
Water Budgets of Managed Forests in Northeast Germany under Climate Change—Results from a Model Study on Forest Monitoring Sites


Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Fitting Infiltration Parameter Infexp
3.2. Model Validation
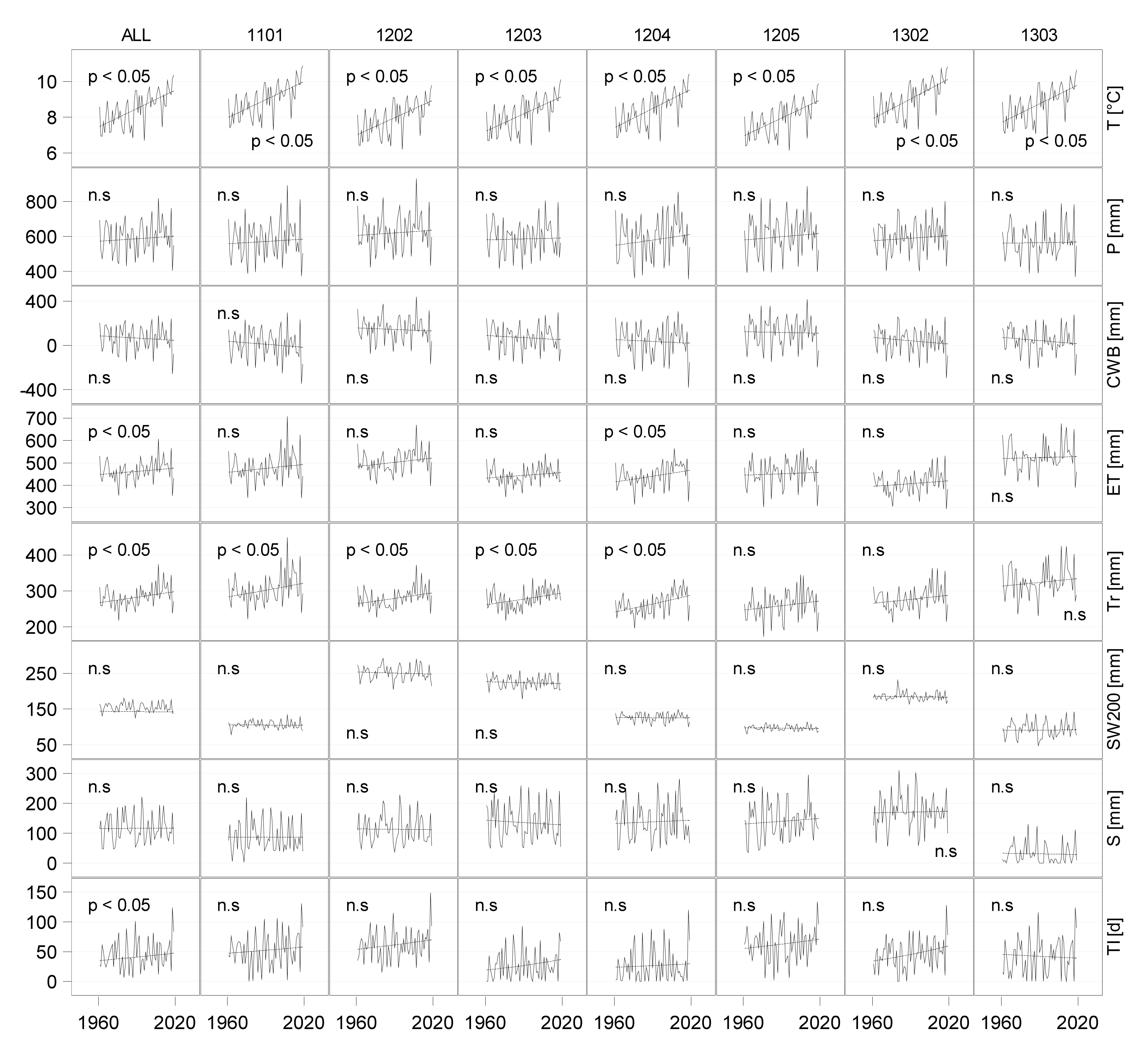
3.3. Water Budgets of Historical Time Series
3.4. Water Budgets under Future Climate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Lasch-Born, P.; Suckow, F.; Gutsch, M.; Reyer, C.; Hauf, Y.; Murawski, A.; Pilz, T. Forests under climate change: Potential risks and opportunities. Meteorol. Z. 2015, 24, 157–172. [Google Scholar] [CrossRef]
- Schwalm, C.W.; Anderegg, W.R.L.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Wolf, A.; et al. Global patterns of drought recovery. Nature 2017, 548, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Hänsel, S.; Ustrnul, Z.; Łupikasza, E.; Skalak, P. Assessing seasonal drought variations and trends over Central Europe. Adv. Water Resour. 2019, 127, 53–75. [Google Scholar] [CrossRef]
- Kaiser, K.; Koch, P.J.; Mauersberger, R.; Stüve, P.; Dreibrodt, J.; Bens, O. Detection and attribution of lake-level dynamics in north-eastern central Europe in recent decades. Reg. Environ. Chang. 2014, 14, 1587–1600. [Google Scholar] [CrossRef]
- Natkhin, M.; Steidl, J.; Dietrich, O.; Dannowski, R.; Lischeid, G. Differentiating between climate effects and forest growth dynamics effects on decreasing groundwater recharge in a lowland region in Northeast Germany. J. Hydrol. 2012, 448–449, 245–254. [Google Scholar] [CrossRef]
- Bauwe, A.; Criegee, C.; Glatzel, S.; Lennartz, B. Model-based analysis of the spatial variability and long-term trends of soil drought at Scots pine stands in northeastern Germany. Eur. J. For. Res. 2012, 131, 1013–1024. [Google Scholar] [CrossRef]
- Kaspar, F.; Müller-Westermeier, G.; Penda, E.; Mächel, H.; Zimmermann, K.; Kaiser-Weiss, A.; Deutschländer, T. Monitoring of climate change in Germany—Data, products and services of Germany’s National Climate Data Centre. Adv. Sci. Res. 2013, 10, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Lian, X.; Piao, S.; Li, L.Z.X.; Li, Y.; Huntingford, C.; Ciais, P.; Cescatti, A.; Janssens, I.A.; Peñuelas, J.; Buermann, W.; et al. Summer soil drying exacerbated by earlier spring greening of northern vegetation. Sci. Adv. 2020, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Deser, C.; Vecchi, G.A.; Collins, M.; Delworth, T.L.; Hall, A.; Hawkins, E.; Johnson, N.C.; Cassou, C.; Giannini, A.; et al. Towards predictive understanding of regional climate change. Nat. Clim. Chang. 2015, 5, 921–930. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hackel, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–756. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2020, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Bose, A.K.; Gessler, A.; Bolte, A.; Bottero, A.; Buras, A.; Cailleret, M.; Camarero, J.J.; Haeni, M.; Hereş, A.M.; Hevia, A.; et al. Growth and resilience responses of Scots pine to extreme droughts across Europe depend on predrought growth conditions. Glob. Change Biol. 2020, 26, 4521–4537. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef]
