S.; Bell, B.D.; Waldman, B. Does chytridiomycosis disrupt amphibian skin function? Copeia 2010, 2010, 487–495. [Google Scholar] [CrossRef]
- Bataille, A.; Cashins, S.D.; Grogan, L.; Skerratt, L.F.; Hunter, D.; McFadden, M.; Scheele, B.; Brannelly, L.A.; Macris, A.; Harlow, P.S.; et al. Susceptibility of amphibians to chytridiomycosis is associated with MHC class II conformation. Proc. Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brannelly, L.A.; Webb, R.; Skerratt, L.F.; Berger, L. Amphibians with infectious disease increase their reproductive effort: Evidence for the terminal investment hypothesis. Open Biol. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- McMahon, T.A.; Rohr, J.R. Transition of chytrid fungus infection from mouthparts to hind limbs during amphibian metamorphosis. EcoHealth 2015, 12, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Woodhams, D.C.; Bigler, L.; Marschang, R. Tolerance of fungal infection in European water frogs exposed to Batrachochytrium dendrobatidis after experimental reduction of innate immune defenses. BMC Vet. Res. 2012, 8, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonner, C.W.; Patel, S.A.; Boord, S.M.; Venesky, M.D.; Woodley, S.K. Effects of corticosterone on infection and disease in salamanders exposed to the amphibian fungal pathogen Batrachochytrium dendrobatidis. Dis. Aquat. Org. 2017, 123, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, S.S.; Hunt, E.G.; Lowry, M.; Blaustein, A.R. Temporal patterns in immunity, infection load and disease susceptibility: Understanding the drivers of host responses in the amphibian-chytrid fungus system. Funct. Ecol. 2014, 28, 569–578. [Google Scholar] [CrossRef]
- Rosenblum, E.B.; Poorten, T.J.; Settles, M.; Murdoch, G.K. Only skin deep: Shared genetic response to the deadly chytrid fungus in susceptible frog species. Mol. Ecol. 2012, 21, 3110–3120. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, J.R.; Waddle, A.W.; Rivera, R.; Harrison, D.T.; Ellison, S.; Forrest, M.J.; Vredenburg, V.T.; van Breukelen, F. Batrachochytrium dendrobatidis and the decline and survival of the relict leopard frog. EcoHealth 2017, 14, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Pask, J.D.; Cary, T.L.; Rollins-Smith, L.A. Skin peptides protect juvenile leopard frogs (Rana pipiens) against chytridiomycosis. J. Exp. Biol. 2013, 216, 2908. [Google Scholar] [CrossRef] [PubMed]
- Jani, A.J.; Briggs, C.J. The pathogen Batrachochytrium dendrobatidis disturbs the frog skin microbiome during a natural epidemic and experimental infection. Proc. Natl. Acad. Sci. USA 2014, 111, E5049. [Google Scholar] [CrossRef] [PubMed]
- Price, S.J.; Garner, T.W.J.; Balloux, F.; Ruis, C.; Paszkiewicz, K.H.; Moore, K.; Griffiths, A.G.F. A de novo assembly of the common frog (Rana temporaria) transcriptome and comparison of transcription following exposure to ranavirus and Batrachochytrium dendrobatidis. PLoS ONE 2015, 10, e0130500. [Google Scholar] [CrossRef] [PubMed]
- Ribas, L.; Li, M.-S.; Doddington, B.J.; Robert, J.; Seidel, J.A.; Kroll, J.S.; Zimmerman, L.B.; Grassly, N.C.; Garner, T.W.; Fisher, M.C. Expression profiling the temperature-dependent amphibian response to infection by Batrachochytrium dendrobatidis. PLoS ONE 2009, 4, e8408. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, E.B.; Poorten, T.J.; Settles, M.; Murdoch, G.K.; Robert, J.; Maddox, N.; Eisen, M.B. Genome-wide transcriptional response of Silurana (Xenopus) tropicalis to infection with the deadly chytrid fungus. PLoS ONE 2009, 4, e6494. [Google Scholar] [CrossRef] [PubMed]
- Fites, J.S.; Ramsey, J.P.; Holden, W.M.; Collier, S.P.; Sutherland, D.M.; Reinert, L.K.; Gayek, A.S.; Dermody, T.S.; Aune, T.M.; Oswald-Richter, K.; et al. The invasive chytrid fungus of amphibians paralyzes lymphocyte responses. Science 2013, 342, 366. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, J.P.; Reinert, L.K.; Harper, L.K.; Woodhams, D.C.; Rollins-Smith, L.A. Immune defenses against Batrachochytrium dendrobatidis, a fungus linked to global amphibian declines, in the South African clawed frog, Xenopus laevis. Infect. Immun. 2010, 78, 3981–3992. [Google Scholar] [CrossRef] [PubMed]
- Fites, J.S.; Reinert, L.K.; Chappell, T.M.; Rollins-Smith, L.A. Inhibition of local immune responses by the frog-killing fungus Batrachochytrium dendrobatidis. Infect. Immun. 2014, 82, 4698–4706. [Google Scholar] [CrossRef] [PubMed]
- Stegen, G.; Pasmans, F.; Schmidt, B.R.; Rouffaer, L.O.; Van Praet, S.; Schaub, M.; Canessa, S.; Laudelout, A.; Kinet, T.; Adriaensen, C.; et al. Drivers of salamander extirpation mediated by Batrachochytrium salamandrivorans. Nature 2017, 544, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Blooi, M.; Pasmans, F.; Rouffaer, L.; Haesebrouck, F.; Vercammen, F.; Martel, A. Successful treatment of Batrachochytrium salamandrivorans infections in salamanders requires synergy between voriconazole, polymyxin E and temperature. Sci. Rep. 2015, 5, 11788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picco, A.M.; Brunner, J.L.; Collins, J.P. Susceptibility of the endangered California tiger salamander, Ambystoma californiense, to ranavirus infection. J. Wildl. Dis. 2007, 43, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Earl, J.E.; Chaney, J.C.; Sutton, W.B.; Lillard, C.E.; Kouba, A.J.; Langhorne, C.; Krebs, J.; Wilkes, R.P.; Hill, R.D.; Miller, D.L.; et al. Ranavirus could facilitate local extinction of rare amphibian species. Oecologia 2016, 182, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Duffus, A.L.J.; Nichols, R.A.; Garner, T.W.J. Experimental evidence in support of single host maintenance of a multihost pathogen. Ecosphere 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Cullen, B.R.; Owens, L. Experimental challenge and clinical cases of Bohle iridovirus (BIV) in native Australian anurans. Dis. Aquat. Org. 2002, 49, 83–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, B.R.; Owens, L.; Whittington, R.J. Experimental infection of Australian anurans (Limnodynastes terraereginae and Litoria latopalmata) with Bohle iridovirus. Dis. Aquat. Org. 1995, 23, 83–92. [Google Scholar] [CrossRef]
