The Effects of Asymmetric Diurnal Warming on Vegetation Growth of the Tibetan Plateau over the Past Three Decades

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
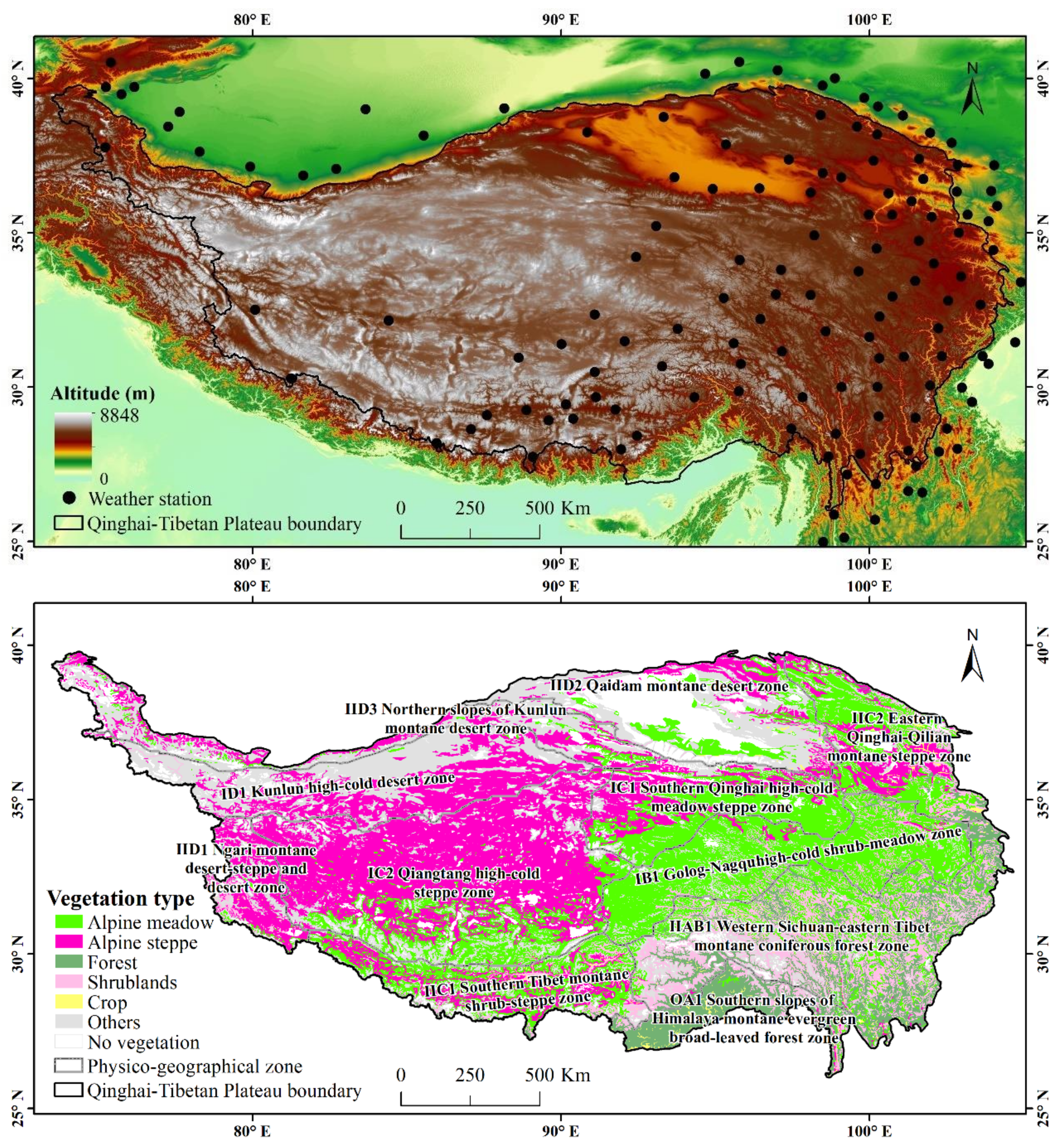
2.1. Data Sources and Processing
2.2. Data Analysis
2.2.1. Linear Regression Trends
2.2.2. Partial Correlation Analysis
3. Results
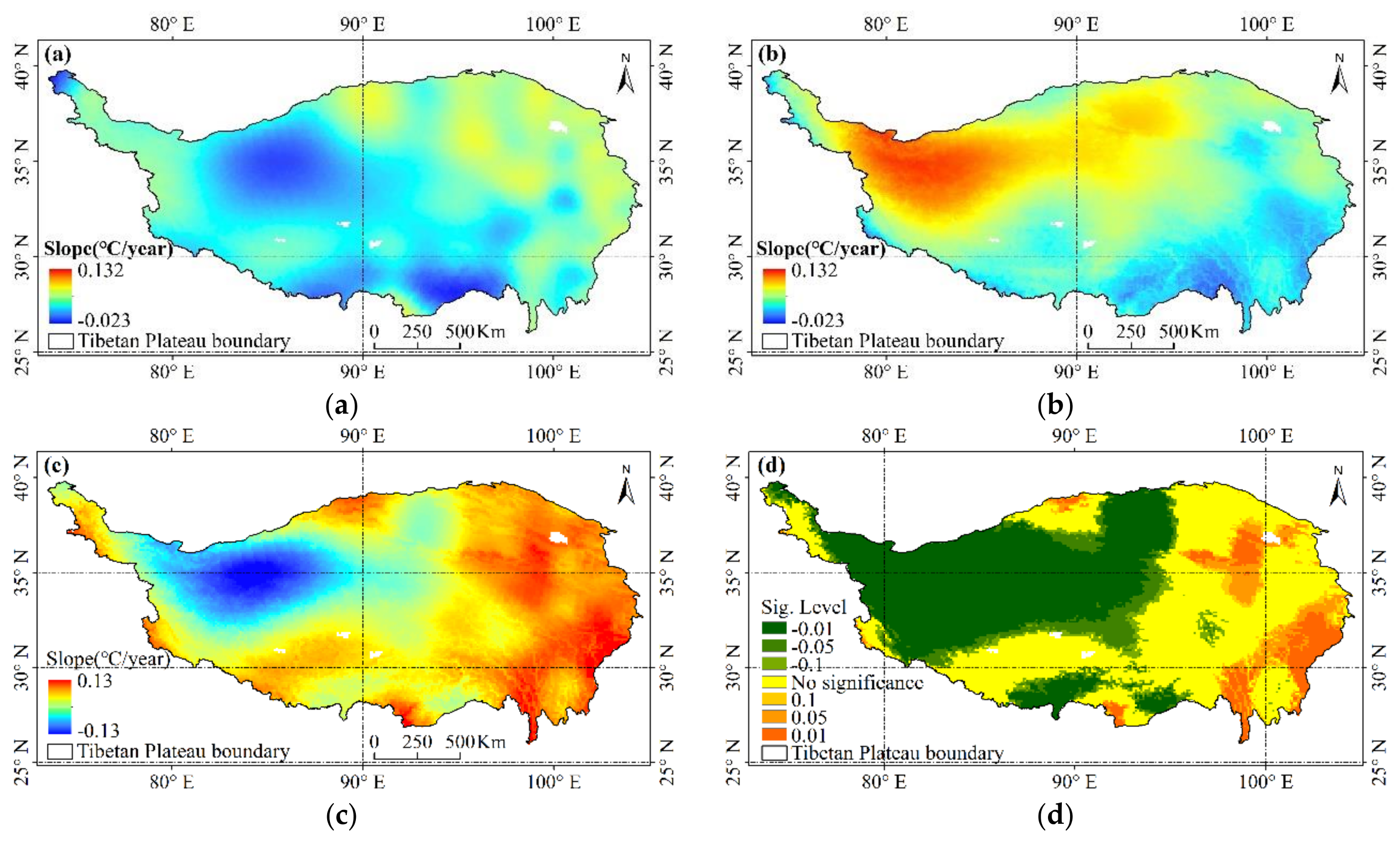
3.1. Trends of Daytime and Night-Time Warming in Tibetan Plateau
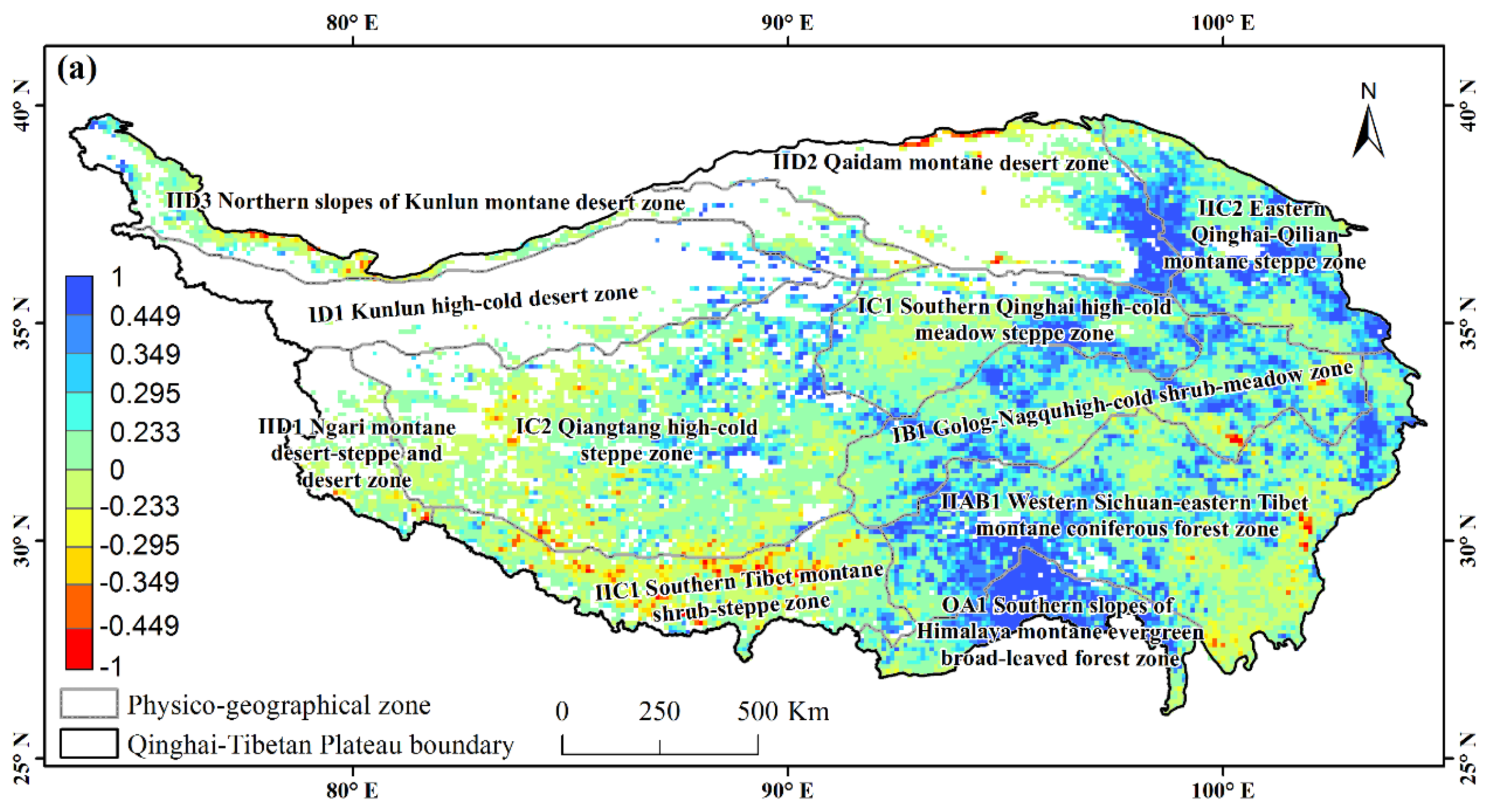
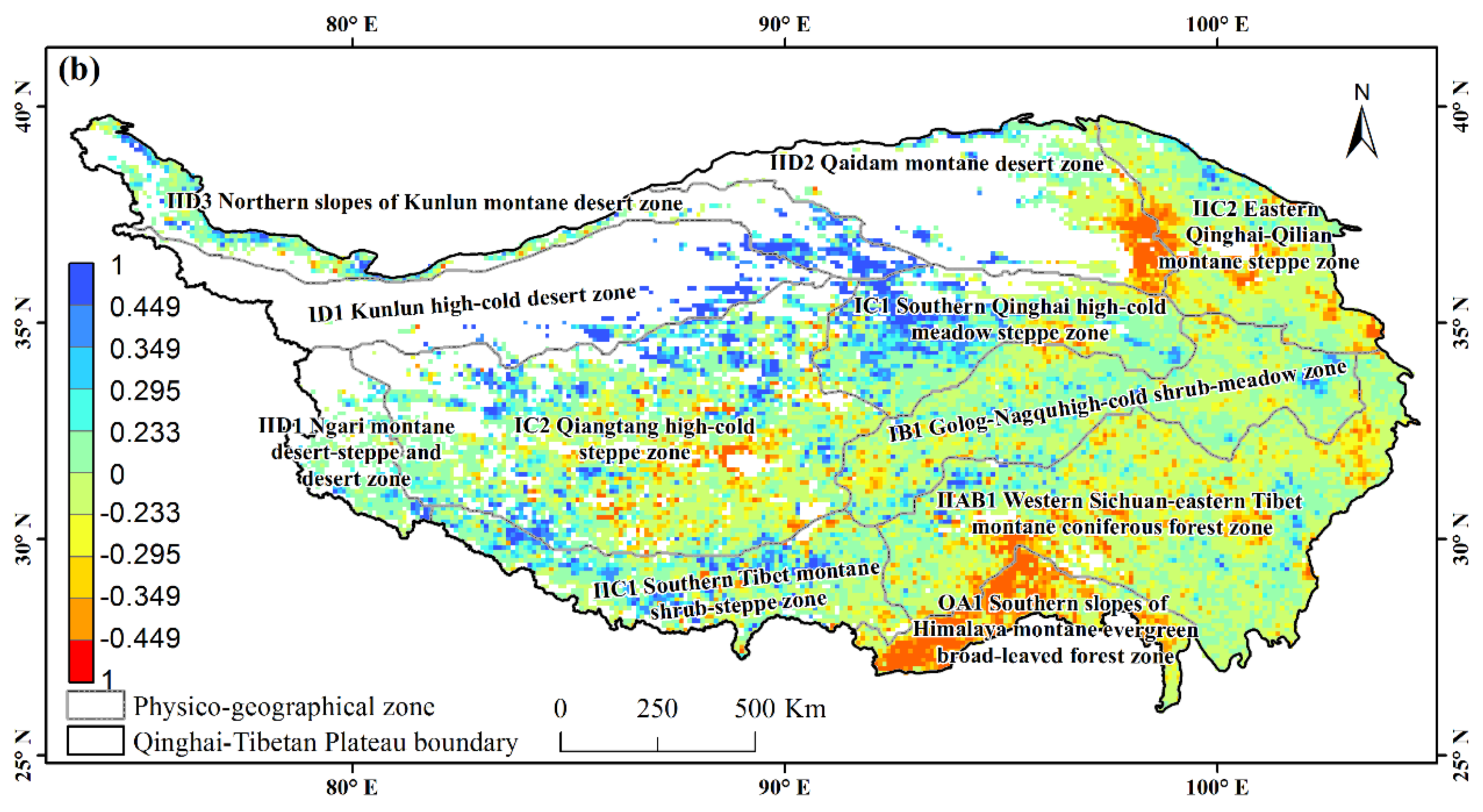
3.2. Partial Correlations between NDVI and Diurnal Extreme Temperature on the Tibetan Plateau
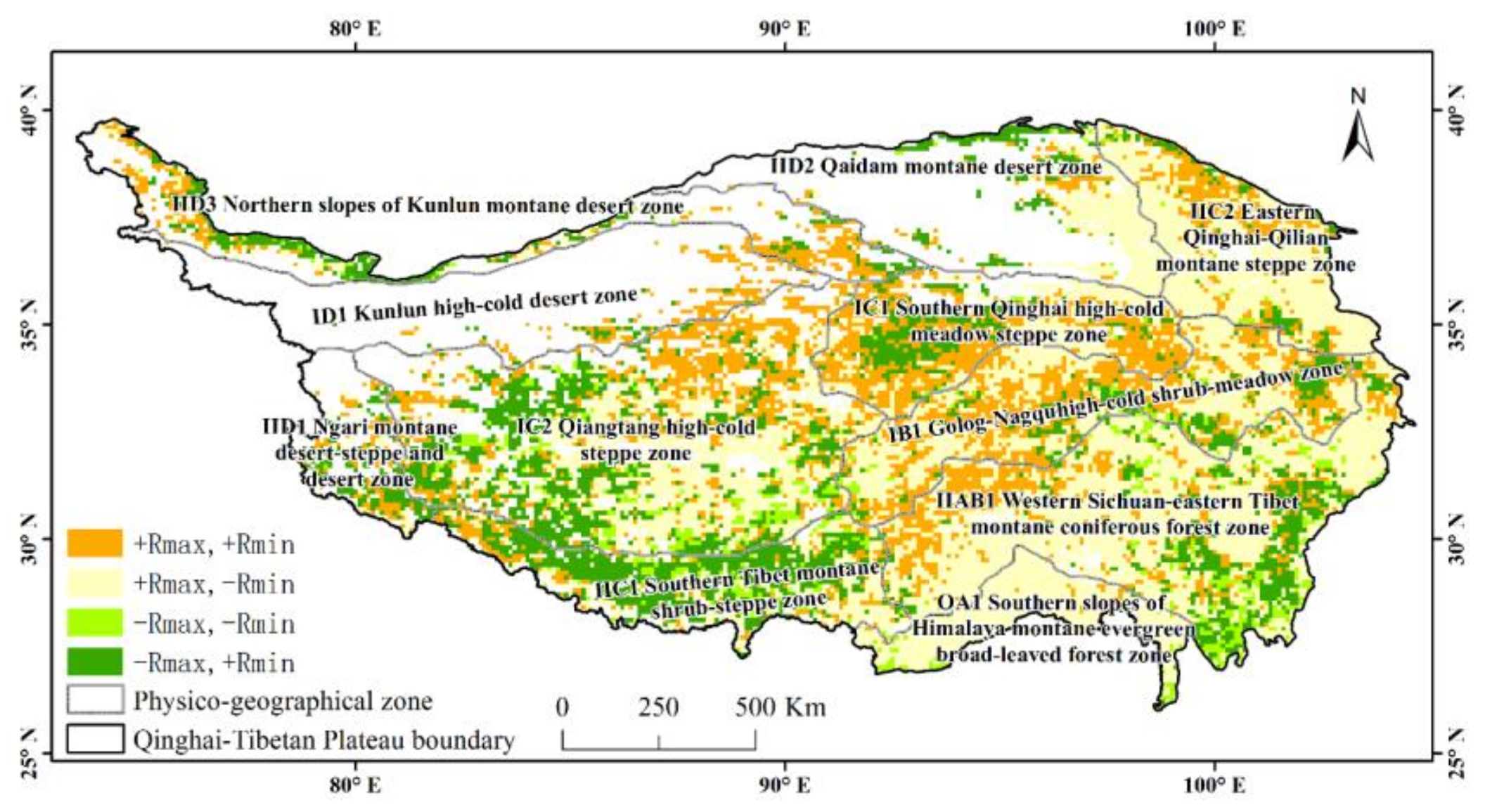
3.3. Partial Correlations between NDVI and Asymmetric Diurnal Warming in Different Physico-Geographical Regions
3.4. Partial Correlations between NDVI and Asymmetric Diurnal Warming among Different Vegetation Types
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

References
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Screen, J.A. Arctic amplification decreases temperature variance in northern mid- to high-latitudes. Nat. Clim. Chang. 2014, 4, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Piao, S.; Ciais, P.; Myneni, R.B.; Chen, A.; Chevallier, F.; Dolman, A.J.; Janssens, I.A.; Peñuelas, J.; Zhang, G. Asymmetric effects of daytime and night-time