
Decreased Expression of KIFC1 in Human Testes with Globozoospermic Defects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. RNA Extraction and Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
2.3. SDS-PAGE and Immunoblot Analysis
2.4. Immunohistochemistry and Immunocytochemistry
2.5. Kifc1 Gene Knockdown and Evaluation of Epididymal Sperm Morphology
2.6. Hematoxylin and Eosin Staining
2.7. Statistical Analysis
2.8. Ethics Statement
3. Results
3.1. Clinical Characteristics of Globozoospermia and Obstructive Azoospermia
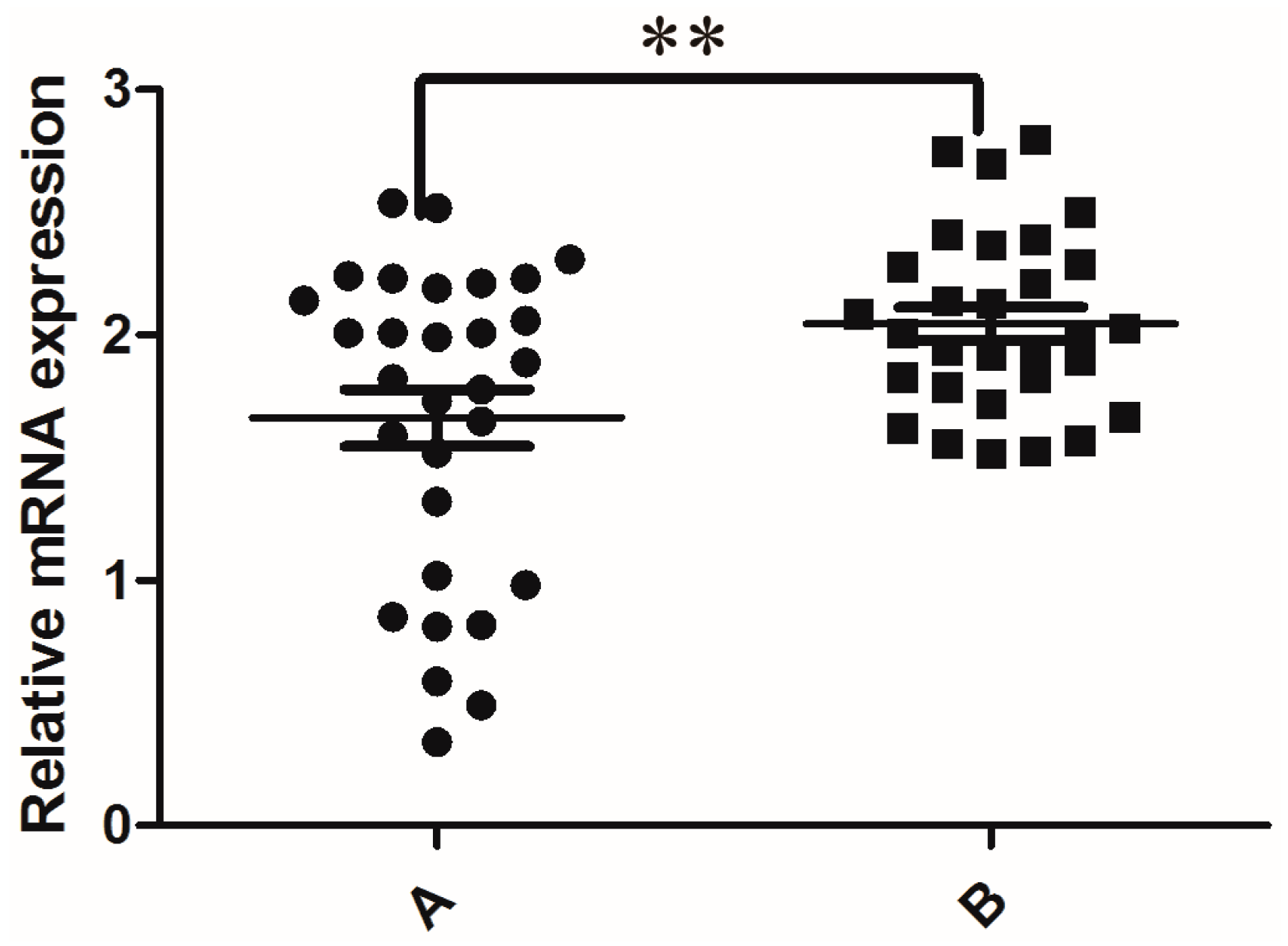
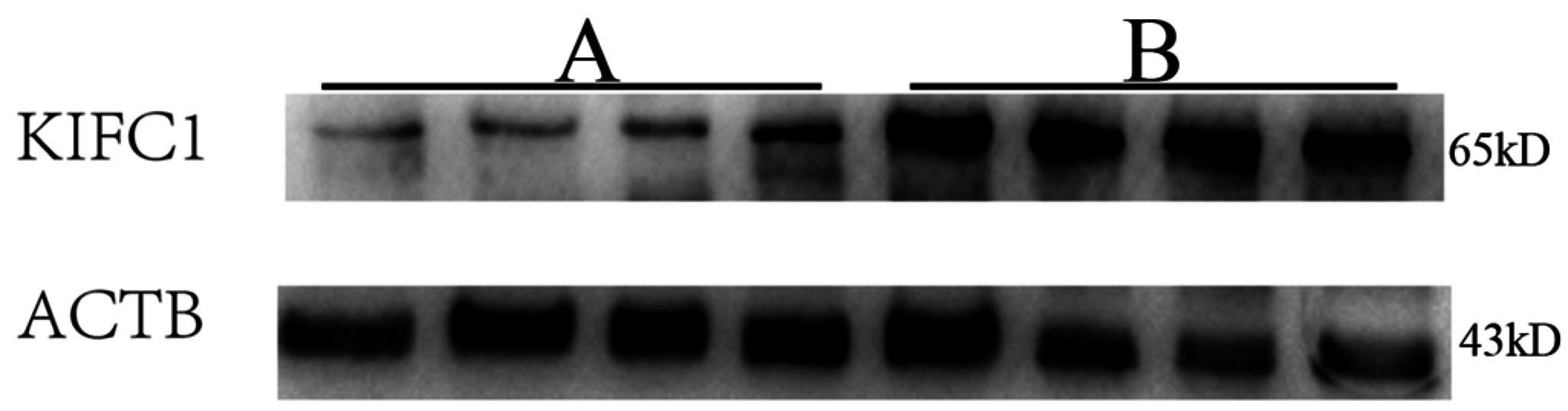
3.2. KIFC1 Expression in the Testes of Patients with Globozoospermia and Obstructive Azoospermia
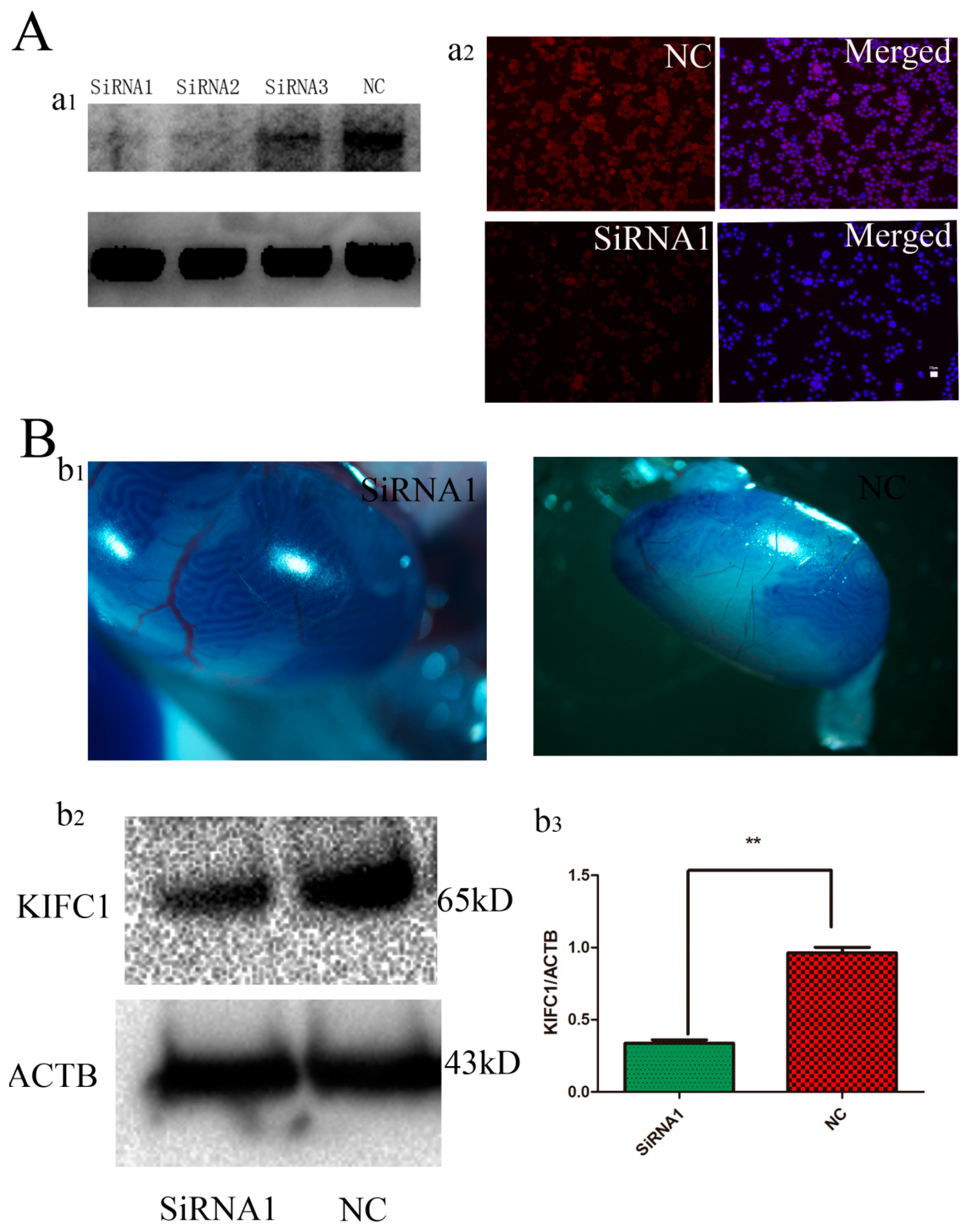
3.3. Kifc1 Knockdown Increased the Rate of Globozoospermic Sperm
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of interest
References
- Ghedir, H.; Ibala-Romdhane, S.; Okutman, O.; Viot, G.; Saad, A.; Viville, S. Identification of a new DPY19L2 mutation and a better definition of DPY19L2 deletion breakpoints leading to globozoospermia. Mol. Hum. Reprod. 2016, 22, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Karaca, N.; Yilmaz, R.; Kanten, G.E.; Kervancioglu, E.; Solakoglu, S.; Kervancioglu, M.E. First successful pregnancy in a globozoospermic patient having homozygous mutation in SPATA16. Fertil. Steril. 2014, 102, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Doran, J.; Walters, C.; Kyle, V.; Wooding, P.; Hammett-Burke, R.; Colledge, W.H. Mfsd14a (Hiat1) gene disruption causes globozoospermia and infertility in male mice. Reproduction 2016, 152, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wan, H.; Li, X.; Liu, W.; Chen, Q.; Wang, Y.; Yang, L.; Tang, H.; Zhang, X.; Duan, E.; et al. Atg7 is required for acrosome biogenesis during spermatogenesis in mice. Cell Res. 2014, 24, 852–869. [Google Scholar] [CrossRef] [PubMed]