- Netherer, S.; Panassiti, B.; Pennerstorfer, J.; Matthews, B. Acute Drought Is an Important Driver of Bark Beetle Infestation in Austrian Norway Spruce Stands. Front. For. Glob. Chang. 2019, 2, 39. [Google Scholar] [CrossRef] [Green Version]
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M.J.S. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat. Commun. 2015, 6, 7537. [Google Scholar] [CrossRef] [PubMed]
- Riek, W.; Russ, A. In Zeiten des Standortswandels: Handlungsempfehlungen aus BZE und Regionalisierung für die nachhaltige Waldnutzung. In Eberswalder Forstliche Schriftenreihe, Bd. 67; Landesbetrieb Forst Brandenburg, Landeskompetenzzentrum Forst Eberswalde: Eberswalde, Germany, 2019; pp. 33–42. [Google Scholar]
- Riek, W.; Russ, A.; Grüll, M. Zur Abschätzung des standörtlichen Anbaurisikos von Baumarten im Klimawandel im nordostdeutschen Tiefland. In Eberswalder Forstliche Schriftenreihe, Bd. 69; Landesbetrieb Forst Brandenburg, Landeskompetenzzentrum Forst Eberswalde: Eberswalde, Germany, 2020; pp. 48–70. [Google Scholar]
- Fleck, S.; Cools, N.; De Vos, B.; Meesenburg, H.; Fischer, R. The Level II aggregated forest soil condition database links soil physicochemical and hydraulic properties with long-term observations of forest condition in Europe. Ann. For. Sci. 2016, 73, 945–957. [Google Scholar] [CrossRef] [Green Version]
- Jacob, D.; Petersen, J.; Eggert, B.; Alias, A.; Christensen, O.B.; Bouwer, L.M.; Braun, A.; Colette, A.; Déqué, M.; Georgievski, G.; et al. EURO-CORDEX: New high-resolution climate change projections for European impact research. Reg. Environ. Chang. 2014, 14, 563–578. [Google Scholar] [CrossRef]
- Moss, R.H.; Edmonds, J.A.; Hibbard, K.A.; Manning, M.R.; Rose, S.K.; van Vuuren, D.P.; Carter, T.R.; Emori, S.; Kainuma, M.; Kram, T.; et al. The Next Generation of Scenarios for Climate Change Research and Assessment. Nature 2010, 463, 747–756. [Google Scholar] [CrossRef]
- Samaniego, L.; Thober, S.; Kumar, R.; Wanders, N.; Rakovec, O.; Pan, M.; Zink, M.; Sheffield, J.; Wood, E.F.; Marx, A. Anthropogenic warming exacerbates European soil moisture droughts. Nat. Clim. Change 2018, 8, 421–426. [Google Scholar] [CrossRef]
- Ruosteenoja, K.; Markkanen, T.; Venäläinen, A.; Räisänen, P.; Peltola, H. Seasonal soil moisture and drought occurrence in Europe in CMIP5 projections for the 21st century. Clim. Dyn. 2018, 50, 1177–1192. [Google Scholar] [CrossRef] [Green Version]
- Grillakis, M.G. Increase in severe and extreme soil moisture droughts for Europe under climate change. Sci. Total Environ. 2019, 660, 1245–1255. [Google Scholar] [CrossRef]