warming on Northern Hemisphere vegetation. Nature 2013, 501, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.D.; Drijber, R.; Smith, H.; Melillo, J. Microbial biomass, functional capacity, and community structure after 12 years of soil warming. Soil Biol. Biochem. 2008, 40, 2904–2907. [Google Scholar] [CrossRef]
- Walker, M.D.; Wahren, C.H.; Hollister, R.D.; Henry, G.H.; Ahlquist, L.E.; Alatalo, J.M.; Bret-Harte, M.S.; Calef, M.P.; Callaghan, T.V.; Carroll, A.B. Plant community responses to experimental warming across the tundra biome. Proc. Natl. Acad. Sci. USA 2006, 103, 1342–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.-X.; Yan, Y.; Hong, J.-T.; Lu, X.-Y.; Wang, X.-D. Impacts of warming on root biomass allocation in alpine steppe on the north Tibetan Plateau. J. Mt. Sci. 2017, 14, 1615–1623. [Google Scholar] [CrossRef]
- Zhang, G.; Kang, Y.; Han, G.; Sakurai, K. Effect of climate change over the past half century on the distribution, extent and NPP of ecosystems of Inner Mongolia. Glob. Chang. Biol. 2015, 17, 377–389. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Piao, S.; Chen, A.; Zeng, Z.; Ciais, P.; Janssens, I.A.; Mao, J.; Myneni, R.B.; Peng, S.; Peñuelas, J. Seasonally different response of photosynthetic activity to daytime and night-time warming in the Northern Hemisphere. Glob. Chang. Biol. 