- Funaki, T.; Kon, S.; Tanabe, K.; Natsume, W.; Sato, S.; Shimizu, T.; Yoshida, N.; Wong, W.F.; Ogura, A.; Ogawa, T.; et al. The Arf GAP SMAP2 is necessary for organized vesicle budding from the trans-Golgi network and subsequent acrosome formation in spermiogenesis. Mol. Biol. Cell 2013, 24, 2633–2644. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, Y.; Satouh, Y.; Inoue, N.; Isotani, A.; Ikawa, M.; Okabe, M. Spaca1-deficient male mice are infertile with abnormally shaped sperm heads reminiscent of globozoospermia. Development 2012, 139, 3583–3589. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.Y.; Li, N.; Li, T.F.; Li, W.W.; Zhang, C.; Shao, Y.; Cui, Y.X.; Yin, Z.M.; Xia, X.Y.; Li, Z.R. Detection of DPY19L2 gene mutation in a globozoospermia patient. Natl. J. Androl. 2013, 19, 1011–1015. (In Chinese) [Google Scholar]
- Jockusch, H.; Holland, A.; Staunton, L.; Schmitt-John, T.; Heimann, P.; Dowling, P.; Ohlendieck, K. Pathoproteomics of testicular tissue deficient in the garp component Vps54: The wobbler mouse model of globozoospermia. Proteomics 2014, 14, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Paiardi, C.; Pasini, M.E.; Gioria, M.; Berruti, G. Failure of acrosome formation and globozoospermia in the wobbler mouse, a Vps54 spontaneous recessive mutant. Spermatogenesis 2011, 1, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Shi, Q.W.; Lu, G.X. A newly discovered mutation in pick1 in a human with globozoospermia. Asian J. Androl. 2010, 12, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Kam, C.; Shen, C.; Jin, W.; Wang, J.; Lee, K.M.; Jiang, L.; Xia, J. Pick1 deficiency causes male infertility in mice by disrupting acrosome formation. J. Clin. Investig. 2009, 119, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Kang-Decker, N.; Mantchev, G.T.; Juneja, S.C.; McNiven, M.A.; van Deursen, J.M. Lack of acrosome formation in Hrb-deficient mice. Science 2001, 294, 1531–1533. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Toselli, P.A.; Russell, L.D.; Seldin, D.C. Globozoospermia in mice lacking the casein kinase II alpha’ catalytic subunit. Nat. Genet. 1999, 23, 118–121. [Google Scholar] [PubMed]
