
Change of Soil Carbon Fractions and Water-Stable Aggregates in a Forest Ecosystem Succession in South China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Coordinate | Forest Age (Years) | Community Biomass (Mg ha−1) |
|---|---|---|---|
| PF | 23°09′ N, 112°33′ E | 60 | 122.5 |
| PBMF | 23°10′ N, 112°32′ E | 110 | 164.1 |
| MEBF | 23°10′ N, 112°32′ E | 400 | 290.4 |
2.2. Soil Sampling and Treatments
2.3. Laboratory Analyses and Calculations
2.3.1. SOC Fractions
2.3.2. POM-C and POM-N from the Whole Soil
2.3.3. Water-Stable Aggregate Fractionation
2.3.4. SOC Concentration from the Bulk Soil and Each Aggregate Size Class
2.4. Statistical Analysis
3. Results
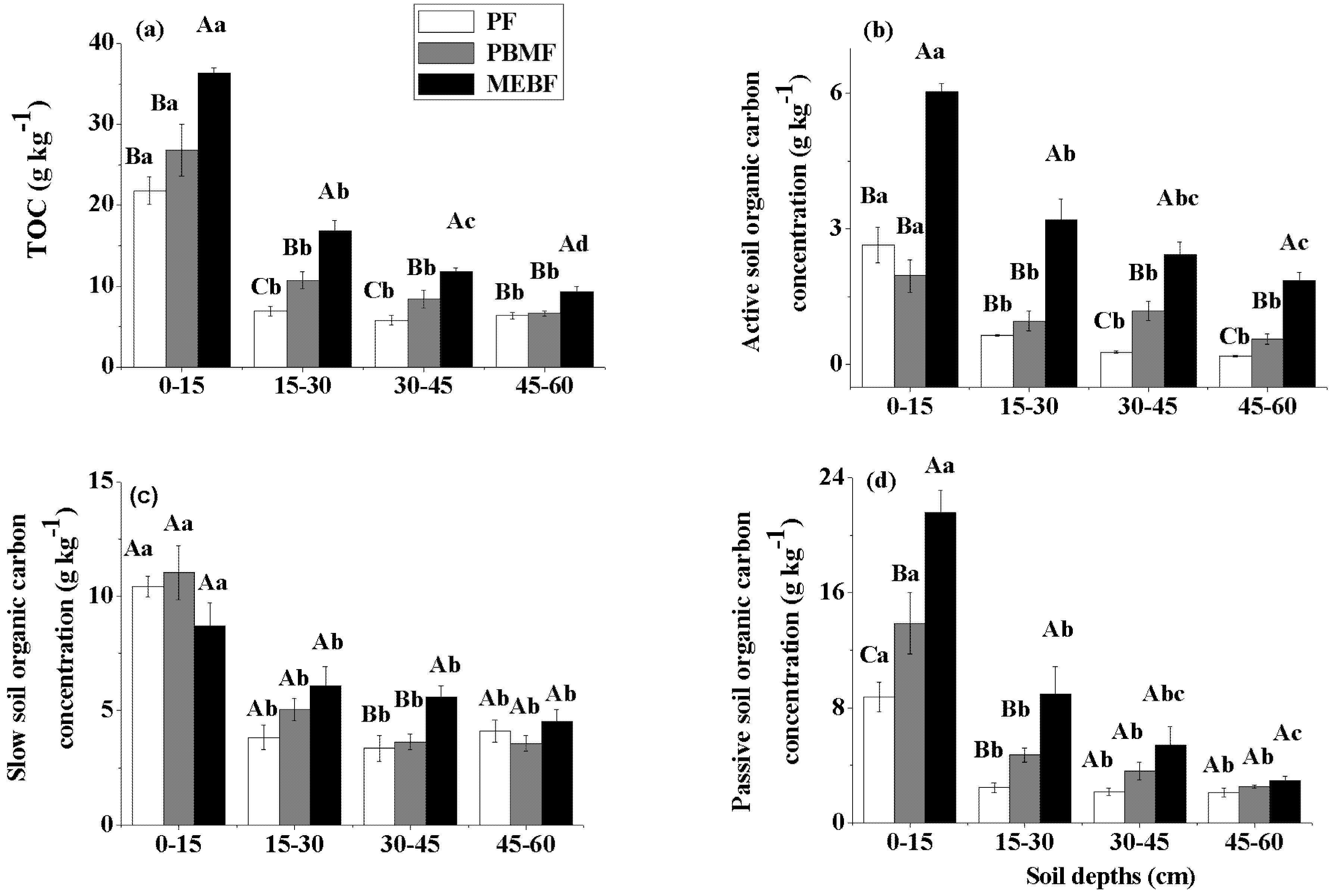
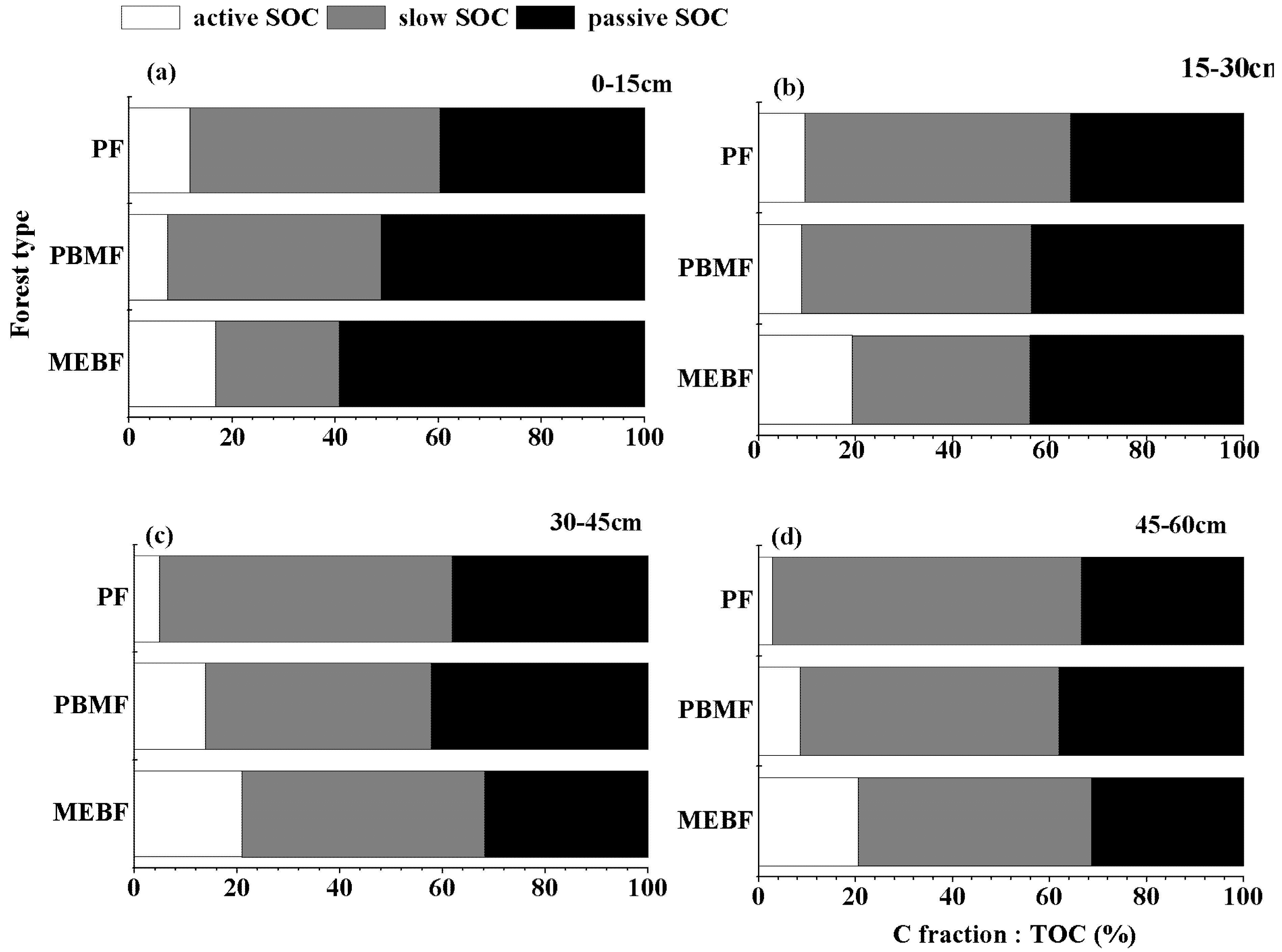
3.1. Organic Carbon Concentration in Soil Fractions
| TOC | Active SOC | Slow SOC | Passive SOC | POM-C | POM-N | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | F | P | F | P | |
| Forest type | 44.7 | 0.00 | 118.1 | 0.00 | 1.5 | 0.24 | 29.2 | 0.00 | 52.7 | 0.00 | 64.9 | 0.00 |
| Soil depth | 176.3 | 0.00 | 68.0 | 0.00 | 56.8 | 0.00 | 77.4 | 0.00 | 45.2 | 0.00 | 52.3 | 0.00 |
| Forest type × Soil depth | 4.0 | 0.00 | 7.3 | 0.00 | 3.0 | 0.02 | 5.8 | 0.00 | 11.4 | 0.00 | 14.7 | 0.00 |


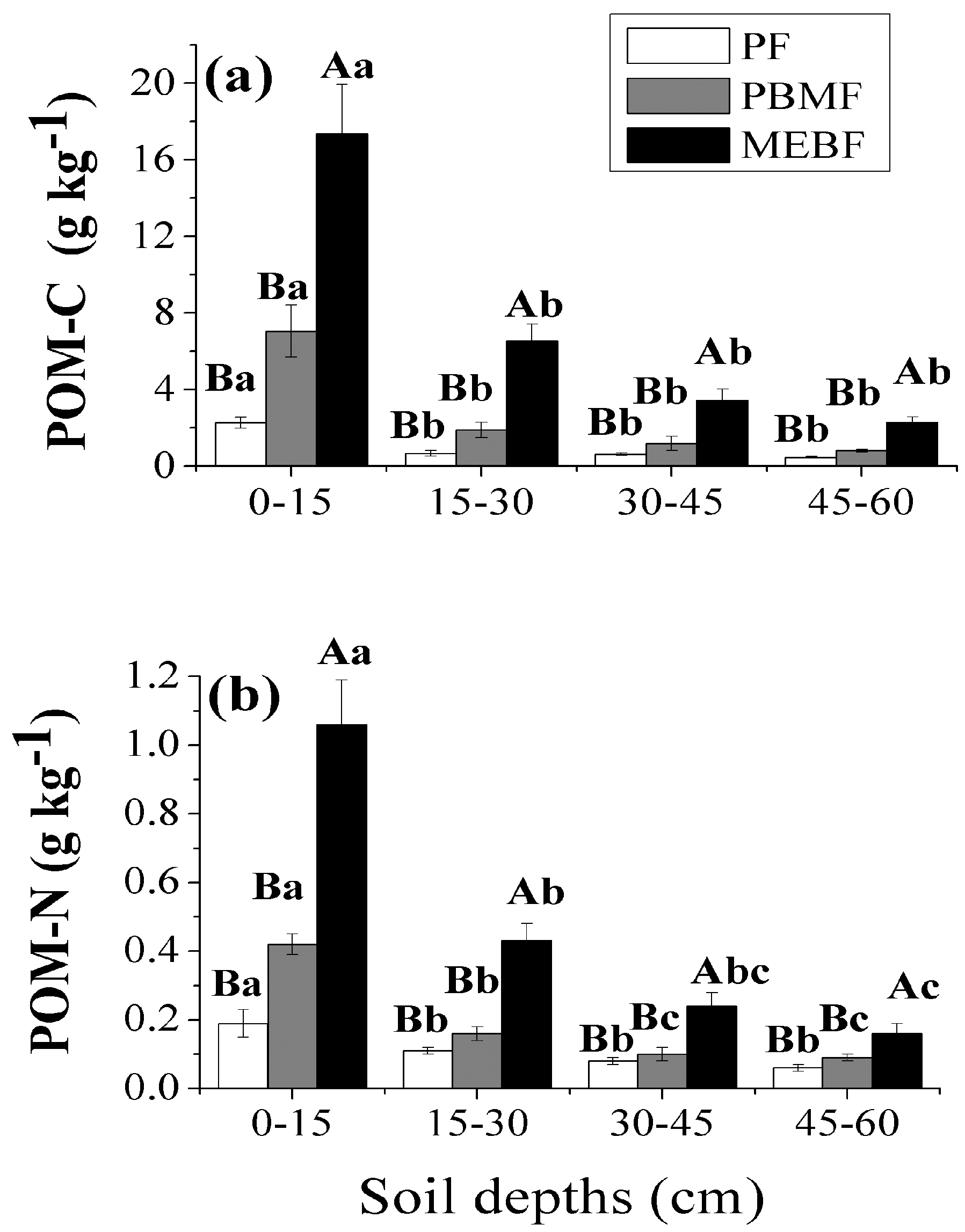
3.2. POM-C and POM-N from the Whole Soil

3.3. The Proportion of Water-Stable Aggregates in Soil
| >2000 μm | 250–2000 μm | 53–250 μm | <53 μm | |||||
|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | |
| Forest type | 18.9 | 0.00 | 3.4 | 0.04 | 0.1 | 0.90 | 2.1 | 0.13 |
| Soil depths | 57.4 | 0.00 | 2.5 | 0.07 | 8.7 | 0.00 | 14.9 | 0.00 |
| Forest type × Soil depths | 4.0 | 0.00 | 0.9 | 0.50 | 0.9 | 0.53 | 1.6 | 0.16 |
| Soil Depth (cm) | Forest Type | % of Dry Soil Weight of Different Size Class Aggregates | |||
|---|---|---|---|---|---|
| >2000 μm | 250–2000 μm | 53–250 μm | <53 μm | ||