- Marx, A.; Kumar, R.; Thober, S.; Rakovec, O.; Wanders, N.; Zink, M.; Wood, E.F.; Pan, M.; Sheffield, J.; Samaniego, L. Climate change alters low flows in Europe under global warming of 1.5, 2, and 3 °C. Hydrol. Earth Syst. Sci. 2018, 22, 1017–1032. [Google Scholar] [CrossRef] [Green Version]
- Jing, M.; Kumar, R.; Heße, F.; Thober, S.; Rakovec, O.; Samaniego, L.; Attinger, S. Assessing the response of groundwater quantity and travel time distribution to 1.5, 2, and 3 °C global warming in a mesoscale central German basin. Hydrol. Earth Syst. Sci. 2020, 24, 1511–1526. [Google Scholar] [CrossRef] [Green Version]
- Riek, W.; Russ, A.; Martin, J. Soil acidification and nutrient sustainability of forest ecosystems in the northeastern German lowlands—Results of the national forest soil inventory. Folia For. Pol. Series A 2012, 54, 187–195. [Google Scholar]
- Russ, A.; Riek, W.; Hentschel, R.; Hannemann, J.; Barth, R.; Becker, F. Wasserhaushalt im Trockenjahr 2018 —Ergebnisse aus dem Level II Programm in Brandenburg. In Eberswalder Forstliche Schriftenreihe, Bd. 67; Landesbetrieb Forst Brandenburg, Landeskompetenzzentrum Forst Eberswalde: Eberswalde, Germany, 2019; pp. 11–24. [Google Scholar]
- Raspe, S.; Beuker, E.; Preuhsler, T.; Bastrup-Birk, A. Part IX: Meteorological Measurements. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-ordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 14. [Google Scholar]
- Dobbertin, M.; Neumann, M. Part V: Tree Growth. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-ordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 17. [Google Scholar]
- Canullo, R.; Starlinger, F.; Granke, O.; Fischer, R.; Aamlid, D. Part VI.1: Assessment of Ground Vegetation. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-ordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 12. [Google Scholar]
- Cools, N.; De Vos, B. Part X: Sampling and Analysis of Soil. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-ordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 115. [Google Scholar]
- Fleck, S.; Raspe, S.; Cater, M.; Schleppi, P.; Ukonmaanaho, L.; Greve, M.; Hertel, C.; Weis, W.; Rumpf, S.; Thimonier, A.; et al. Part XVII: Leaf Area Measurements. In Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; UNECE ICP Forests Programme Co-ordinating Centre, Ed.; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; p. 34. [Google Scholar]
- Hammel, K.; Kennel, M. Charakterisierung und Analyse der Wasserverfügbarkeit und des Wasserhaushalts von Waldstandorten in Bayern mit dem Simulationsmodell Brook90. In Forstliche Forschungsberichte München 185; Heinrich Frank: München, Germany, 2001; p. 135. [Google Scholar]
- Federer, C.A. BROOK 90: A Simulation Model for Evaporation, Soil Water, and Streamflow. 2002. Available online: http://www.ecoshift.net/brook/brook90.htm (accessed on 31 July 2020).