2015, 21, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Alward, R.D.; Detling, J.K.; Milchunas, D.G. Grassland vegetation changes and nocturnal global warming. Science 1999, 283, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9975. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. Atmos. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Pvv, P.; Pisipati, S.R.; Ristic, Z.; Bukovnik, U.; Fritz, A.K. Impact of Nighttime Temperature on Physiology and Growth of Spring Wheat. Crop Sci. 2008, 48, 2372–2380. [Google Scholar]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Sun, H.; Du, Z.; Yao, T.; Zhang, Y. Protection and Construction of the National Ecological Security Shelter Zone on Tibetan Plateau. Acta Ecol. Sin. 2012, 67, 3–12. [Google Scholar]
- Duan, A.; Wu, G.; Zhang, Q.; Liu, Q. The warming of the Qinghai-Tibet Plateau is new evidence of the intensification of greenhouse gas emissions. Chin. Sci. Bull. 2006, 51, 989–992. [Google Scholar] [CrossRef]
- Xu, H.J.; Wang, X.P.; Yang, T.B. Trend shifts in satellite-derived vegetation growth in Central Eurasia, 1982–2013. Sci. Total Environ. 2017, 579, 1658–1674. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Wang, X.; Ciais, P.; Zhu, B.; Wang, T.; Liu, J. Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Glob. Chang. Biol. 