| 0–15 | PF | 18.2 ± 4.7 B | 66.8 ± 3.4 A | 11.1 ± 1.6 A | 4.0 ± 0.6 A |
| PBMF | 26.2 ± 4.1 B | 62.9 ± 5.3 A | 6.7 ± 1.2 A | 4.3 ± 0.4 A | |
| MEBF | 40.7 ± 4.6 A | 46.3 ± 6.6 B | 6.6 ± 1.4 A | 6.4 ± 3.3 A | |
| 15–30 | PF | 7.4 ± 2.2 B | 75.1 ± 2.5 A | 14.3 ± 3.3 A | 3.3 ± 0.6 A |
| PBMF | 4.9 ± 1.9 B | 72.3 ± 4.3 A | 14.2 ± 1.0 A | 8.6 ± 3.2 A | |
| MEBF | 14.5 ± 1.9 A | 64.2 ± 4.3 A | 15.1 ± 3.3 A | 6.2 ± 1.1 A | |
| 30–45 | PF | 3.9 ± 1.4 A | 68.1 ± 6.3 A | 21.5 ± 6.0 A | 6.5 ± 2.7 A |
| PBMF | 3.0 ± 1.1 A | 67.9 ± 3.4 A | 21.8 ± 2.7 A | 7.30 ± 0.8 A | |
| MEBF | 10.0 ± 3.5 A | 68.3 ± 4.1 A | 14.6 ± 1.1 A | 7.1 ± 1.2 A | |
| 45–60 | PF | 2.5 ± 0.9 A | 75.6 ± 4.8 A | 17.7 ± 4.0 A | 4.2 ± 0.9 B |
| PBMF | 1.7 ± 0.5 A | 69.1 ± 3.1 A | 21.9 ± 3.5 A | 7.3 ± 0.2 AB | |
| MEBF | 4.5 ± 1.8 A | 63.4 ± 4.1 A | 23.4 ± 3.8 A | 8.7 ± 1.6 A | |
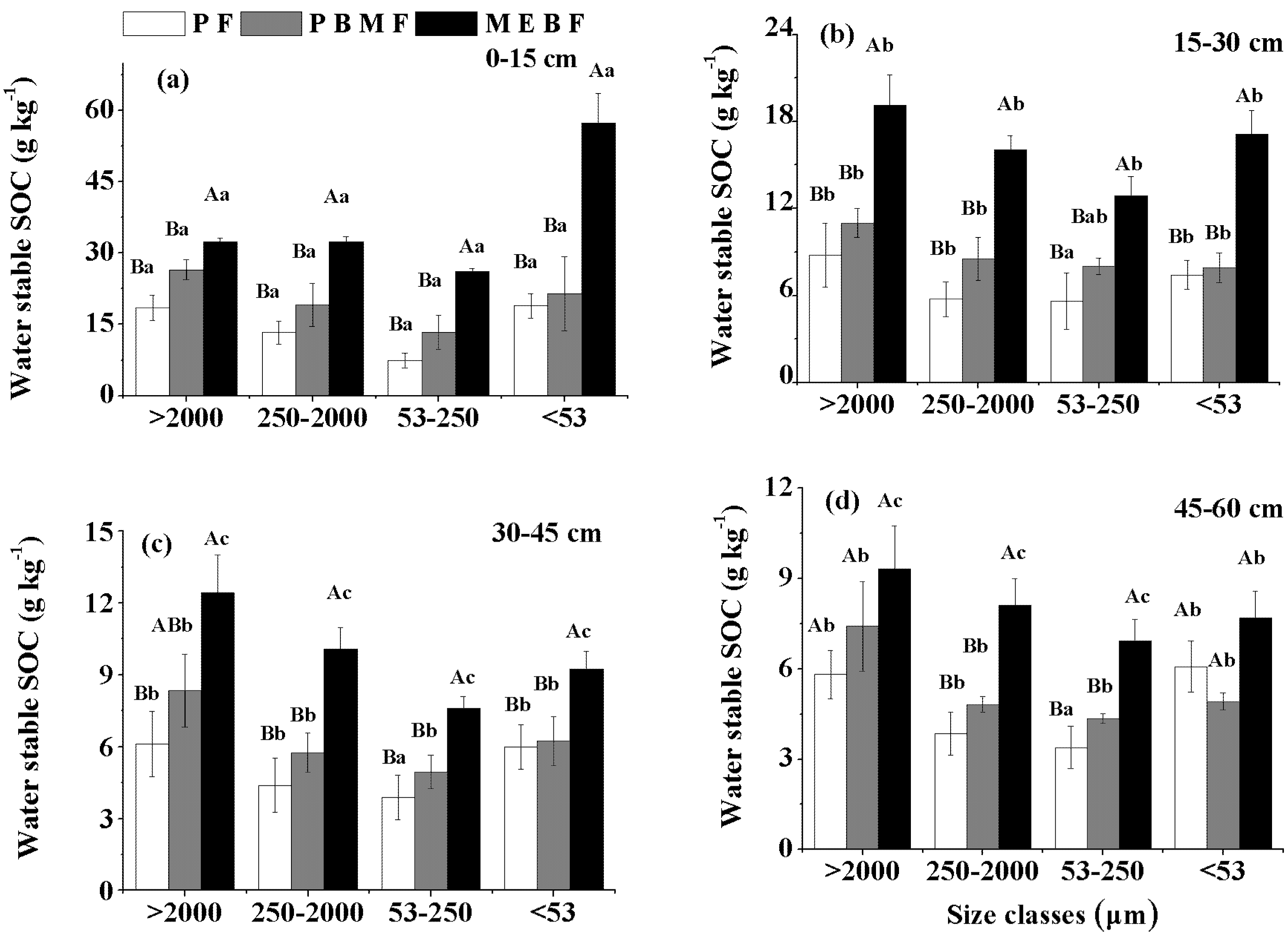
3.4. Sand-Free SOC Concentration in Water-Stable Aggregates
| >2000 μm | 250–2000 μm | 53–250 μm | <53 μm | |||||
|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | |
| Forest type | 14.1 | 0.00 | 34.1 | 0.00 | 36.1 | 0.00 | 23.9 | 0.00 |
| Soil depths | 51.9 | 0.00 | 53.3 | 0.00 | 36.3 | 0.00 | 45.3 | 0.00 |
| Forest type × Soil depths | 1.3 | 0.28 | 3.9 | 0.00 | 6.6 | 0.00 | 9.9 | 0.00 |

3.5. Relationship between SOC Fractions and Water-Stable Aggregation
| Water-Stable Aggregate Size (μm) | Active SOC | Slow SOC | Passive SOC | POM-C | POM-N | ||||
|---|---|---|---|---|---|---|---|---|---|
| >2000 | 250–2000 | 53–250 | <53 | ||||||
| TOC | 0.892 ** | −0.551 ** | −0.561 | −0.046 | 0.836 ** | 0.820 ** | 0.954 ** | 0.868 ** | 0.851 ** |
| Active SOC | 0.760 ** | −0.572 ** | −0.370 ** | −0.035 | 0.532 ** | 0.824 ** | 0.820 ** | 0.831 ** | |
| Slow SOC | 0.667 ** | −0.251 | −0.566 ** | −0.191 | 0.681 ** | 0.507 ** | 0.488 ** | ||
| Passive SOC | 0.914 ** | −0.630 ** | −0.516 ** | 0.019 | 0.909 ** | 0.888 ** | |||