- Shuttleworth, W.J.; Wallace, J.S. Evaporation from sparse crops-an energy combination theory. Q. J. R. Meteorol. Soc. 1985, 111, 839–855. [Google Scholar] [CrossRef]
- Federer, C.A.; Vörösmarty, C.; Fekete, B. Intercomparison of methods for calculating potential evaporation in regional and global water balance models. Water Resour. Res. 1996, 32, 2315–2321. [Google Scholar] [CrossRef]
- Rutter, A.J.; Kershaw, K.A.; Robins, P.C.; Morton, A.J. A predictive Model of Rainfall Interception in Forests. Agric. Meteorol. 1972, 9, 367–384. [Google Scholar] [CrossRef]
- Jarvis, P.G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Phil. Trans. R. Soc. Lond. B 1976, 273, 593–610. [Google Scholar] [CrossRef]
- Clapp, R.B.; Hornberger, G.M. Empirical equations for some soil hydraulic properties. Water Resour. Res. 1978, 14, 601–604. [Google Scholar] [CrossRef] [Green Version]
- Van Genuchten, M.T. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Deutscher Wetterdienst. Klimastationsdaten des Climate Data Center (CDC). 2019. Available online: ftp://opendata.dwd.de/climate_environment/CDC/observations_germany/climate/daily/ (accessed on 30 September 2020).
- Ziche, D.; Seidling, W. Homogenisation of climate time series from ICP Forests Level II monitoring sites in Germany based on interpolated climate data. Ann. For. Sci. 2010, 67, 804. [Google Scholar] [CrossRef] [Green Version]
- Lange, S. Trend-preserving bias adjustment and statistical downscaling with ISIMIP3BASD (v1.0). Geosci. Model. Dev. 2019, 12, 3055–3070. [Google Scholar] [CrossRef] [Green Version]
- Hübener, H.; Bülow, K.; Fooken, C.; Früh, B.; Hoffmann, P.; Höpp, S.; Keuler, K.; Menz, C.; Mohr, V.; Radtke, K.; et al. ReKliEs-De Ergebnisbericht. World Data Center for Climate (WDCC) at DKRZ. 2017. Available online: https://cera-www.dkrz.de/WDCC/ui/cerasearch/entry?acronym=ReKliEs-De_Ergebnisbericht (accessed on 31 October 2019). [CrossRef]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999, 397, 659. [Google Scholar] [CrossRef]
- Nagel, J. ForestTools2: Forstliche Software-Sammlung; Selbstverlag, J. Nagel: Göttingen, Germany, 2007. [Google Scholar]
- Ziche, D.; Grüneberg, E.; Hilbrig, L.; Höhle, J.; Kompa, T.; Liski, J.; Repo, A.; Wellbrock, N. Comparing soil inventory with modelling: Carbon balance in central European forest soils varies among forest types. Sci. Total Environ. 2019, 647, 1573–1585. [Google Scholar] [CrossRef] [PubMed]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; Van Groenendael, J.M.; Klimes, L.; Klimesová, J.; et al. The LEDA Traitbase: A database of life-history traits of Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Ahrends, B.; Penne, C.; Panferov, O. Impact of Target Diameter Harvesting on Spatial and Temporal Pattern of Drought Risk in Forest Ecosystems Under Climate Change Conditions. Open Geogr. J. 2010, 3, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Bolte, A.; Anders, S.; Roloff, A. Schätzmodelle zum oberirdischen Vorrat der Waldbodenflora an Trockensubstanz, Kohlenstoff und Makronährelementen. Allg. Forst Jagdztg. 2002, 173, 57–66. [Google Scholar]