- Morrison, E.A.; Garner, S.; Echaubard, P.; Lesbarrères, D.; Kyle, C.J.; Brunetti, C.R. Complete genome analysis of a frog virus 3 (FV3) isolate and sequence comparison with isolates of differing levels of virulence. Virol. J. 2014, 11, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, W.B.; Gray, M.J.; Hardman, R.H.; Wilkes, R.P.; Kouba, A.J.; Miller, D.L. High susceptibility of the endangered dusky gopher frog to ranavirus. Dis. Aquat. Org. 2014, 112, 9–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayley, A.E.; Hill, B.J.; Feist, S.W. Susceptibility of the European common frog Rana temporaria to a panel of ranavirus isolates from fish and amphibian hosts. Dis. Aquat. Org. 2013, 103, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Grayfer, L.; Andino, F.D.J.; Robert, J. Prominent Amphibian (Xenopus laevis) tadpole type iii interferon response to the frog virus 3 ranavirus. J. Virol. 2015, 89, 5072–5082. [Google Scholar] [CrossRef] [PubMed]
- Morales, H.D.; Abramowitz, L.; Gertz, J.; Sowa, J.; Vogel, A.; Robert, J. Innate immune responses and permissiveness to ranavirus infection of peritoneal leukocytes in the frog Xenopus laevis. J. Virol. 2010, 84, 4912–4922. [Google Scholar] [CrossRef] [PubMed]
- Wendel, E.S.; Yaparla, A.; Koubourli, D.V.; Grayfer, L. Amphibian (Xenopus laevis) tadpoles and adult frogs mount distinct interferon responses to the frog virus 3 ranavirus. Virology 2017, 503, 12–20. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a. Effects of Batrachochytrium dendrobatidis on amphibian hosts | ||||||||
| Species | IUCN status | Bd Strain | Dose (Total zoospores) | Life Stage | Effect on host | Reference | ||
| Agalychnis callidryas | LC | JEL 423 | 5 × 105 zoospores | na | Increased expression of genes of proteolitic enzymes | [216] | ||
| Alytes muletensis | VU | UKTvB, TF5al | 23,000 zoospores over two weeks | Through metamorphosis | Strain differences in infection | [93] | ||
| Alytes obstetricans | LC | na | na | Through metamorphosis | Population differences in survival | [217] ** | ||
| LC | na | Dose reported in the field | Larvae different cohorts | Mitigation of Bd with fungicide was transient not able to prevent spread of Bd | [218] | |||
| Ambystoma californiense | VU | JEL 270 | 1000 and 100,000 zoospores | Juveniles | No significant differences in survival or mass | [219] ** | ||
| Ambystoma laterale | LC | JEL 423, JEL 404 | 105–106 zoosporangia | Juveniles | No significant differences in survival | [86] | ||
| Ambystoma opacum | LC | 277 | 250,000 zoospores | Larvae | No infection detected, no significant differences in survival | [119] | ||
| Ambystoma tigrinum | LC | A-277, R-230 | 9,000,000 and 6,000,000 zoospores | Juveniles | No significant differences in survival | [220] | ||
| Bd-GPL isolate | 10,000 and 200,000 zoospores | Juveniles | No differences in zoospore outputs | [221] ** | ||||
| Amietia delalandii | LC | South Africa 1a and 1b, South Africa 2 and 3, UK 1 and 2, Spain and Sardinia | 1 × 106 zoospores | Adults (mucosome) | Skin mucosomes inhibited Bd growth | [222] | ||
| Anaxyrus americanus | LC | JEL 197 | 500,000 zoospores | Juveniles | Age dependent effect of Bd susceptibility | [116] | ||
| JEL 423, JEL 404 | 106–107 zoospores and 105–106 zoosporangia | Larvae | Reduced survival | [86] | ||||
| JEL 213 | 2.10 × 106 zoospores | Juveniles | Reduced survival | [172] | ||||
| JEL 660 | 1 × 105 zoospores | Juveniles | Elevated body temperatures | [223] | ||||
| Anaxyrus boreas | NT | JEL 215 | 12,600 zoospores | Larvae | Reduced survival | [98] | ||
| JEL 274 | 170,000 zoospores | Larvae | Higher stress hormones and increased length | [104] | ||||
| JEL 274 | 2 culture dishes inoculated in batches with 20 tadpoles | Larvae | Did not avoid infected conspecifics, increased activity, no differences in temperature selection | [108] | ||||
| JEL 274 | 100,000, 50,000, or 1000 zoospores | Larvae | No significant differences in survival | [80] | ||||
| JEL 274 | 100,000, 50,000, or 1000 zoospores | Juveniles | No significant differences in survival | [80] | ||||
| JEL 215 | 2.08 × 107/plate | Juveniles | Reduced survival | [144] * | ||||
| JEL 275 | 106 zoospores/toadlet daily | Juveniles | Mass dependent survival time, exposed toadlets held bodies out of water as much as possible | [99] | ||||
| JEL 275 | 5.8 × 105 zp/mL | Adults | Reduced survival | [152] | ||||
| JEL 275 | 1.13 × 106 zoospores | Adults | High infection intensity, loss in body weight, mild hyperkeratosis and perturbations in gene expression | [224] | ||||
| JEL 425, JEL 630, JEL 646, JEL 627 | 1 × 105 zoospores | Larvae | Increased mortality dependent on isolate | [91] | ||||
| JEL 423 | 2.0 × 106 zoospores | na | Bufadienolides extracted inhibited Bd growth | [225] | ||||
| Anaxyrus boreas boreas | LC | JEL 275 | 100,000 zoospores | Adults | Electrolyte alterations, lymphocytic infiltration | [226] ** | ||
| Anaxyrus fowleri | LC | na | na | Larvae | Reduced foraging efficiency | [119] | ||
| FMB 001 | 6,000,000 zoospores | Larvae | Negatively impacts growth | [109] | ||||
| USA isolate 284 | 6,000,000 zoospores | Larvae | Reduced foraging efficiency | [120] | ||||
| Anaxyrus terrestris | LC | JEL 274 | 2.6 × 105 zoospores | Juveniles | Reduced survival, decreased feeding | [100] | ||
| Anaxyrus woodhousii | LC | Bd-GPL isolate | 10,000 and 200,000 zoospores | Juveniles | No significant differences in zoospore outputs | [221] ** | ||
| Atelopus glyphus | CR | JEL 423 | 3 × 105 | na | Genes with elevated expression in infected individuals were enriched for GO terms, including cell adhesion, immune response and regulation of cell proliferation. | [216] | ||
| Atelopus varius | CR | JEL 410, JEL 412, JEL 413, and 3 contemporary isolates | 50 × 102 | Adults | No differences in infection intensity or survival by Bd strain | [227] | ||
| Atelopus zeteki | CR | JEL 423 | 30,000 zoospores | Adults | Infection intensity and zoospore output were positively correlated. | [228] | ||
| JEL 423 | 30,000 zoospores | Adults | Significant differences in expression of numerous genes involved in innate and inflamatory responses | [229] ** | ||||
| JEL 408 | 100 zoospores, 104, 106 | Adults | Dose and temperature dependent effects | [78] | ||||
| JEL 310 | 3000 zoospores | Juveniles | Probiotics use did not prevent or delay mortality by Bd. | [230] | ||||
| JEL 423 | 3 × 103 zoospores | na | Genes with increased expression were enriched for GO terms, including response to wounding, inflammatory response and apoptosis | [216] | ||||
| Batrachoseps attenuatus | LC | na | 3 × 109 zoospore equivalents | Adults | Cleared infection, wild caught infected individuals experienced 100% mortality in the laboratory | [84] ** | ||