| POM-C | 0.845 ** | −0.625 ** | −0.471 ** | 0.130 | 0.981 ** | ||||
| POM-N | 0.849 ** | −0.608 ** | −0.487 ** | 0.102 | |||||
4. Discussion
4.1. Change of Various C Fractions along Forest Succession Gradient
4.2. Change of Soil Aggregation and Organic C Protection along Forest Succession Gradient
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Parton, W.J.; Schimel, D.S.; Cole, C.V.; Ojima, D.S. Analysis of factors controlling soil organic matter levels in great plains grasslands. Soil Sci. Soc. Am. J. 1987, 51, 1173–1179. [Google Scholar] [CrossRef]
- Chen, X.M.; Liu, J.X.; Deng, Q.; Yan, J.H.; Zhang, D.Q. Effects of elevated CO2 and nitrogen addition on soil organic carbon fractions in a subtropical forest. Plant Soil 2012, 357, 25–34. [Google Scholar] [CrossRef]
- Guenet, B.; Sabrina, J.; Bardoux, G.; Abbadie, L.; Chenu, C. Evidence that stable C is as vulnerable to priming effect as is more labile C in soil. Soil Biol. Biochem. 2012, 52, 43–48. [Google Scholar] [CrossRef]
- Amezketa, E. Soil aggregate stability: A review. J. Sustain. Agric. 1999, 14, 83–151. [Google Scholar] [CrossRef]
- Bronick, C.J.; Lal, R. Soil structure and management: A review. Geoderma 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Elliott, E.T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci. Soc. Am. J. 1986, 50, 627–633. [Google Scholar] [CrossRef]
- Chaney, K.; Swift, R.S. The influence of organic matter on aggregate stability in some British soils. J. Soil Sci. 1984, 35, 223–230. [Google Scholar] [CrossRef]
- Oades, J.M. Soil organic matter and structural stability mechanisms and implications for management. Plant Soil 1984, 76, 319–337. [Google Scholar] [CrossRef]
- Angers, D.A. Changes in soil aggregation and organic carbon under corn and alfalfa. Soil Sci. Soc. Am. J. 1992, 56, 1244–1249. [Google Scholar] [CrossRef]