- Dam, A.H.; Koscinski, I.; Kremer, J.A.; Moutou, C.; Jaeger, A.S.; Oudakker, A.R.; Tournaye, H.; Charlet, N.; Lagier-Tourenne, C.; van Bokhoven, H.; et al. Homozygous mutation in Spata16 is associated with male infertility in human globozoospermia. Am. J. Hum. Genet. 2007, 81, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Harbuz, R.; Zouari, R.; Pierre, V.; Ben Khelifa, M.; Kharouf, M.; Coutton, C.; Merdassi, G.; Abada, F.; Escoffier, J.; Nikas, Y.; et al. A recurrent deletion of DPY19L2 causes infertility in man by blocking sperm head elongation and acrosome formation. Am. J. Hum. Genet. 2011, 88, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Koscinski, I.; Elinati, E.; Fossard, C.; Redin, C.; Muller, J.; Velez de la Calle, J.; Schmitt, F.; Ben Khelifa, M.; Ray, P.F.; Kilani, Z.; et al. DPY19L2 deletion as a major cause of globozoospermia. Am. J. Hum. Genet. 2011, 88, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Saito, N.; Okada, Y.; Noda, Y.; Kinoshita, Y.; Kondo, S.; Hirokawa, N. KIFC2 is a novel neuron-specific C-terminal type kinesin superfamily motor for dendritic transport of multivesicular body-like organelles. Neuron 1997, 18, 425–438. [Google Scholar] [CrossRef]
- Wang, D.H.; Yang, W.X. Molecular cloning and characterization of KIFC1-like kinesin gene (es-KIFC1) in the testis of the chinese mitten crab Eriocheir sinensis. Comp. Biochem. Physiol. A 2010, 157, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, J.Q.; Yang, W.X. Molecular cloning and characterization of KIFC1-like kinesin gene (ot-KIFC1) from Octopus tankahkeei. Comp. Biochem. Physiol. B 2010, 156, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.T.; Mao, H.; Hou, C.C.; Sun, X.; Wang, D.H.; Zhou, H.; Yang, W.X. Characterization and expression pattern of KIFC1-like kinesin gene in the testis of the macrobrachium nipponense with discussion of its relationship with structure lamellar complex (LCx) and acroframosome (AFS). Mol. Biol. Rep. 2012, 39, 7591–7598. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.X.; Sperry, A.O. C-terminal kinesin motor KIFC1 participates in acrosome biogenesis and vesicle transport. Biol. Reprod. 2003, 69, 1719–1729. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, C.; Yang, S.; Tian, R.; Wang, J.; Yuan, Q.; Dong, H.; He, Z.; Wang, S.; Li, Z. Dynamics of the transcriptome during human spermatogenesis: Predicting the potential key genes regulating male gametes generation. Sci. Rep. 2016, 6, 19069. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Hamada, A.; Esteves, S.C. Engaging practicing gynecologists in the management of infertile men. J. Obstet. Gynaecol. India 2015, 65, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Ikawa, M.; Tergaonkar, V.; Ogura, A.; Ogonuki, N.; Inoue, K.; Verma, I.M. Restoration of spermatogenesis by lentiviral gene transfer: Offspring from infertile mice. Proc. Natl. Acad. Sci. USA 2002, 99, 7524–7529. [Google Scholar] [CrossRef] [PubMed]
- Fatima, R. Drosophila Dynein intermediate chain gene, Dic61B, is required for spermatogenesis. PLoS ONE 2011, 6, e27822. [Google Scholar] [CrossRef] [PubMed]
- Dam, A.H.; Pijnenburg, A.J.; Hendriks, J.C.; Westphal, H.; Ramos, L.; Kremer, J.A. Intracytoplasmic sperm injection in partial globozoospermia. Fertil. Steril. 2012, 97, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Alvarez Sedo, C.; Rawe, V.Y.; Chemes, H.E. Acrosomal biogenesis in human globozoospermia: Immunocytochemical, ultrastructural and proteomic studies. Hum. Reprod. 2012, 27, 1912–1921. [Google Scholar] [CrossRef] [PubMed]
- Yassine, S.; Escoffier, J.; Abi Nahed, R.; Pierre, V.; Karaouzene, T.; Ray, P.F.; Arnoult, C. Dynamics of Sun5 localization during spermatogenesis in wild type and DPY19L2 knock-out mice indicates that Sun5 is not involved in acrosome attachment to the nuclear envelope. PLoS ONE 2015, 10, e0118698. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.X.; Jefferson, H.; Sperry, A.O. The molecular motor KIFC1 associates with a complex containing nucleoporin NUP62 that is regulated during development and by the small GTPase RAN. Biol. Reprod. 