| Bufo bufo | LC | IA042, IA043, 0711 (Pyrenees, BdGPL), VAo2, VAo4, VAo5 (Valencia, BdGPL lineage), CCB1, TF5a1 and TF1.1 (Mallorca, BdCAPE lineage) | 3000–17,000 active zoospores | Larvae | Strain differences in mortality and infection dynamics | [85] | ||
| UK Bd UKTvB, Mallorca Bd TF5a1, Pyrenneen Bd IA042 | 19,000 zoospores, 190 zoospores | Larvae | Reduced survival, differences in mass, strain differences in virulence and infection | [15] | ||||
| Bd-GPL IA-42 | 160, 16,000 zoospores | Juveniles | Reduced survival, mass-dependent effects | [196] | ||||
| IA2004 043 | 30 to 70, 3000 to 15,000 zoospores | Through metamorphosis | Dose-, size-, and age-dependent effects | [79] | ||||
| na | 120–300 zoospores, 12,000–30,000 zoospores | Juveniles | Warmer overwintering regime increases the probability of infection. Proliferation of Bd in the host was better in toadlets that experienced a colder winter | [81] | ||||
| Bufo marinus | LC | JEL 275 | 2.04 × 106 zoospores | Adults | Minimal hyperkeratosis, no differences in survival neither in body weight | [224] | ||
| Bufo quercicus (Anaxyrus quercicus) | LC | SRS 812 | 60,000 zoospores | Adults | Learned behavioral resistance to Bd | [83] | ||
| Craugastor fitzingeri | LC | JEL 423 | 5 × 105 zoospores | na | Genes with increased expression were enriched for GO terms, including response to wounding, inflammatory response and apoptosis. | [216] | ||
| Dendropsophus meridensis | EN | BdLEcat10CG-1 | 9 × 106 zoospores | Juveniles | Reduced survival | [231] ** | ||
| Dendrobates auratus | LC | na | na | Juveniles | Reduced survival | [36] | ||
| Dendrobates tinctorius | LC | na | na | Juveniles | Reduced survival, skin lesions | [36] | ||
| Desmognathus monticola | LC | JEL 197 | 1.068 × 107 zoospores | Adults | Reduced survival | [232] ** | ||
| Desmognathus orestes | LC | BD 197 | 1,000,000 zoospores | Adults | No clinical signs of infection | [233] ** | ||
| Eleutherodactylus coqui | LC | JEL 427 | 50,000 or 100,000 zoospores | Juveniles | Reduced survival, population differences | [94] ** | ||
| JEL 427 | 106 and 105 zp/mL in 10 mL | Adults | No significant differences in survival, cleared or reduced infection | [94]** | ||||
| Hyla chrysoscelis | LC | na | 7000 zoospores/mL | Through metamorphosis | No significant differences in survival, reduced metamorphic body mass, delayed time to metamorphosis | [234] | ||
| JEL 646, JEL 423, JEL 213, JEL 660, FMB 003, JEL 404 | 8 × 103 zoospores | Through metamorphosis | No significant differences in survival, growth, or time to metamorphosis | [235] | ||||
| na | 7000 zp/mL | Larvae | No significant differences in survival or larval period length, reduced body mass at metamorphosis | [118] | ||||
| na | 125,000 zoospores | Larvae | Reduced foraging efficiency | [119] | ||||
| na | 6,000,000 zoospores | Larvae | Reduced foraging efficiency | [120] | ||||
| Hyla cinerea | LC | JEL 423, SRS810 | 76.7 × 106, 4.7 × 106 zoospores | Juveniles and Adults | No clinical signs of infection. Infection did not negitively affect body condition or growth rate for either strain or lifestage | [89] ** | ||
| Hyla versicolor | LC | JEL 274 | 2.6 × 105 | Juveniles | Reduced survival | [100] | ||
| FMB 003 | 75,000 | Larvae | Reduced survival, age-dependent effects | [167] | ||||
| FMB 001 | 6,000,000 zoospores | Larvae | Negatively impacts growth | [109] | ||||
| Hypsiboas crepitans | LC | Bd1006 | 9,000,000 zoospores | Juveniles | Cleared infection | [82] | ||
| Ichthyosaura alpestris | LC | na | na | Adults | Reduced survival | [236] | ||
| Lechriodus fletcheri | LC | EPS4 | 750,000 zoospores | Sub-adults | Significant differences in survival, increased sloughing rates | [237] | ||
| Leiopelma archeyi | CR | JEL 197 | 250,000 zoospores | Adults | Cleared infection | [238] ** | ||
| Limnodynastes peronii | LC | Gibbo River-Llesueuri-00-LB-1 | 20 × 106 zoospores | Larvae and Juveniles | Reduced survival, infection loads increased over time | [239] | ||
| EPS4 | 750,000 zoospores | Adults | Low mortality rates, increase in sloughing rates | [237] | ||||
| Limnodynastes tasmaniensis | LC | GibboRiver-Llesueuri-00-LB-1 | 5000 zoospores + 2 mL water | Juveniles | No significant differences in survival | [240] | ||
| EPS4 | 750,000 zoospores | Adults | No significant differences in survival, sloughing rate increased at lower Bd loads | [237] | ||||
| Lissotriton helveticus | LC | na | ~2000 zoospores | Adults | Decreased mass, no evidence of hastened secondary sexual trait regression, exposure associated with a 50% earlier initiation of the terrestrial phase | [241] ** | ||
| Lithobates catesbeianus | LC | JEL 274 | 48,000 zoospores | Larvae | Higher stress hormones and increased length | [104] | ||
| JEL 215 | 8400 zoospores | Larvae | No significant differences in survival | [98] | ||||
| JEL 274, JEL 630 | 1.7 × 104 zoospores/mL in 15 mL | Juveniles | Strain differences in infection | [80] | ||||
| JEL 423 | 8 × 107 to 2 × 108 zoospores | Juveniles | Disruption of the epidermal cell maturation cycle | [35] ** | ||||
| JEL 423, JEL 404 | 106–107 zoospores and 105–106 zoosporangia | Juveniles | No significant differences in survival | [86] | ||||
| Bd-GPL isolate | 10,000 or 200,000 zoospores | Juveniles | Produces more infective zoospore stage than other species tested | [221] ** | ||||
| Crater Meadow isolate, Finley Lake isolate | 106 and 2 × 106 zoospores | Juveniles | No significant differences in survival, low infection prevalence, relatively low infection loads and lack of clinical disease for Finley Lake strain | [86] ** | ||||
| JEL 310 | 7 × 106 zoospores and 4.8 × 107 zoospores | Juveniles | Manipulation of frogs microbiota did not affect Bd infection intensity. | [242] | ||||
| Isolate from dead Alytes obstetricans | 150,000 zoospores | Larvae | No significant differences in survival | [236] | ||||
| Lithobates clamitans | LC | JEL 423, JEL 404 | 106–107 zoospores and 105–106 zoosporangia | Juveniles | Strain differences in infection | [86] | ||
| Lithobates pipiens | LC | JEL 423, JEL 404 | 106–107 zoospores and 105–106 zoosporangia | Juveniles | No significant differences in survival | [86] | ||
| JEL 423 | 3.98 × 106 zoospores | Juveniles | Increased skin shedding, no significant differences in survival or splenosomatic or hepatosomatic | [171] ** | ||||
| JEL 424 | 3.98 × 106 zoospores | Juveniles | indices, the densities and sizes of hepatic and splenic melanomacrophage aggregates, the density and size | [171] ** | ||||