- Besnard, E.; Chenu, C.; Balesdent, J.; Puget, P.; Arrouaya, D. Fate of particulate organic matter in soil aggregates during cultivation. Eur. J. Soil Sci. 1996, 47, 495–503. [Google Scholar] [CrossRef]
- Veum, K.S.; Goyne, K.W.; Kremer, R.; Motavalli, P.P. Relationships among water stable aggregates and organic matter fractions under conservation management. Soil Sci. Soc. Am. J. 2012, 76, 2143–2153. [Google Scholar] [CrossRef]
- Zhao, H.; Lv, Y.Z.; Wang, X.K.; Zhang, H.L.; Yang, X.M. Tillage impacts on the fractions and compositions of soil organic carbon. Geoderma 2012, 189, 397–403. [Google Scholar] [CrossRef]
- Andruschkewitsch, R.; Geisseler, D.; Koch, H.J.; Ludwig, B. Effects of tillage on contents of organic carbon, nitrogen, water-stable aggregates and light fraction for four different long-term trials. Geoderma 2013, 192, 368–377. [Google Scholar] [CrossRef]
- Linsler, D.; Geisseler, D.; Loges, R.; Taube, F.; Ludwig, B. Temporal dynamics of soil organic matter composition and aggregate distribution in permanent grassland after a single tillage event in a temperate climate. Soil Tillage Res. 2013, 126, 90–99. [Google Scholar] [CrossRef]
- Plaza-Bonilla, D.; Cantero-Martínez, C.; Vinas, P.; Alvaro-Fuentes, J. Soil aggregation and organic carbon protection in a no-tillage chronosequence under Mediterranean conditions. Geoderma 2013, 193–194, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, J.S. Microbial biomass associated with water-stable aggregates in forest, savanna and cropland soils of a seasonally dry tropical region, India. Soil Biol. Biochem. 1995, 27, 1027–1033. [Google Scholar] [CrossRef]
- Singh, S.; Singh, J.S. Water-stable aggregates and associated organic matter in forest, savanna and cropland soils of a seasonally dry tropical region, India. Biol. Fertil. Soils 1996, 22, 76–82. [Google Scholar] [CrossRef]