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- Buitenwerf, R.; Rose, L.; Higgins, S.I. Three decades of multi-dimensional change in global leaf phenology. Nat. Clim. Chang. 2015, 5, 364–368. [Google Scholar] [CrossRef]
- Pinzon, J.; Tucker, C. A Non-Stationary 1981–2012 AVHRR NDVI3g Time Series. Remote Sens. (Basel) 2014, 6, 6929–6960. [Google Scholar] [CrossRef]
- Xu, L.; Myneni, R.; Chapin, F., III; Callaghan, T.; Pinzon, J.; Tucker, C.; Zhu, Z.; Bi, J.; Ciais, P.; Tømmervik, H.; et al. Diminished temperature and vegetation seasonality over northern high latitudes. Nat. Clim. Chang. 2013, 1–7. [Google Scholar] [CrossRef]
- Piao, S.; Cui, M.; Chen, A.; Wang, X.; Ciais, P.; Liu, J.; Tang, Y. Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau. Agric. For. Meteorol. 2011, 151, 1599–1608. [Google Scholar] [CrossRef]
- Hutchinson, M.F. Interpolating mean rainfall using thin plate smoothing splines. Int. J. Geogr. Inf. Sci. 1995, 9, 385–403. [Google Scholar] [CrossRef]
- Yang, K.; Jie, H.; Tang, W.; Qin, J.; Cheng, C.C.K. On downward shortwave and longwave radiations over high altitude regions: Observation and modeling in the Tibetan Plateau. Agric. For. Meteorol. 2010, 150, 38–46. [Google Scholar] [CrossRef]
- Chinese Academy of Sciences. Vegetation Atlas of China; Science Press: Beijing, China, 2001. [Google Scholar]