| JEL 425 | 3.98 × 106 zoospores | Juveniles | of hepaticgranulomas, proportions of circulating leucocytes, the ratio of neutrophils to lymphocytes, | [171] ** | ||||
| JEL 426 | 3.98 × 106 zoospores | Juveniles | or the ratio of leucocytes to erythrocytes | [171] ** | ||||
| JEL 197 | 500,000 zoospores | Juveniles | No significant differences in survival regardless of age | [116] | ||||
| JEL 423 | 1.69 × 107–7.43 × 108 zoospores | Adults | Lower peak jumping velocity in infected subjects, testes width significantly greater in infected individuals | [243] ** | ||||
| Lithobates sphenocephalus | LC | na | 2.88 × 106 zoospores | Larvae | No significant differences in survival, reduced foraging efficiency | [117] | ||
| na | 400,000 zoospores | Larvae | Low protein diets resulted in smaller and less developed tadpoles and reduced immune responses, high protein diets significantly increased resistance to Bd | [244] | ||||
| JEL 197 | 106 zoospores | Juveniles | Increased pathogen skin burden within two weeks of exposure, higher pathogen burden in deceased frogs, decrease in pathogen loads over time | [245] | ||||
| Lithobates sylvaticus | LC | JEL 404, JEL 423 | 106–107 zoospores and 105–106 zoosporangia | Larvae | Reduced survival, no differences in growth or time to metamorphosis | [86] | ||
| JEL 404, JEL 423 | 106–107 zoospores and 105–106 zoosporangia | Larvae | Reduced survival | [86] | ||||
| JEL 197 | 104 zoospores | Juveniles | No significant differences in survival regardless of age | [116] | ||||
| JEL 274 | 2.6 × 105 zoospores | Juveniles | Reduced survival | [100] | ||||
| JEL 274 | 1.55 × 105 | Juveniles | Population differences in survival | [206] | ||||
| JEL 423 | 1 × 107 to 2 × 107 zoospores | Juveniles | Disruption of the epidermal cell maturation cycle | [35] ** | ||||
| Lithobates yavapaiensis | LC | Arizona Bd strain PsTr2004 | 1 × 105 zoospores | Juveniles | MHC heterozygosity as a predictor of survival | [246] | ||
| Litoria aurea | VU | Gibbo River-Llesueuri-00-LB-1 | 20 × 106 zoospores | Larvae and Juveniles | No significant differences in survival, decrease in pathogen loads over time | [239] | ||
| Litoria booroolongensis | VU | AbercrombieNP-L.booroolongensis-09-LB-P7) | 750,000 zp in 5 mL | Juveniles | No evidence that prior Bd infection increases protective immunity | [247] | ||
| Litoria caerulea | LC | GibboRiver-Llesueuri-00-LB-1 | 5000 zoospores + 2 mL water | Juveniles | Reduced survival | [240] | ||
| Strain 98 1469/10, Strain 99 1385/12, Strain 00 545 | 50,000 zoospores | Juveniles | Differences in survival rates among infected groups | [34] | ||||
| na | na | Adults | Decreased blood pH, low plasma osmolality and reduced concentrations of sodium, potassium, chloride and magnesium | [38] | ||||
| EPS4 | 250,000 zoospores | Adults | Increased skin sloughing rate with increased infection intensity | [248] ** | ||||
| Gibboriver-Llesueuri-00-LB-1P50 and P10 (passages) | 93 × 104/mL-1 | Adults | No significant differences in survival or mass | [96]** | ||||
| na | 250,000 zoospores | Adults | Impaired immune response | [249] ** | ||||
| na | na | Adults | Impaired stress and immune response, increased skin shedding | [103] * | ||||
| Paluma-Lseratta-2012RW-1 | 6 × 105 zoospores | Juveniles | Immunological profiles changed according to acclimated regime | [250] | ||||
| EPS4 and Waste point-Lverreauxii-2013-LB | 1.25 × 106 zoospores | Adults | Low mortality rates, increase in sloughing rates | [237] | ||||
| JEL 423 and Rio Maria isolate | 1.5 × 106 zoospores | Adults | No differences in infection intensity or survival by Bd strain | [227] | ||||
| JEL 423 and Rio Maria isolate | indirect | Adults | No differences in infection intensity or survival by Bd strain | [227] | ||||
| Litoria chloris | LC | GibboRiver-Llesueuri-00-LB-1 | 5000 zoospores + 2 mL water | Juveniles | Reduced survival | [240] | ||
| GibboRiver-Llesueuri-00-LB-1 | 15,000 zoospores + 2 mL water | Juveniles | Temperature did not influence leukocyte populations | [240] | ||||
| na | 15,000 zoospores | Juveniles | Temperature dependent effects on survival | [77] *** | ||||
| Litoria infrafrenata | LC | na | 250,000 zoospores | Adults | Reduction in white blood cells and serum globulin concentrations | [249] ** | ||
| Litoria raniformis | EN | na | 100,000 zoospores | Adults | Compromised ability to osmoregulate and rehydrate, no significant difference in metabolic or breathing rates | [251] ** | ||
| Litoria verreauxii alpina | LC | AbercrombieNP-L.booroolongensis-09-LB-P7) | 750,000 zoospores | Adults | No effect of MHC heterozygosity or allelic divergence on survival | [252] | ||
| AbercrombieR-L.booroologensis-2009-LB1 and WastePoint-L.v.alpina-2013-LB2 | 1 × 106 zoospores in 3 mL and 5 × 105 zoospores in 10 mL | Adults | Oogenesis and spermatogenesis increased in infected animals | [253] | ||||
| Mixophyes fasciolatus | LC | GibboRiver-Llesueuri-00-LB-1 | 5000 zoospores + 2 mL water | Juveniles | Reduced survival | [240] | ||
| No. 00/545 | 1000 zoospores | Adults | Lower temperatures enhanced pathogenicity | [76] * | ||||
| Osteopilus septentrionalis | LC | SRS 812 | 3 × 104 zp/mL in 2 mL | Larvae | The loss of keratin in the mouthparts associated with a loss of Bd | [254] | ||
| SRS 812 | 3 mL of 6 × 104 (after each water change) | Larvae | Reduced survival | [170] | ||||
| SRS 812 | 3 × 106 zp/mL | Juveniles | Pathogen loads decreased over time; increased lymphocyte proliferation with increased exposures; previous exposure increased chances of survival | [83] | ||||
| Pelophylax esculentus | LC | TG 739 | 1.5–2 × 105 zoospores | Adults | Reduction in skin peptide and microbiota immune defenses caused less weight gain and increased infection rates. | [255] ** | ||
| Pelophylax lessonae | LC | TG 739 | 1.5–2 × 105 zoospores | Adults | Reduction in skin peptide and microbiota immune defenses caused less weight gain and increased infection rates. | [255] ** | ||
| Platyplectrum ornatum | LC | EPS4 | 750,000 zoospores | Adults | Significant differences in survival | [237] | ||
| Plethodon cinereus | LC | JEL 660/JS OH-1 | 7 × 105 in 5 mL | Adults | Increased feeding activity | [121] * | ||
| Plethodon glutinosus | LC | BD 197 | 1,000,000 zoospores | Adults | Clinical symptoms of infection | [233] ** | ||
| BD 197 | 10,000 or 100,000 zoospores | Adults | No significant differences in survival | [233] ** | ||||