- Spaccini, R.; Zena, A.; Igwe, C.A.; Mbagwu, J.S.C.; Piccolo, A. Carbohydrates in water-stable aggregates and particle size fractions of forested and cultivated soils in two contrasting tropical ecosystems. Biogeochemistry 2001, 53, 1–22. [Google Scholar] [CrossRef]
- Caravaca, F.; Lax, A.; Albaladejo, J. Aggregate stability and carbon characteristics of particle-size fractions in cultivated and forested soils of semiarid Spain. Soil Tillage Res. 2004, 78, 83–90. [Google Scholar] [CrossRef]
- An, S.S.; Mentler, A.; Mayer, H.; Blum, W.E.H. Soil aggregation, aggregate stability, organic carbon and nitrogen in different soil aggregate fractions under forest and shrub vegetation on the Loess Plateau, China. Catena 2010, 81, 226–233. [Google Scholar] [CrossRef]
- Buol, S.W.; Southard, R.J.; Graham, R.C.; McDaniel, P.A. Soil Genesis and Classification, 5th ed.; Iowa State University Press: Iowa, IA, USA, 2003; pp. 339–347. [Google Scholar]
- Wang, B.S.; Ma, M.J. The successions of the forest community in Dinghushan. Trop. Subtrop. For. Ecosyst. Res. 1982, 1, 142–156. (In Chinese) [Google Scholar]
- Peng, S.L.; Wang, B.S. Forest succession at Dinghushan, Guangdong, China. J. Trop. Subtrop. Bot. 1992, 7, 75–80. (In Chinese) [Google Scholar]
- Kong, G.H.; Huang, Z.L.; Zhang, Q.M.; Liu, S.Z.; Mo, J.M.; He, D.Q. Type, structure, dynamics and management of the lower subtropical evergreen broad-leaved forest in the Dinghushan Biosphere Reserve of China. Tropics 1997, 6, 335–350. [Google Scholar] [CrossRef]
- Tang, X.L.; Liu, S.G.; Zhou, G.Y.; Zhang, D.Q.; Zhou, C.Y. Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China. Global Chang. Biol. 2006, 12, 546–560. [Google Scholar] [CrossRef]
- Yan, J.H.; Zhang, D.Q.; Zhou, G.Y.; Liu, J.X. Soil respiration associated with forest succession in subtropical forests in Dinghushan Biosphere Reserve. Soil Biol. Biochem. 2009, 41, 991–999. [Google Scholar] [CrossRef]