- Bolte, A.; Czajkowski, T.; Bielefeldt, J.; Wolff, B.; Heinrichs, S. Estimating aboveground biomass of forest tree and shrub understorey based on relevées. Forstarchiv 2009, 80, 222–228. [Google Scholar]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Von Wilpert, K.; Hartmann, P.; Puhlmann, H.; Schmidt-Walter, P.; Meesenburg, H.; Müller, J. Bodenwasserhaushalt und Trockenstress. In Dynamik und Räumliche MUSTER Forstlicher Standorte in Deutschland; Wellbrock, N., Bolte, A., Flessa, H., Eds.; Thünen Report: Braunschweig, Germany, 2016; Volume 43, pp. 343–386. [Google Scholar] [CrossRef]
- Gale, M.R.; Grigal, D.F. Vertical root distributions of northern tree species in relation to successional status. Can. J. For. Res. 1987, 17, 829–834. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Wessolek, G.; Duijnisveld, W.H.M.; Trinks, S. Hydro-Pedotransferfunktionen zur Berechnung der Sickerwasserrate aus dem Boden: Das TUB-BGR-Verfahren. In Bodenphysikalische Kennwerte und Berechnungsverfahren für die Praxis; Wessolek, G., Kaupenjohann, M., Renger, M., Eds.; Rote Reihe 40, TU-Berlin: Berlin, Germany, 2009; pp. 66–80. [Google Scholar]
- Schmidt-Walter, P. Brook90r: Run the LWF-BROOK90 hydrological model from within R. R-package v1.1.1. 2019. Available online: https://github.com/pschmidtwalter/brook90r (accessed on 1 September 2019).
- Breuer, L.; Eckhardt, K.; Frede, H.G. Plant parameter values for models in temperate climates. Ecol. Model. 2003, 169, 237–293. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop. Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO Irrigation and drainage paper 56; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; p. 300. [Google Scholar]
- Shaw, R.H.; Laing, D.R. Moisture stress and plant response. In Plant Environment and Efficient Water Use; Pierre, W.H., Kirkham, D., Pesek, J., Shaw, R., Eds.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1966; pp. 73–94. [Google Scholar] [CrossRef]
- Schwärzel, K.; Feger, K.H.; Häntzschel, J.; Menzer, A.; Spank, U.; Clausnitzer, F.; Köstner, B.; Bernhofer, C. A novel approach in model-based mapping of soil water conditions at forest sites. For. Ecol. Manag. 2009, 258, 2163–2174. [Google Scholar] [CrossRef]
- Joe, H.; Zhu, R. Generalized Poisson Distribution: The Property of Mixture of Poisson and Comparison with Negative Binomial Distribution. Biom. J. 2005, 47, 219–229. [Google Scholar] [CrossRef] [PubMed]
- David, J.S.; Valente, F.; Gash, J.H. Evaporation of Intercepted Rainfall. In Encyclopedia of Hydrological Sciences; Anderson, M.G., McDonnell, J.J., Eds.; John Wiley & Sons, Ltd: New York, NY, USA, 2005. [Google Scholar] [CrossRef]
- Krämer, I.; Hölscher, D. Rainfall partitioning along a tree diversity gradient in a deciduous old-growth forest in Central Germany. Ecohydrogeomorphology 2009, 2, 102–114. [Google Scholar] [CrossRef]
- Bogner, C.; Gaul, D.; Kolb, A.; Schmiedinger, I.; Huwe, B. Investigating flow mechanisms in a forest soil by mixed-effects modelling. Eur. J. Soil Sci. 2010, 61, 1079–1090. [Google Scholar] [CrossRef]
- Schwärzel, K.; Ebermann, S.; Schalling, N. Evidence of double funneling of beech trees by visualization of flow pathways using dye tracer. J. Hydrol. 2012, 470–471, 184–192. [Google Scholar] [CrossRef]
- Spencer, S.A.; van Meerveld, H.J. Double funneling in a mature coastal British Columbia forest: Spatial patterns of stemflow after infiltration. Hydrol. Process. 2016, 30, 4185–4201. [Google Scholar] [CrossRef]
- Carlyle-Moses, D.E.; Iida, S.; Germer, S.; Llorens, P.; Michalzik, B.; Nanko, K.; Tischer, A.; Levia, D.F. Expressing stemflow commensurate with its ecohydrological importance. Adv. Water Resour. 2018, 121, 472–479. [Google Scholar] [CrossRef]