| Plethodon metcalfi | LC | JEL 197 | 1.068 × 107 zoospores | Adults | Reduced survival | [232] ** | ||
| Plethodon shermani | JEL 197 | 1 × 107 zoospores | Adults | Decreased body mass, reduction in locomotory activity | [256] | |||
| Pseudacris crucifer | LC | JEL 423, JEL 404 | 106–107 zoospores and 105–106 zoosporangia | Adults | No significant differences in survival | [86] | ||
| Pseudacris feriarum | LC | JEL 274 | 2.6 × 105 zoospores | Juveniles | Reduced survival | [100] | ||
| Pseudacris regilla | LC | JEL 215 | 12,600 zoospores | Larvae | No significant differences in survival | [98] | ||
| JEL 626 | 27,800 zoospores | Larvae | Reduced survival and activity, delayed time to metamorphosis | [169] | ||||
| JEL 215 | 2 culture dishes inoculated in batches with 20 tadpoles | Larvae | No differences in temperature selection | [108] | ||||
| JEL 216 | 6.18 × 106/mL | Larvae | No significant differences in activity or refuge use | [115] | ||||
| JEL 274 | 100,000, 50,000, or 1000 zoospores | Larvae | No significant differences in survival, dose-dependent infection loads | [80] | ||||
| JEL 274 | 100,000, 50,000, or 1000 zoospores | Juveniles | Reduced survival, dose-dependent infection loads | [80] | ||||
| JEL 215 | 2.08 × 107 zoospores | Juveniles | No significant differences in survival | [144] * | ||||
| JEL 274 | 50,000 zoospores | Juveniles | Reduced survival, Infection load increased over time, lower lymphocyte levels | [257] | ||||
| JEL 274 | 2.6 × 107 and 1.1 × 106 zoospores/L | Through metamorphosis | Dose-dependent effects | [74] | ||||
| JEL 425, JEL 630, JEL 646 | 1 × 105 zoospores | Larvae | No significant differences in survival | [91] | ||||
| Pseudacris triseriata | LC | 27-mile lake isolate, Lost lake isolate | 8 × 104 zoospores | na “frogs” | Strain differences in infection | [88] | ||
| LC | Bd-GPL isolate | 10,000 and 200,000 zoospores | Juveniles | No significant differences in zoospore outputs | [221] | |||
| Pseudophryne corroboree | CR | AbercrombieR-L.booroologensis-2009-LB1 | 1 × 106 zoospores in 3 mL | Adults | Oogenesis and spermatogenesis increased in infected animals | [253] | ||
| Pyxicephalus adspersus | LC | South Africa 1a and 1b, South Africa 2 and 3, UK 1 and 2, Spain and Sardinia | 1 × 106 zoospores | Adults (mucosome) | Skin mucosomes inhibited Bd growth | [222] | ||
| Rana aurora | LC | JEL 215 | 2 culture dishes inoculated in batches with 20 tadpoles | Larvae | No differences in temperature selection | [108] | ||
| na | 2 × 105 zp added every other day for 8 days | Larvae | High temperature variability in the presence of Bd had decreased growth | [149] | ||||
| JEL 216 | 6.18 × 106/mL | Larvae | No significant differences in activity or refuge use | [115] | ||||
| Rana blairi/Rana sphenocephala (Lithobates blairi/Lithobates sphenocephala) | na | na | 7000 zp/mL | Larvae | No significant differences in survival, reduced metamorphic body mass | [118] | ||
| Rana boylii | NT | LJR 119 | 9.4 × 106 zoospores in 50 mL | Juveniles | No significant differences in survival, reduced growth, increased skin peptide concentrations | [165] * | ||
| A-227, R-230 | 1,275,000; 127,500 zoospores | Juveniles | No significant differences in survival | [220] | ||||
| Rana cascadae | LC | JEL 215 | 12,600 zoospores | Larvae | No significant differences in survival, increased incidence of mouthpart abnormalities | [98] | ||
| JEL 274 | 48,000 zoospores | Larvae | Higher stress hormones and increased length and mass | [104] | ||||
| JEL 274 | 50,000 zoospores | Larvae | No significant differences in mortality, Infection load decreased over time, stronger bacterial killing response over time, elevated neutrophil levels | [257] | ||||
| JEL 274 | 4 culture dishes inoculated in batches with 90 tadpoles | Larvae | Non-infected individuals were observed more frequently on Bd+ side of test chamber | [108] | ||||
| JEL 216 | 6.18 × 106/mL | Larvae | No significant differences in activity or refuge use | [115] | ||||
| JEL 274 | 100,000, 50,000, or 1000 zoospores | Larvae | No significant differences in survival | [80] | ||||
| JEL 274 | 100,000, 50,000, or 1000 zoospores | Juveniles | Reduced survival | [80] | ||||
| JEL 215 | 2 culture dishes inoculated in batches with 20 tadpoles | Juveniles | No differences in temperature selection | [108] | ||||
| JEL 274 | 8.5 × 104 zp | Juveniles | Lower stress hormone levels | [104] | ||||
| Section line lake and Carter Meadow | 2.2 × 105 zoospores | Juveniles | Strain differences in mortality and infection dynamic, no differences in survivorship between populations BUT Bd prevalence and infection intensity differed between populations | [92] | ||||
| JEL 215 | 2.08 × 107 zoospores | Juveniles | Reduced survival | [144] * | ||||
| JEL 425, JEL 630, JEL 646 | 1 × 105 zoospores | Larvae | No significant differences in survival | [91] | ||||
| Rana draytonii | VU | JEL 270 | 1000 and 100,000 zoospores | Juveniles | No significant differences in survival or mass | [219] ** | ||
| Rana muscosa | EN | JEL 217 | 3.6 × 109 zoospores | Larvae | Infected but appear healthy, loss of mouth pigmentation | [208] ** | ||
| JEL 217 | na | Larvae | Transmitted infection to each other and to post-metamorphic individuals | [208] ** | ||||
| LJR089 | 1 × 107 zoospores | Larvae | Proportion of hosts that became infected increased with the number of previously infected R. muscosa tadpoles to which they were exposed | [73] | ||||
| na | >100,000 in 1 mL | Adults | Disruption of skin integrity, ion imbalance | [258] | ||||
| LJR089 | 1 × 107 zoospores | Juveniles | Temperature dependent effects on survival, increased skin shedding | [75] | ||||
| Rana Once (Lithobates Onca) | EN | CJB7 from Rana muscosa and SLL from Rana cascadae | 3 × 106 | Juveniles | No significant differences in survival, cleared infection | [259] | ||
| Rana pipiens (Lithobates pipiens) | LC | na | 2,800,000 zoospores | Larvae | Reduced activity | [72] | ||
| JEL 275 | 104 zoospores | Juveniles | Reduced survival | [260] ** | ||||
| JEL 274 | 2.6 × 105 zoospores | Juveniles | Reduced survival | [115] | ||||
| Rana sierrae | EN | TST75,CJB4, CJB5, CJB7 | 200,000 zoospores | Juveniles | Altered microbiome | [261] ** | ||
| Rana temporaria | LC | BdGPL IA-42 | 160 and 16,000 zoospores | Juveniles | No significant differences in survival, high dose resulted in less weight gain or weight loss | [196] | ||
