Evaluation of Predation Capability of Periodontopathogens Bacteria by Bdellovibrio Bacteriovorus HD100. An in Vitro Study

 ,
, 
 , and
, and 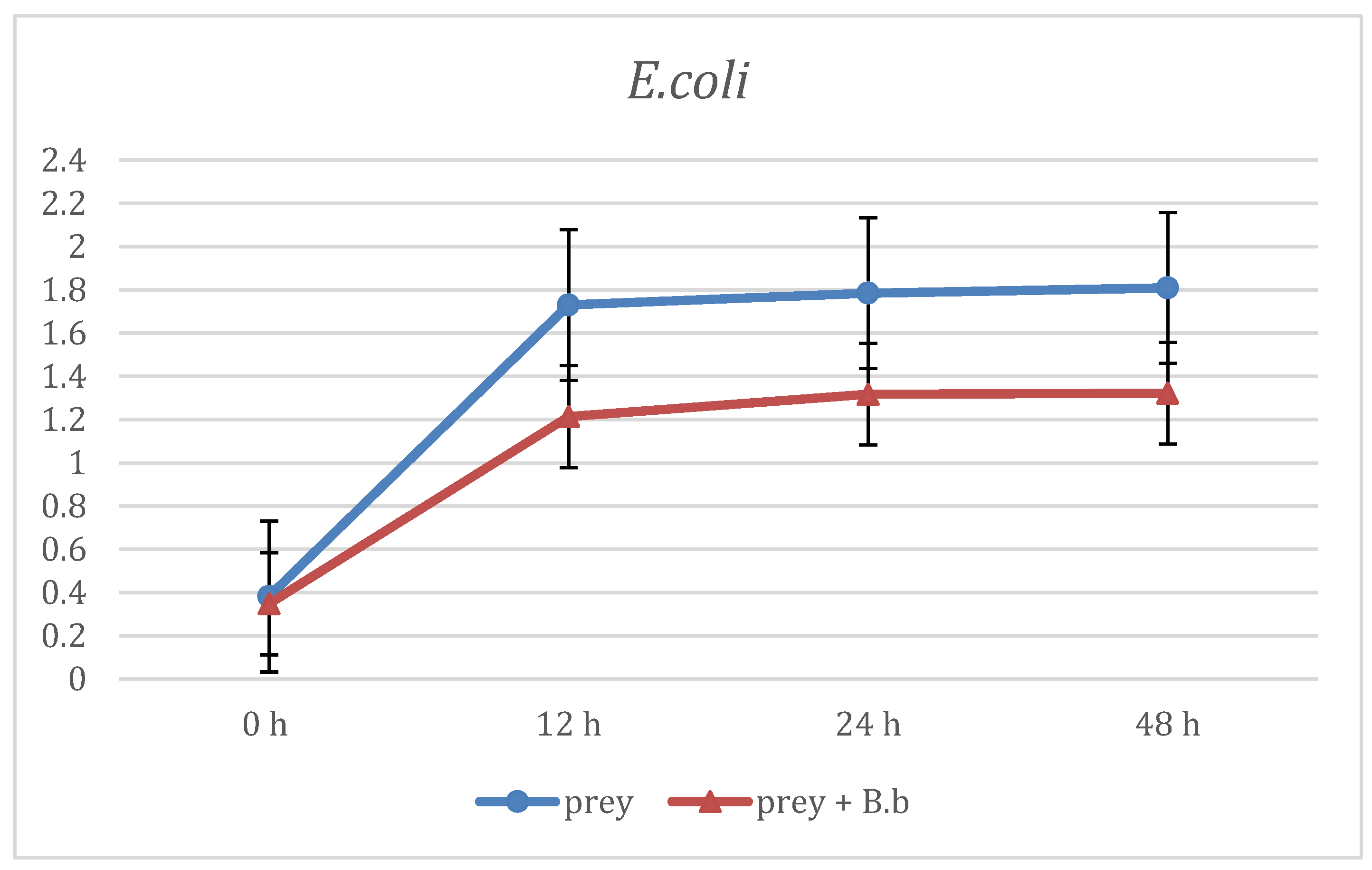{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Strains and Growth Conditions
2.2. Preparation of B. Bacteriovorus Suspensions
2.3. Prey Range Assay
- I Group: Prey without predator as positive control;
- II Group: Prey with predator;
- III Group: Predator without prey as negative control.