- Shen, C.D.; Yi, W.X.; Sun, Y.M.; Xing, C.P.; Yang, Y.; Peng, S.L.; Li, Z.A. 14C apparent ages and δ13C distribution of forest soils in Dinghushan Natural Reserve. Quat. Sci. 2000, 20, 335–344. (In Chinese) [Google Scholar]
- Brown, S.; Lenart, M.T.; Mo, J.M.; Kong, G.H. Structure and organic matter dynamics of a human-impacted pine forest in a MAB reserve of subtropical China. Biotropica 1995, 27, 276–289. [Google Scholar] [CrossRef]
- Blair, G.J.R.; Lefroy, D.B.; Lisle, L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural system. Aust. J. Agric. Res. 1995, 46, 1459–1466. [Google Scholar] [CrossRef]
- Leavitt, S.W.; Follett, R.F.; Paul, E.A. Estimation of the slow and fast cycling soil organic carbon pools from 6 N HCl hydrolysis. Radiocarbon 1996, 38, 230–231. [Google Scholar]
- Cambardella, C.A.; Elliott, E.T. Particulate soil organic matter changes across a grassland cultivation sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Lu, R.K. Soil Physical and Chemical Analysis, Description of Soil Profiles; Standard Press of China: Beijing, China, 1996; pp. 107–108. (In Chinese) [Google Scholar]
- Tan, Z.; Lal, R.; Owens, L.; Izaurralde, R.C. Distribution of light and heavy fraction of soil organic carbon as related to land use and tillage practice. Soil Tillage Res. 2007, 92, 53–59. [Google Scholar] [CrossRef]
- Six, J.; Callewaert, P.; Lenders, S.; de Gryze, S.; Morris, S.J.; Gregorich, E.G.; Paul, E.A.; Paustian, K. Measuring and understanding carbon storage in afforested soils by physical fractionation. Soil Sci. Soc. Am. J. 2002, 66, 1981–1987. [Google Scholar] [CrossRef]
- Côté, L.; Brown, S.; Pare, D.; Fyles, J.; Bauhus, J. Dynamics of carbon and nitrogen mineralization in relation to stand type, stand age and soil texture in the boreal mixedwood. Soil Biol. Biochem. 2000, 32, 1079–1090. [Google Scholar] [CrossRef]
- Jia, G.M.; Cao, J.; Wang, C.Y.; Wang, G. Microbial biomass and nutrients in soil at the different stages of secondary forest succession in Ziwulin, northwest China. For. Ecol. Manag. 2005, 217, 117–125. [Google Scholar] [CrossRef]