| Isolate IA 042 | 100,000 zoospores | Juveniles | Significant transcriptional response to Bd | [262] | ||||
| Rana yavapaiensis (Lithobates yavapainensis) | LC | A-277, R-230 | 8.5 × 103 zoopores/mL | Juveniles | No significant differences in survival | [220] | ||
| Silurana tropicalis (Xenopus tropicalis) | LC | IA042 | 106 zoospores | Adults | Temperature dependent effects on immune response | [263] ** | ||
| na | na | Adults | Altered gene expression to physiological and immunological genes | [264] ** | ||||
| Xenopus laevis | LC | JEL 197 and JEL 275 | na | Adults | Impaired lymphocyte proliferation and induced splenocyte apoptosis | [265] | ||
| JEL 197 and JEL 275 | 106 zoospores | Adults | Peptide-depleted frogs became more susceptible to Bd infection with higher burdens and weight loss | [266] ** | ||||
| JEL 197 | 107 zoospores | Adults | Inhibition of local lymphocyte responses in host to promote infection | [267] | ||||
| b. Effects of Batrachochytrium salamandrivorans on amphibian hosts | ||||||||
| Species | IUCN Status | Bsal Strain | Bsal Dose (Total zoospores) | Life Stage | Effect on Host | Reference | ||
| Alytes obstetricans | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| AMFP13/1 | 5000 in 1 mL | Adults | No significant effect | [18] | ||||
| AMFP13/1, AMFP14/1, AMFP14/2, AMFP15/1 | 105 | Juvenile | No signs of disease but able to transmit infection after 14 days | [18] | ||||
| Ambystoma maculatum | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| Ambystoma opacum | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] ** | ||
| Bombina variegata | LC | AMFP13/1 | 5000 in 1 mL | Adults | No infection or disease detected | [42] | ||
| Cynops pyrrhogaster | LC | AMFP13/1 | 5000 in 1 mL | <1 year | Susceptible to infection and disease | [42] | ||
| Discoglossus scovazzi | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| Epidalea calamita | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| Euproctus platycephalus | EN | AMFP13/1 | 5000 in 1 mL | Adults | Reduced survival, confirmed invasion of the skin | [42] | ||
| Gyrinophilus porphyriticus | LC | AMFP13/1 | 5000 in 1 mL | Adults | No infection or disease detected | [42] | ||
| Hyla arborea | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| Hynobius retardatus | LC | AMFP13/1 | 5000 in 1 mL | Adults | No infection or disease detected | [42] | ||
| Hypselotriton cyanurus | LC | AMFP13/1 | 5000 in 1 mL | Adults | Susceptible to infection and disease | [42] ** | ||
| Ichthyosaura alpestris | LC | AMFP13/1 | 5000 in 1 mL | <1 year | Reduced survival, confirmed invasion of the skin | [42] | ||
| AMFP13/1 | 104, 103, 102, 10 | Juvenile | High doses resulted in mortality, previous infection offered no protection on reinfection | [268] | ||||
| Lissotriton helveticus | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| Lissotriton italicus | LC | AMFP13/1 | 5000 in 1 mL | <1 year | Reduced survival | [42] | ||
| Lithobates catesbeianus | LC | AMFP13/1 | 5000 in 1 mL | Adults | No infection or disease detected | [42] | ||
| Neurergus crocatus | VU | AMFP13/1 | 5000 in 1 mL | Adults | Reduced survival, confirmed invasion of the skin | [42] | ||
| Notophthalmus viridescens | LC | AMFP13/1 | 5000 in 1 mL | Adults | Reduced survival, confirmed invasion of the skin | [42]** | ||
| Pachyhynobius shangchengensis | VU | AMFP13/1 | 5000 in 1 mL | Adults | No infection or disease detected | [42] | ||
| Paramesotriton deloustali | VU | AMFP13/1 | 5000 in 1 mL | Adults | Susceptible to infection and disease | [42] | ||
| Pelobates fascus | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| Plethodon glutinosus | LC | AMFP13/1 | 5000 in 1 mL | Adults | Confirmed infection of the skin, no disease detected | [42] ** | ||
| Pleurodeles waltl | NT | AMFP13/1 | 5000 in 1 mL | <1 year | Reduced survival, confirmed invasion of the skin | [42] | ||
| Rana temporaria | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| Salamandra salamandra | LC | AMFP13/1 | 5000 in 1 mL | Adults | Reduced survival, ataxia. Cohousing effectively transmits infection | [18] | ||
| AMFP13/1 | 5000 in 1 mL | Adults | Warmer temperatures can clear infection | [269] | ||||
| AMFP13/1 | 105 in 1 mL | Adults | Topical treatments can reduce fungal loads and in combination with warmer temperature can clear infection | [269] | ||||
| AMFP13/1 | 5000 in 1 mL | <1 year | Reduced survival, confirmed invasion of the skin | [42] | ||||
| AMFP13/1, AMFP14/1, AMFP14/2, AMFP15/1 | 100 spores (low), 104 (high) | Juvenile | Mortality was delayed in low dose treatment | [268] | ||||
| na | 2.6 × 104, 1.3 × 104 | na | Mortality was delayed in low temp treatment | [268] | ||||
| AMFP13/1 | 103 | na | Reinfection did not change disease dynamics | [268] | ||||
| Salamandrella keyserlingii | LC | AMFP13/1 | 5000 in 1 mL | Adults | Confirmed infection but no effects of disease or on survival | [42] | ||
| Salamandrina perspicillata) | LC | AMFP13/1 | 5000 in 1 mL | <1 year | Reduced survival | [42] | ||
| Silurana tropicalis | LC | AMFP13/1 | 5000 in 1 mL | <1 year | No infection or disease detected | [42] | ||
| Siren intermedia | LC | AMFP13/1 | 5000 in 1 mL | Adults | Confirmed infection but no effects of disease or on survival | [42] | ||
| Speleomantes strinatii | NT | AMFP13/1 | 5000 in 1 mL | Adults | Reduced survival | [42] ** | ||
| Taricha granulosa | LC | AMFP13/1 | 5000 in 1 mL | <1 year | Reduced survival | [42] | ||
| Triturus cristatus | LC | AMFP13/1 | 5000 in 1 mL | <1 year | Reduced survival, confirmed invasion of the skin | [42] | ||
| Tylototriton wenxianensis | VU | AMFP13/1 | 5000 in 1 mL | <1 year | Reduced survival | [42] | ||
| Typhlonectes compressicauda | LC | AMFP13/1 | 5000 in 1 mL | Adults | No infection or disease detected | [42] | ||
| c. Effects of ranavirus on amphibian hosts | ||||||||
| Species | IUCN Status | Rv Strain | Dose | Type of Exposure | Life-Stage | Effect on Host | Reference | |
| Ambystoma californiense | VU | ATV | 200 uL of inoculum w/1000 virions of ATV in APBS solution | Injection | Adults | Reduced survival | [270] ** | |
| Ambystoma gracile | LC | ATV | na | Water bath | Larvae | Reduced survival | [128] * | |
| Ambystoma maculatum | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival, strain differences in infection | [132] | |
| Ambystoma mavortium | na | ATV | 1 × 103.3 and 7.1 × 103 TCID50/mL (1.4 million virions per animal) | Water bath | Larvae | Population differences in infection | [133] | |