2.4. Statistical Analysis
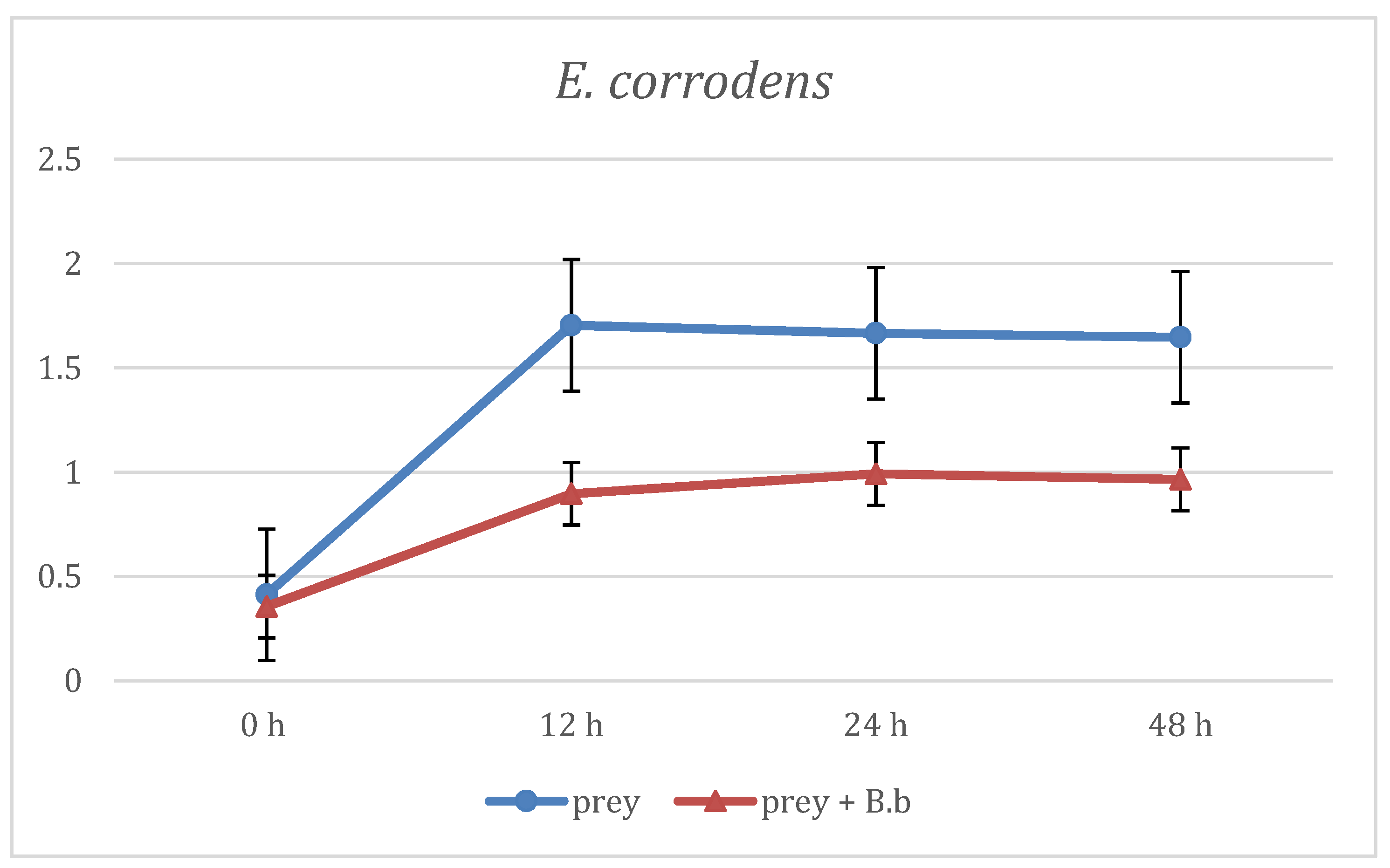
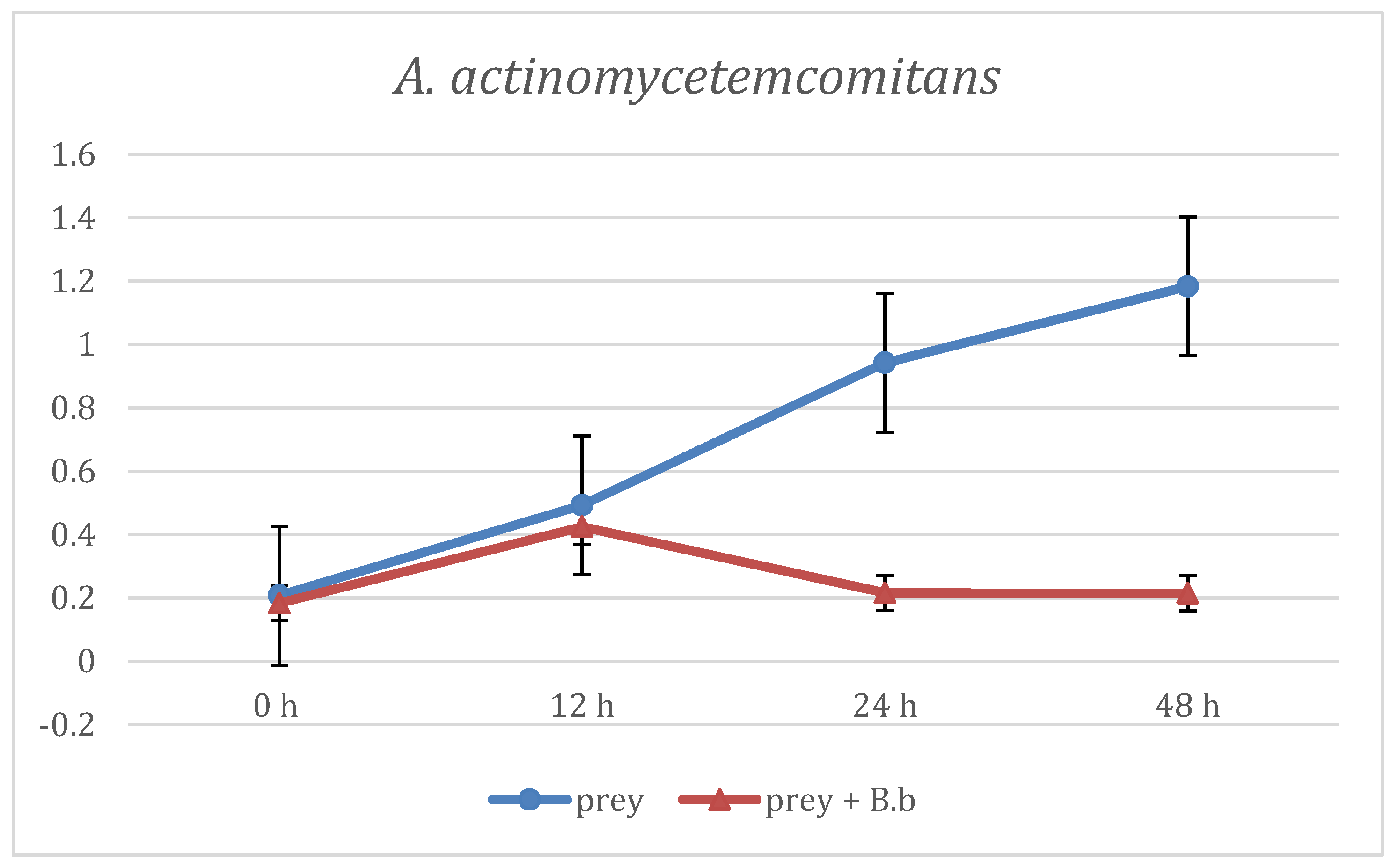
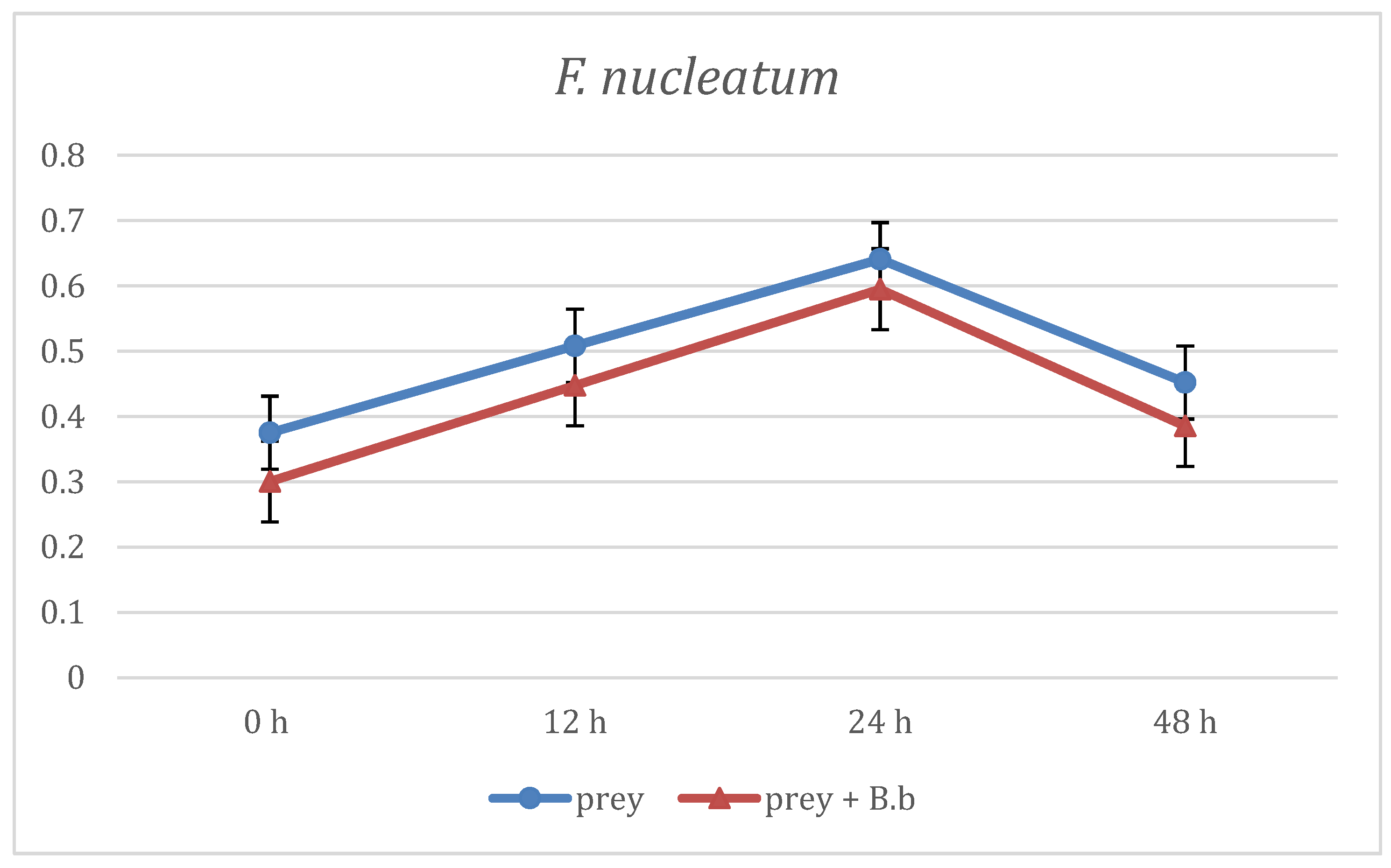
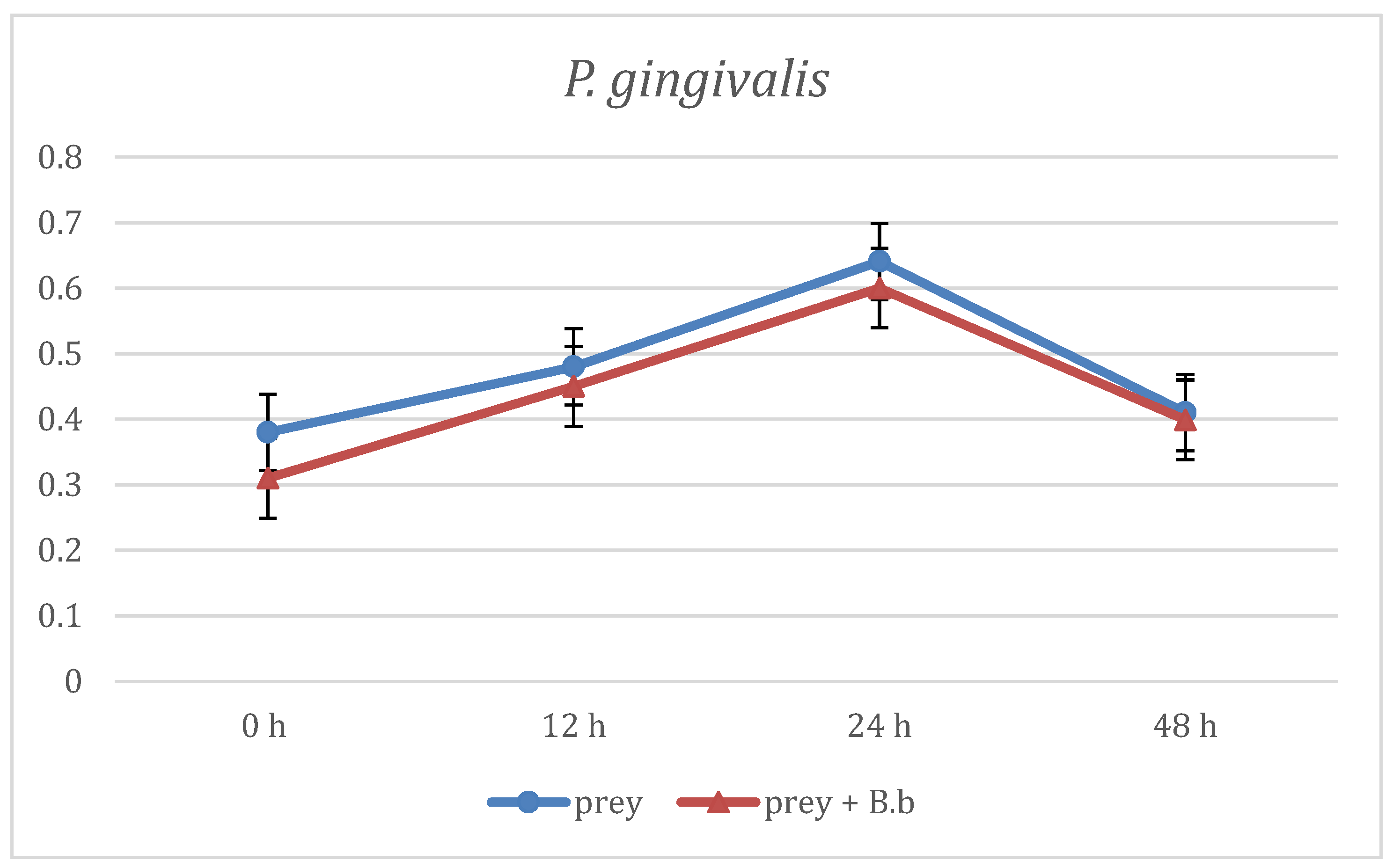
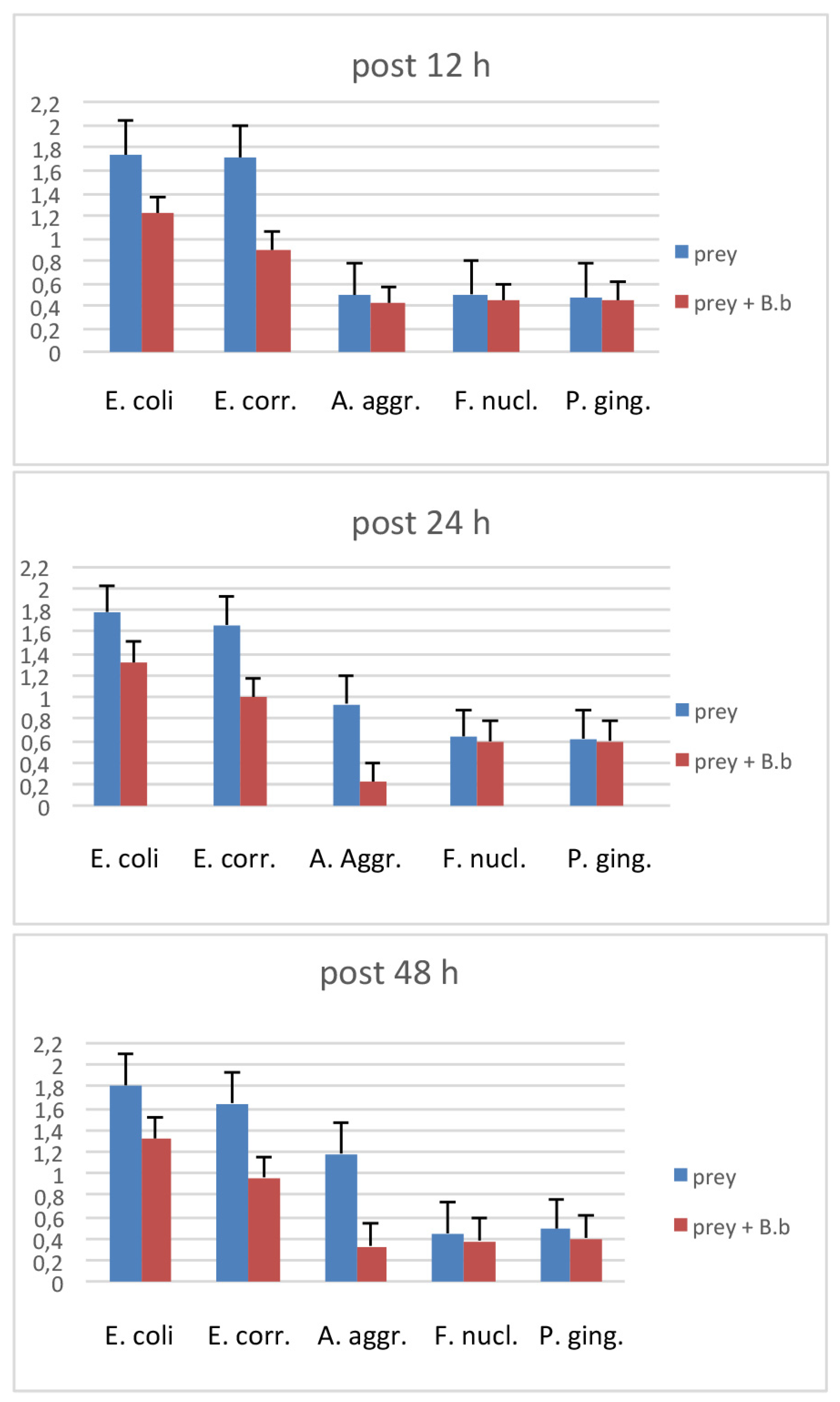
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tonetti, M.S.; Jepsen, S.; Jin, L.; Otomo-Corgel, J. Impact of the global burden of periodontal diseases on health, nutrition and wellbeing of mankind: A call for global action. J. Clin. Periodontol. 2017, 44, 456–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.J.; Lamster, I.B.; Greenspan, J.S.; Pitts, N.B.; Scully, C.; Warnakulasuriya, S. Global burden of oral diseases: Emerging concepts, management and interplay with systemic health. Oral Dis. 2016, 22, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Patini, R.; Staderini, E.; Lajolo, C.; Lopetuso, L.; Mohammed, H.; Rimondini, L.; Rocchetti, V.; Franceschi, F.; Cordaro, M.; Gallenzi, P. Relationship between oral microbiota and periodontal disease: a systematic review. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5775–5788. [Google Scholar] [PubMed]
- Patini, R.; Cantiani, M.; Spagnuolo, G.; Cordaro, M.; Cantiani, M.; Amalfitano, A.; Arcovito, A.; Callà, C.A.M.; Mingrone, G.; Nocca, G. Metabolic syndrome and periodontitis: association with Reactive Oxygen Species production. A pilot study. Open Dent. J. 2017, 11, 621–627. [Google Scholar] [CrossRef]
- Patini, R.; Staderini, E.; Gallenzi, P. Multidisciplinary surgical management of Cowden syndrome: Report of a case. J. Clin. Exp. Dent. 2016, 18, 472–474. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, H.; Varoni, E.M.; Cochis, A.; Cordaro, M.; Gallenzi, P.; Patini, R.; Staderini, E.; Lajolo, C.; Rimondini, L.; Rocchetti, V. Oral dysbiosis in pancreatic cancer and liver cirrhosis: a review of the literature. Biomedicines 2018, 6, 115. [Google Scholar] [CrossRef]
- Vartoukian, S.R.; Palmer, R.M.; Wade, W.G. The division “Synergistes”. Anaerobe 2007, 13, 99–106. [Google Scholar] [CrossRef]
- Rodriguez-Segade, S.; Velasco, D.; Marcos, P.J. Empyema due to Aggregatibacter aphrophilus and Parvimonas micra coinfection. Arch. Bronconeumol. 2015, 51, 254–255. [Google Scholar] [CrossRef]
- Baghban, A.; Gupta, S. Parvimonas micra: A rare case of native joint septic arthritis. Anaerobe 2016, 39, 26–27. [Google Scholar] [CrossRef]
- Roberts, F.A.; Darveau, R.P. Beneficial bacteria of the periodontium. Periodontol. 2000 2002, 30, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Stolp, H.; Starr, M.P. Bdellovibrio bacteriovorus gen. et sp. n., a predatory, ectoparasitic, and bacteriolytic microorganism. Antonie Van Leeuwenhoek 1963, 29, 217–248. [Google Scholar] [CrossRef] [PubMed]