- Zhang, Y.; Qi, W.; Zhou, C.; Ding, M.; Liu, L.; Gao, J.; Bai, W.; Wang, Z.; Zheng, D. Spatial and temporal variability in the net primary production (NPP) of alpine grassland on Tibetan Plateau from 1982 to 2009. Acta Ecol. Sin. 2013, 24, 1197–1211. [Google Scholar]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Luedeling, E.; Xu, J. Winter and spring warming result in delayed spring phenology on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2010, 107, 22151–22156. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, B. Climatic warming in the Tibetan Plateau during recent decades. Int. J. Climatol. 2000, 20, 1729–1742. [Google Scholar] [CrossRef]
- Jun, D.U.; Hongya, L.U.; Jian, J. Variations of extreme air temperature events over Tibet from 1961 to 2010. Acta Ecol. Sin. 2013, 68, 1269–1280. [Google Scholar]
- Ma, X.; Li, D. Analyses on air temperature and its abrupt change over Qinghai-Xizang Plateau in Modern age. Plateau Meteorol. 2003, 22, 507–512. [Google Scholar]
- Liang, C.; Yu, Q.; Liu, Y.; Zhang, Z. Effects of air temperature circadian on the NDVI of Nansi lake wetland vegetation. Trop. Geogr. 2015, 35, 422–426. [Google Scholar]