- Metzger, J.C.; Wutzler, T.; Dalla Valle, N.; Filipzik, J.; Grauer, C.; Lehmann, R.; Roggenbuck, M.; Schelhorn, D.; Weckmüller, J.; Küsel, K.; et al. Vegetation impacts soil water content patterns by shaping canopy water fluxes and soil properties. Hydrol. Process. 2017, 31, 3783–3795. [Google Scholar] [CrossRef]
- Cienciala, E.; Kučera, J.; Lindroth, A.; Čermàk, J.; Grelle, A.; Halldin, S. Canopy transpiration from a boreal forest in Sweden during a dry year. Agric. For. Meteorol. 1997, 86, 157–167. [Google Scholar] [CrossRef]
- Poyatos, R.; Llorens, P.; Gallart, F. Transpiration of montane Pinus sylvestris L. and Quercus pubescens Willd. forest stands measured with sap flow sensors in NE Spain. Hydrol. Earth Syst. Sci. 2005, 9, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Vincke, C.; Thiry, Y. Water table is a relevant source for water uptake by a Scots pine (Pinus sylvestris L.) stand: Evidences from continuous evapotranspiration and water table monitoring. Agric. For. Meteorol. 2008, 148, 1419–1432. [Google Scholar] [CrossRef]
- Müller, J. Forestry and water budget of the lowlands in northeast Germany—consequences for the choice of tree species and for forest management. J. Water Land Dev. 2009, 13, 133–148. [Google Scholar] [CrossRef] [Green Version]
- Gebauer, T. Water turnover in species-rich and species-poor deciduous forests: Xylem sap flow and canopy transpiration. Ph.D. Thesis, Göttingen Centre for Biodiversity and Ecology, University of Göttingen, Göttingen, Germany, 2010; p. 146. [Google Scholar]
- Hentschel, R.; Bittner, S.; Janott, M.; Biernath, C.; Holst, J.; Ferrio, J.P.; Gessler, A.; Priesack, E. Simulation of stand transpiration based on a xylem water flow model for individual trees. Agric. For. Meteorol. 2013, 182–183, 31–42. [Google Scholar] [CrossRef]
- Keitel, C.; Adams, M.A.; Holst, T.; Matzarakis, A.; Mayer, H.; Rennenberg, H.; Gessler, A. Carbon and oxygen isotope composition of organic compounds in the phloem sap provides a short-term measure for stomatal conductance of European beech (Fagus sylvatica L.). Plant. Cell Environ. 2003, 26, 1157–1168. [Google Scholar] [CrossRef] [Green Version]
- Köstner, B. Evaporation and transpiration from forests in Central Europe–relevance of patch-level studies for spatial scaling. Meteorol. Atmos. Phys. 2001, 82, 69–82. [Google Scholar] [CrossRef]
- Schipka, F.; Heimann, J.; Leuschner, C. Regional variation in canopy transpiration of Central European beech forests. Oecologia 2005, 143, 260–270. [Google Scholar] [CrossRef]
- Wang, K.; Dickinson, R. A review of global terrestrial evapotranspiration: Observation, modeling, climatology, and climatic variability. Rev. Geophys. 2012, 50, 1–54. [Google Scholar] [CrossRef]
- Jung, M.; Reichstein, M.; Ciais, P.; Seneviratne, S.I.; Sheffield, J.; Goulden, M.L.; Bonan, G.; Cescatti, A.; Chen, J.; De Jeu, R.; et al. Recent decline in the global land evapotranspiration trend due to limited moisture supply. Nature 2010, 467, 951–954. [Google Scholar] [CrossRef] [PubMed]
- Tyree, M.T. Hydraulic limits on tree performance: Transpiration, carbon gain and growth of trees. Trees 2003, 17, 95–100. [Google Scholar] [CrossRef] [Green Version]
- LM (Ministerium für Landwirtschaft und Umwelt Mecklenburg-Vorpommern). Waldzustandsbericht 2019—Ergebnisse der Waldzustandserhebung; Ministerium für Landwirtschaft und Umwelt Mecklenburg-Vorpommern: Schwerin, Germany, 2019; p. 46. [Google Scholar]