- Sockett, R.E. Predatory lifestyle of Bdellovibrio bacteriovorus. Annu. Rev. Microbiol 2009, 63, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Van Essche, M.; Quirynen, M.; Sliepen, I.; Van Eldere, J.; Teughels, W. Bdellovibrio bacteriovorus attacks Aggregatibacter actinomycetemcomitans. J. Dent. Res. 2009, 88, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Iebba, V.; Santangelo, F.; Totino, V.; Nicoletti, M.; Gagliardi, A.; De Biase, R.V.; Cucchiara, S.; Nencioni, L.; Conte, M.P.; Schippa, S. Higher prevalence and abundance of Bdellovibrio Bacteriovorus in the human gut of healthy subjects. PLoS ONE 2013, 8, e61608. [Google Scholar] [CrossRef]
- Ferlazzo, N.; Currò, M.; Zinellu, A.; Caccamo, D.; Isola, G.; Ventura, V.; Carru, C.; Matarese, G.; Ientile, R. Influence of MTHFR genetic background on P16 and MGMT methylation in oral squamous cell cancer. Int. J. Mol. Sci. 2017, 18, 724. [Google Scholar] [CrossRef] [PubMed]
- Schwudke, D.; Linscheid, M.; Strauch, E.; Appel, B.; Zahringer, U.; Moll, H.; Muller, M.; Brecker, L.; Gronow, S.; Lindner, B. The obligate predatory Bdellovibrio bacteriovorus possesses a neutral lipid a containing α-D-mannoses that replace phosphate residues. Similarities and differences between the lipid As and the lipopolysaccharides of the wild type strain B. bacteriovorus HD100 and its host-independent derivative HI100. J. Biolog. Chem. 2003, 278, 27502–27512. [Google Scholar]
- Van Essche, M.; Quirynen, M.; Sliepen, I.; Loozen, G.; Boon, N.; Van Eldere, J.; Teughels, W. Killing of anaerobic pathogens by predatory bacteria. Molec. Oral Microbiol. 2011, 26, 52–61. [Google Scholar] [CrossRef]
- Dashiff, A.; Kadouri, D.E. Predation of oral pathogens by Bdellovibrio bacteriovorus 109J. Molec. Oral Microbiol. 2011, 26, 19–34. [Google Scholar] [CrossRef]