- Lucht, W.; Prentice, I.C.; Myneni, R.B.; Sitch, S.; Friedlingstein, P.; Cramer, W.; Bousquet, P.; Buermann, W.; Smith, B. Climatic control of the high-latitude vegetation greening trend and Pinatubo effect. Science 2002, 296, 1687–1689. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, M.H.; Murthy, R.; Griffin, K.L. The relative impacts of daytime and night-time warming on photosynthetic capacity in Populus deltoides. Plant Cell Environ. 2002, 25, 1729–1737. [Google Scholar] [CrossRef]
- Vicenteserrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; Lópezmoreno, J.I.; Azorínmolina, C.; Pasho, E.; Lorenzolacruz, J.; Revuelto, J. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.J.; Ho, C.H.; Kim, K.Y.; Jeong, J.H. Reduction of spring warming over East Asia associated with vegetation feedback. Geophys. Res. Lett. 2009, 36, 1–5. [Google Scholar] [CrossRef]
- Adams, H.D.; Guardiolaclaramonte, M.; Barrongafford, G.A.; Villegas, J.C.; Breshears, D.D.; Zou, C.B.; Troch, P.A.; Huxman, T.E. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. USA 2009, 106, 7063–7066. [Google Scholar] [CrossRef] [PubMed]
- Llorens, L.; Penuelas, J.; Filella, I. Diurnal and seasonal variations in the photosynthetic performance and water relations of two co-occurring Mediterranean shrubs, Erica multiflora and Globularia alypum. Physiol. Plant. 2003, 118, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Xia, J.; Liu, W.; Niu, S. Photosynthetic Overcompensation under Nocturnal Warming Enhances Grassland Carbon Sequestration. Ecology 2009, 90, 2700–2710. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, G. Response of Leymus Chinensis Grassland Vegetation in Inner Mongolia to Temperature change. Acta Phytoecol. Sin. 2004, 28, 507–514. [Google Scholar]
- Hu, T.; Kang, S. The compensatory effect in drought resistance of plants and its application in water-saving agriculture. Acta Ecol. Sin. 2005, 25, 885–891. [Google Scholar]
- Hu, H.C.; Wang, G.X.; Liu, G.S.; Li, T.B.; Ren, D.X.; Wang, Y.; Cheng, H.Y.; Wang, J.F. Influences of alpine ecosystem degradation on soil temperature in the freezing-thawing process on Qinghai-Tibet Plateau. Environ. Geol. 2009, 57, 1391–1397. [Google Scholar] [CrossRef]
- Yang, M.; Yao, T.; Gou, X.; Koike, T.; He, Y. The soil moisture distribution, thawing–freezing processes and their effects on the seasonal transition on the Qinghai–Xizang (Tibetan) plateau. J. Asian Earth Sci. 2003, 21, 457–465. [Google Scholar] [CrossRef]
- Peng, S.; Piao, S.; Ciais, P.; Fang, J.; Wang, X. Change in winter snow depth and its impacts on vegetation in China. Glob. Chang. Biol. 2010, 16, 3004–3013. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Tmax | Tmin |
|---|---|---|
| Forest | 0.598 *** | –0.561 *** |
| Shrublands | 0.446 ** | –0.337 * |
| Alpine Steppe | 0.132 | 0.263 |
| Alpine Meadow | 0.301 * | –0.39 |