| Ambystoma opacum | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival, strain differences in infection | [132] | |
| Ambystoma talpoideum | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | No difference in survival, no difference in infection | [132] | |
| Ambystoma tigrinum | LC | ATV (ATV-DO211) | 102, 102.5, 103, 103.5, 104, 105 PFU from original plaque assay of 4.5 × 107 | Water bath | Larvae | Dose dependent infection and survival rates | [70] | |
| ATV | 2 × 106 from 200 mL of 104 PFU/mL in aged tap water | Water bath | Larvae | No differences between transmission rates | [56] | |||
| ATV | 2 × 107 of ATV for a final concentration of 6.67 × 104 PFU/mL | Water bath with pond sediment | Larvae | No infection when exposed to virus in dried substrate, but when substrate was kept moist they became infected and experienced reduced survival | [56] | |||
| ATV | 500 PFU in 200 uL | Injection | Larvae | 1s ventral surface to ventral surface contact results in infection | [56] | |||
| ATV | 4 × 106 PFU from 400 mL of 104 PFU/mL in aged tap water | Water bath | Larvae | Infection rate increases with time and increased SVL | [56] | |||
| ATV | 103 PFU/mL, 104 PFU/mL | Water bath | Larvae | Temperature influences infectivity, survival, and time to death. Sublethal infections result in viral carrier status. | [130] | |||
| ATV | 102, 102.5, 103, 103.5, 104, 105 PFU from original plaque assay of 4.5 × 107 | Water bath | Larvae | Dose and developmental stage dependent infection rates | [70] | |||
| FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |||
| ATV | 103 PFU/mL | Water bath | Larvae | No differences in survival rates between larvae and juveniles | [56] | |||
| ATV | 103 PFU/mL | Water bath | Juveniles | Reduced survival | [56] | |||
| Ambystoma mavortum | na | ATV | 200 uL of inoculum w/1000 virions of ATV in APBS solution | Injection | Adults | Reduced survival | [270] * | |
| Ambystoma tigrinum nebulosum | na | ATV | 200 uL of inoculum w/1000 virions of ATV in APBS solution | Injection | Adults | Reduced survival | [270] | |
| Ambystoma tigrinum stebbinsi | na | ATV | 200 uL of inoculum w/1000 virions of ATV in APBS solution | Injection | Adults | Reduced survival | [270] * | |
| Anaxyrus americanus | LC | FV3-like isolate | 103 PFU/mL | Water bath | Embryo through metamorphosis | Developmental stage dependent infection and survival rates | [136] | |
| FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |||
| Anaxyrus boreas | LC | FV3-like isolate | 103 PFU | Water bath | Larvae | 100% mortality | [55] | |
| FV3-like isolate | 103 PFU | Water bath | Juveniles | 100% mortality | [271] | |||
| Bufo bufo | LC | RUK 11, RUK 13, BUK 2, BUK 3 | 106 pfu, 104 pfu [all exposures standardized to 30 mL] | Water bath | Larvae | Reduced survival, dose dependent infection and survival, strain differences in infection | [272] | |
| Cophixalus ornatus | LC | BIV | 103 TCID50/mL | Water bath, Injection, contact | Adults | Reduced survival | [273] * | |
| Gastrophryne carolinensis | LC | FV3 and FV3-like isolate | 106 PFUs in 10 uL of Eagle’s MEM | oral dose, Water bath | Larvae | No differences in survival and no strain differences in viral load | [126] | |
| FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |||
| Hyla chrysoscelis | LC | FV3-like isolate | 103 PFU/mL | Water bath | Embryo through metamorphosis | Reduced survival | [136] | |
| FV3 and FV3-like isolate | 106 PFUs in 10 uL of Eagle’s MEM | Oral dose, Water bath | Larvae | Reduced survival, exposure type dependent effects on survival and infection | [126] | |||
| FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |||
| FV-3 | 103 PFU/mL | Water bath | Larvae | Transmission can occur between vertebrate classes. Amphibian larvae more susceptible to ranavirus than other vertebrate classes. | [62] | |||
| Limnodynastes terraereginae | LC | BIV | 100, 101, 102.5, and 104 TCID50/mL (bath); 0.1 mL of 103 TCID50/mL (injection) | Water bath, Injection | Larvae | Reduced survival, renal, hepatic, splenic, and pulmonary necrosis | [274] * | |
| BIV | 100, 101, 102.5, and 104 TCID50/mL (bath); 0.1 mL of 103 TCID50/mL (injection) | Water bath, Injection | Juveniles | Reduced survival, renal, hepatic, splenic, and pulmonary necrosis | [274] * | |||
| Lithobates catesbeianus | LC | ATV | Tadpoles were fed infected salamander | feeding | Larvae | Reduced survival | [128] * | |
| FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | No differences in survival | [132] | |||
| ATV | 200 uL ATV/EPC which had 4 × 105 PFU/mL for adults injection. | Injection | Adults | Reduced survival | [128] * | |||
| Lithobates clamitans | LC | FV3-like isolate | 103 PFU/mL | Water bath | Embryo through metamorphosis | Reduced survival | [136] | |
| Lithobates palustris | LC | FV3 and FV3-like isolate | 106 PFUs in 10 uL of Eagle’s MEM | oral dose, Water bath | Larvae | Reduced survival, exposure type dependent effects on survival and infection | [126] | |
| Lithobates pipiens | LC | FV3-like isolate | 103 PFU/mL | Water bath | Embryo through metamorphosis | Reduced survival | [136] | |
| FV3 strains (SSME, wt-FV3, aza-C) | 50 mL of water with 10,000 PFU/mL | Water bath | Larvae | Strain dependent effects on survival | [275] * | |||
| FV3 isolate (wt-FV3), azacR, SsMeV | 50 mL of water with 10,000 PFU/mL | Water bath | Larvae | Infection dependent on temperature and strain | [129] | |||
| ATV | 100 uL of ATV/EPC which had 4 × 105 PFU/mL in EPC cells | Injection | Adults | Reduced survival | [128] * | |||
| Lithobates sevosus | CR | FV3-like isolate | 400 mL of water with 103 PFU/mL | Water bath, Injection, oral dose | Adults | Reduced survival, exposure type dependent effects on survival | [276] | |
| FV3-like isolate | 103 PFU | Water bath | Eggs | Reduced survival | [271] | |||
| FV3-like isolate | 103 PFU | Water bath | Hatchling | 100% mortality | [271] | |||
| FV3-like isolate | 103 PFU | Water bath | Larvae | 100% mortality | [271] | |||
| FV3-like isolate | 103 PFU | Water bath | Juveniles | 100% mortality | [271] | |||
| FV3-like isolate | 103 PFU | Water bath | Juveniles | Reduced survival | [271] | |||
| FV3-like isolate | 103 PFU | Water bath | Adults | Reduced survival | [271] | |||
| Lithobates sylvaticus | LC | FV3-like isolate | 103 PFU/mL | Water bath | Embryo through metamorphosis | Reduced survival | [136] | |