- Liu, S.; Luo, Y.; Huang, Y.H.; Zhou, G.Y. Studies on the community biomass and its allocations of five forest types in Dinghushan Nature Reserve. Ecol. Sci. 2007, 26, 387–393. (In Chinese) [Google Scholar]
- Zhang, D.Q.; Ye, W.H.; Yu, Q.F.; Kong, G.H.; Zhang, Y.C. The litter-fall of representative forests of successional series in Dinghushan. Acta Ecol. Sin. 2000, 20, 938–944. (In Chinese) [Google Scholar]
- Zhou, G.Y.; Liu, S.G.; Li, Z.A.; Zhang, D.Q.; Tang, X.L.; Zhou, C.Y.; Yan, J.H.; Mo, J.M. Old-growth forests can accumulate carbon in soils. Science 2006, 314, 1417. [Google Scholar] [CrossRef] [PubMed]
- Hertel, D.; Hölscher, D.; Köhler, L.; Leuschner, C. Changes in fine root system size and structure during secondary succession in a Costa Rican montane oak forest. Ecol. Stud. 2006, 185, 283–297. [Google Scholar]
- Crow, S.E.; Lajtha, K.; Bowden, R.D.; Yano, Y.; Brant, J.B.; Caldwell, B.A.; Sulzman, E.W. Increased coniferous needle inputs accelerate decomposition of soil carbon in an old-growth forest. For. Ecol. Manag. 2009, 258, 2224–2232. [Google Scholar] [CrossRef]
- Asaye, Z.; Zewdie, S. Fine root dynamics and soil carbon accretion under thinned and un-thinned Cupressus lusitanica stands in Southern Ethiopia. Plant Soil 2003, 366, 261–271. [Google Scholar] [CrossRef]
- Peng, S.L.; Hao, Y.R. The dynamics of forest root and its distribution during succession. Acta Sci. Nat. Univ. Sunyatseni. 2005, 44, 65–69. (In Chinese) [Google Scholar]
- Wen, D.Z.; Wei, P.; Kong, G.H.; Ye, W.H. Production and turnover rate of fine roots in two lower subtropical forest sites at Dinghushan. Acta Phytoecol. Sin. 1999, 23, 361–369. (In Chinese) [Google Scholar]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Monnier, G. Action des matiéres organiques sur la stabilité structurale des sols. Ann. Agron. 1965, 16, 327–400. (In French) [Google Scholar]