| Crop | 0.455 *** | –0.428 ** |
| Others | 0.407 ** | 0.125 |
| Vegetation Types | Tmax | Tmin | ||||
|---|---|---|---|---|---|---|
| + | − | Total | + | − | Total | |
| Forest | 34.22 | 0.42 | 34.64 | 0.31 | 25.73 | 26.04 |
| Shrublands | 22.18 | 0.86 | 23.04 | 1.97 | 8.97 | 10.94 |
| Alpine Steppe | 11.06 | 1.62 | 12.68 | 14.01 | 5.83 | 19.86 |
| Alpine Meadow | 17.81 | 0.87 | 18.68 | 5.92 | 4.08 | 10.00 |
| Crop | 41.46 | 0 | 41.46 | 0 | 39.02 | 39.02 |
| Others | 17.75 | 1.04 | 18.79 | 12.25 | 8.24 | 20.49 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, H.; Li, A.; Feng, G.; Li, Y.; Qin, Y.; Lei, G.; Cui, Y. The Effects of Asymmetric Diurnal Warming on Vegetation Growth of the Tibetan Plateau over the Past Three Decades. Sustainability 2018, 10, 1103. https://doi.org/10.3390/su10041103
Xia H, Li A, Feng G, Li Y, Qin Y, Lei G, Cui Y. The Effects of Asymmetric Diurnal Warming on Vegetation Growth of the Tibetan Plateau over the Past Three Decades. Sustainability. 2018; 10(4):1103. https://doi.org/10.3390/su10041103
Chicago/Turabian StyleXia, Haoming, Ainong Li, Gary Feng, Yang Li, Yaochen Qin, Guangbin Lei, and Yaoping Cui. 2018. "The Effects of Asymmetric Diurnal Warming on Vegetation Growth of the Tibetan Plateau over the Past Three Decades" Sustainability 10, no. 4: 1103. https://doi.org/10.3390/su10041103