2006, 74, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Perrin, A.; Coat, C.; Nguyen, M.H.; Talagas, M.; Morel, F.; Amice, J.; De Braekeleer, M. Molecular cytogenetic and genetic aspects of globozoospermia: A review. Andrologia 2013, 45, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.; Bananis, E.; Sarkar, S.; Stockert, R.J.; Sperry, A.O.; Murray, J.W.; Wolkoff, A.W. KIF5B and KIFC1 interact and are required for motility and fission of early endocytic vesicles in mouse liver. Mol. Biol. Cell 2007, 18, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; An, L.M.; Xia, X.Y. Molecular genetics of globozoospermia: An update. Natl. J. Androl. 2011, 17, 935–938. (In Chinese) [Google Scholar]
- Thornhill, J.A.; Fanning, D.M.; Davis, N.F.; Ward, F.; Shamoun, O.; Brinsden, P. Testicular sperm extraction and intracytoplasmic sperm injection: Outcomes in a specialist fertility centre. Ir. Med. J. 2015, 108, 263–265. [Google Scholar] [PubMed]
- Wu, Y.; Chen, X.; Wang, S.; Jiang, M.; Zheng, B.; Zhou, Q.; Bi, Y.; Zhou, Z.; Huang, X.; Sha, J. Flotillin-2 is an acrosome-related protein involved in mouse spermiogenesis. J. Biomed. Res. 2012, 26, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, M.; Sobczak, A.; Weitzel, J.M. In vivo microinjection and electroporation of mouse testis. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.D.; Moreira, A.C.; Sa, R.; Monteiro, M.P.; Sousa, M.; Carvalho, R.A.; Silva, B.M.; Oliveira, P.F.; Alves, M.G. Leptin modulates human Sertoli cells acetate production and glycolytic profile: A novel mechanism of obesity-induced male infertility? Biochim. Biophys. Acta 2015, 1852, 1824–1832. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhi, E.; Li, P.; Chen, H.; Xu, P.; Zhu, X.; Zhu, Z.; He, Z.; Li, Z. Decreased Expression of KIFC1 in Human Testes with Globozoospermic Defects. Genes 2016, 7, 75. https://doi.org/10.3390/genes7100075
Zhi E, Li P, Chen H, Xu P, Zhu X, Zhu Z, He Z, Li Z. Decreased Expression of KIFC1 in Human Testes with Globozoospermic Defects. Genes. 2016; 7(10):75. https://doi.org/10.3390/genes7100075
Chicago/Turabian StyleZhi, Erlei, Peng Li, Huixing Chen, Peng Xu, Xiaobin Zhu, Zijue Zhu, Zuping He, and Zheng Li. 2016. "Decreased Expression of KIFC1 in Human Testes with Globozoospermic Defects" Genes 7, no. 10: 75. https://doi.org/10.3390/genes7100075