- Miller, R.M.; Jastrow, J.D. Hierarchy of root and mycorrhizal fungal interactions with soil aggregation. Soil Biol. Biochem. 1990, 22, 579–584. [Google Scholar] [CrossRef]
- Jastrow, J.D.; Miller, R.M. Soil aggregate stabilization and carbon sequestration: Feedbacks through organomineral associations. In Soil Processes and the Carbon Cycle; Lal, R., Kimble, J.M., et al., Eds.; CRC Press: Boca Raton, FL, USA, 1998; pp. 207–223. [Google Scholar]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.L.; Zuo, W.Y.; Zhao, Z.Y.; Qiu, L.H. Bacterial diversity of different successional stage forest soils in Dinghushan. Acta Microbiol. Sin. 2012, 52, 1489–1496. (In Chinese) [Google Scholar]
- Six, J.; Conant, R.T.; Oaul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Six, J.; Elliott, E.T.; Paustia, K.; Doran, J.W. Aggregation and soil organic matter accumulation in cultivated and native grassland soils. Soil Sci. Soc. Am. J. 1998, 62, 1367–1377. [Google Scholar] [CrossRef]
- Gale, W.J.; Cambardella, C.A.; Bailey, T.B. Root-derived carbon and the formation and stabilization of aggregates. Soil Sci. Soc. Am. J. 2000, 64, 201–207. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, H.; Zhang, L.; Wen, D. Change of Soil Carbon Fractions and Water-Stable Aggregates in a Forest Ecosystem Succession in South China. Forests 2015, 6, 2703-2718. https://doi.org/10.3390/f6082703
Xiang H, Zhang L, Wen D. Change of Soil Carbon Fractions and Water-Stable Aggregates in a Forest Ecosystem Succession in South China. Forests. 2015; 6(8):2703-2718. https://doi.org/10.3390/f6082703
Chicago/Turabian StyleXiang, Huimin, Lingling Zhang, and Dazhi Wen. 2015. "Change of Soil Carbon Fractions and Water-Stable Aggregates in a Forest Ecosystem Succession in South China" Forests 6, no. 8: 2703-2718. https://doi.org/10.3390/f6082703