- Armitage, G.C. Development of a classification system for periodontal diseases and conditions. Ann. Periodontol. 1999, 4, 1–6. [Google Scholar] [CrossRef]
- Lasserre, J.F.; Brecx, M.C.; Toma, S. Oral microbes, biofilm and their role in periodontal and peri-implant diseases. Materials 2018, 11, 1802. [Google Scholar] [CrossRef] [PubMed]
- Cannavale, R.; Matarese, G.; Isola, G.; Grassia, V.; Perillo, L. Early treatment of an ectopic premolar to prevent molar-premolar transposition. Am. J. Orthod. Dentofacial. Orthop. 2013, 143, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Quirynen, M.; De Soete, M.; Dierickx, K.; Van Steenberghe, D. The intra-oral translocation of periodontopathogens jeopardises the outcome of periodontal therapy. A review of the literature. J. Clin. Periodontol. 2001, 28, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Pippi, R.; Santoro, M.; Patini, R. The central odontogenic fibroma: How difficult can be making a preliminary diagnosis. J. Clin. Exp. Dent. 2016, 8, 223–225. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patini, R.; Cattani, P.; Marchetti, S.; Isola, G.; Quaranta, G.; Gallenzi, P. Evaluation of Predation Capability of Periodontopathogens Bacteria by Bdellovibrio Bacteriovorus HD100. An in Vitro Study. Materials 2019, 12, 2008. https://doi.org/10.3390/ma12122008
Patini R, Cattani P, Marchetti S, Isola G, Quaranta G, Gallenzi P. Evaluation of Predation Capability of Periodontopathogens Bacteria by Bdellovibrio Bacteriovorus HD100. An in Vitro Study. Materials. 2019; 12(12):2008. https://doi.org/10.3390/ma12122008
Chicago/Turabian StylePatini, Romeo, Paola Cattani, Simona Marchetti, Gaetano Isola, Gianluca Quaranta, and Patrizia Gallenzi. 2019. "Evaluation of Predation Capability of Periodontopathogens Bacteria by Bdellovibrio Bacteriovorus HD100. An in Vitro Study" Materials 12, no. 12: 2008. https://doi.org/10.3390/ma12122008