| FV3 isolate (wt-FV3), azacR, SsMeV | 50 mL of water with 10,000 PFU/mL | Water bath | Larvae | Infection dependent on temperature and strain | [129] | |||
| na | na | contact and feeding on infected individuals | Larvae | Reduced survival | [57] * | |||
| na | na | Exposure to contaminated sediment and Water | Larvae | Reduced survival | [57] | |||
| Litoria caerulea | LC | BIV | 103 TCID50/mL; 104.5 TCID50/mL | Water bath, Injection | Juvenile | Reduced survival, exposure type dependent effects on survival | [273] * | |
| BIV | 103 TCID50/mL | Water bath, Injection, contact | Adults | No differences in survival | [273] * | |||
| Litoria inermis | LC | BIV | 103 TCID50/mL | Injection | Adults | Tested negative for infection | [273] * | |
| Litoria latopalmata | LC | BIV | 103 TCID50/mL | Injection | Larvae | Reduced survival, renal, hepatic, splenic, and pulmonary necroses | [274] * | |
| LC | BIV | 103 TCID50 mL | Injection | Juveniles | Reduced survival, renal, hepatic, splenic, and pulmonary necrosis | [274] * | ||
| Litoria rubella | LC | BIV | 104.5 TCID50/mL | Injection | Adults | No differences in survival | [273] * | |
| Notophtalmus viridescens | LC | ATV | na | contaminated Water | Larvae | Reduced survival | [128] * | |
| LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | ||
| Pseudacris brachyphona | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |
| Pseudacris feriarum | LC | FV3-like isolate | 103 PFU/mL | Water bath | Embryo through metamorphosis | Reduced survival | [136] | |
| FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |||
| Pseudacris triseriata | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |
| Rana capito (Lithobates capito) | NT | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |
| Rana clamitans (Lithobates clamitans) | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |
| Rana latastei | VU | FV3 | 2.25 × 106 pfu/mL (aliquots of 10 mL) from 70 mL of stock solution with 5.5 × 108 PFU/mL added to aged tap water | Larvae | Reduced survival | [124] | ||
| FV3 | 4.5 × 106 pfu/mL (aliquots of 10 mL), 4.5 × 105, 4.5 × 104, 4.5 × 103, 4.5 × 102 | Larvae | Dose dependent survival and survival rates | [124] | ||||
| FV3 | na, but feeder tadpoles infected with 4.5 × 106 PFU/mL | Consuming infected carcasses | Larvae | Exposure type dependent survival rate | [124] | |||
| FV3 | 4.5 × 104 PFU/mL, 4.5 × 106 PFU/mL (this was achieved by adding 2.796 × 108 PFU of FV3 to 615 mL of aged water, low exposure was a 1:100 dilution of this.) | Larvae | Dose dependent survival, effect of genetic diversity on survival | [207] | ||||
| Rana palustris (Lithobates palustris) | LC | FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |
| Rana pipiens (Lithobates palustris) | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |
| Rana sphenocephala (Lithobates Sphenocephala) | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |
| Rana sylvatica (Lithobates sylvatica) | LC | FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |
| FV3-like isolates (from wood frog and spotted salamander) | 10 fold dilutions from 2.36 × 101 through 2.36 × 105 PFU/mL for wood frog isolate and 2.51 × 101 through 2.51 × 105 PFU/mL for spotted salamander isolate) | Water bath | Larvae | Dose dependent survival rates, no strain differences in infection | [105] | |||
| FV3-like isolate | 2.36 × 103 PFU/mL | Water bath | Larvae | Higher stress hormone levels | [105] | |||
| FV3 | 67; 670; and 6,700 PFU/mL | Water bath | Larvae | Horizontal transmission the most likely means of FV3 transmission | [60] | |||
| Rana temporaria | LC | RUK 11, RUK 13, BUK 2, BUK 3 | 106 pfu, 104 pfu [all exposures standardized to 30 mL] | Water bath | Larvae | Dose and strain dependent effects on survival | [272] | |
| BIV, DFV, ECV, EHNV, FV3, GV6, PPIV, REV, and SERV | 104 TCID50/mL | Water bath | Larvae | Strain and temperature dependent effects on survival | [277] | |||
| BIV, DFV, ECV, EHNV, FV3, GV6, PPIV, REV, and SERV | 104 TCID50/mL | Water bath | Juveniles | Strain dependent effects on survival | [277] | |||
| RUK11 and RUK13 | 0.25 mL intraperitoneally, 0.25 subcutaneously both from 106∙2 and 105∙6 TCID 50/mL stock | Injection | Adults | Reduced survival | [125] ** | |||
| Scaphiopus holbrookii | LC | FV3-like isolate | 103 PFU/mL | Water bath | Embryo through metamorphosis | Reduced survival | [136] | |
| FV3, FV3-like isolate | 103 PFU/mL | Water bath | Larvae | Reduced survival | [132] | |||
| Taudactylus acutirostris | CR | BIV | 103 TCID50/mL | Water bath | Adults | Reduced survival | [273] * | |
| Xenopus laevis | LC | FV3 | 1 × 104 PFU in 10 uL | Injection | Larvae | Developmental stage differences in immune response to FV3 | [278] | |
| FV3 | 5 × 106 PFU in 100 uL | Injection | Adults | Developmental stage differences in immune response to FV3 | [278] | |||
| FV3 | 1 × 104 PFU in 10uL for injection; 10 uL of 1 × 105 PFU for oral ingestion; and 2 mL of 5 × 106 PFU for water bath | Water bath, Injection, oral ingestion | Larvae | Developmental stage dependent immune function and infection rates | [134] | |||
| FV3 | 0.1 mL volume of 1 × 106 PFU | Injection | Juveniles | Developmental stage dependent immune function and infection rates | [134] | |||
| FV3 | 1 × 106 to 5 × 106 PFU in 300 uL | Injection | Adults | Host cell differences in viral clearance | [279] | |||
| FV3 | 1 × 106 PFU | na | Adults | Immunocompromised adults can transmit infection within 3 h | [134] | |||
| FV3 | 106 PFU | Injection | Larvae & Adults | Developmental stage differences in immune response to FV3 | [280] | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blaustein, A.R.; Urbina, J.; Snyder, P.W.; Reynolds, E.; Dang, T.; Hoverman, J.T.; Han, B.; Olson, D.H.; Searle, C.; Hambalek, N.M. Effects of Emerging Infectious Diseases on Amphibians: A Review of Experimental Studies. Diversity 2018, 10, 81. https://doi.org/10.3390/d10030081
Blaustein AR, Urbina J, Snyder PW, Reynolds E, Dang T, Hoverman JT, Han B, Olson DH, Searle C, Hambalek NM. Effects of Emerging Infectious Diseases on Amphibians: A Review of Experimental Studies. Diversity. 2018; 10(3):81. https://doi.org/10.3390/d10030081
Chicago/Turabian StyleBlaustein, Andrew R., Jenny Urbina, Paul W. Snyder, Emily Reynolds, Trang Dang, Jason T. Hoverman, Barbara Han, Deanna H. Olson, Catherine Searle, and Natalie M. Hambalek. 2018. "Effects of Emerging Infectious Diseases on Amphibians: A Review of Experimental Studies" Diversity 10, no. 3: 81. https://doi.org/10.3390/d10030081