- MLUK (Ministerium für Ländliche Entwicklung, Umwelt und Klimaschutz des Landes Brandenburg). Waldschutzbericht 2019; Landeskompetenzzentrum Forst Eberswalde: Eberswalde, Germany, 2019; p. 50. [Google Scholar]
- Wullschleger, S.D.; Meinzer, F.C.; Vertessy, R.A. A review of whole-plant water use studies in tree. Tree Physiol. 1998, 18, 499–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betsch, P.; Bonal, D.; Breda, N.; Montpied, P.; Peiffer, M.; Tuzet, A.; Granier, A. Drought effects on water relations in beech: The contribution of exchangeable water reservoirs. Agric. For. Meteorol. 2011, 151, 531–543. [Google Scholar] [CrossRef]
- Tor-Ngern, P.; Oren, R.; Oishi, A.C.; Uebelherr, J.M.; Palmroth, S.; Tarvainen, L.; Ottosson-Löfvenius, M.; Linder, S.; Domec, J.C.; Näsholm, T. Ecophysiological variation of transpiration of pine forests: Synthesis of new and published results. Ecol. Appl. 2017, 27, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Steckel, M.; del Río, M.; Heym, M.; Aldea, J.; Bielak, K.; Brazaitis, G.; Černý, J.; Coll, L.; Collet, C.; Ehbrecht, M.; et al. Species mixing reduces drought susceptibility of Scots pine (Pinus sylvestris L.) and oak (Quercus robur L., Quercus petraea (Matt.) Liebl.)—Site water supply and fertility modify the mixing effect. For. Ecol. Manag. 2020, 461, 117908. [Google Scholar] [CrossRef]
- Spathelf, P.; Ammer, C. Forest management of scots pine (Pinus sylvestris L) in northern Germany—A brief review of the history and current trends. Forstarchiv 2015, 86, 59–66. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Grau, J.M.; de la Cruz, A.C.; Gil, P.M.; Minaya, M.; Fernández-Cancio, Á. Analysing atmospheric processes and climatic drivers of tree defoliation to determine forest vulnerability to climate warming. Forests 2017, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Bugmann, H.; Palahí, M.; Bontemps, J.; Tomé, M. Trends in modeling to address forest management and environmental challenges in Europe. For. Syst. 2010, 19, 3–7. [Google Scholar] [CrossRef]
- GISTEMP Team. GISS Surface Temperature Analysis (GISTEMP), version 4. In NASA Goddard Institute for Space Studies; 2020. Available online: https://data.giss.nasa.gov/gistemp/ (accessed on 25 October 2020).
- Deutscher Wetterdienst (DWD). Klimazeitreihen. 2020. Available online: https://www.dwd.de/DE/leistungen/zeitreihen/zeitreihen.html (accessed on 25 October 2020).
- Garfinkel, C.I.; Adam, O.; Morin, E.; Enzel, Y.; Elbaum, E.; Bartov, M.; Rostkier-Edelstein, D.; Dayan, U. The Role of Zonally Averaged Climate Change in Contributing to Intermodel Spread in CMIP5 Predicted Local Precipitation Changes. J. Clim. 2020, 33, 1141–1154. [Google Scholar] [CrossRef]
- Huang, F.; Xu, Z.; Guo, W. The linkage between CMIP5 climate models’ abilities to simulate precipitation and vector winds. Clim. Dyn. 2020, 54, 4953–4970. [Google Scholar] [CrossRef]
- Christensen, O.B.; Kjellström, E. Partitioning uncertainty components of mean climate and climate change in a large ensemble of European regional climate model projections. Clim. Dyn. 2020, 54, 4293–4308. [Google Scholar] [CrossRef]
- Song, J.; Wan, S.; Piao, S.; Knapp, A.K.; Classen, A.T.; Vicca, S.; Ciais, P.; Hovenden, M.J.; Leuzinger, S.; Beier, C.; et al. A meta-analysis of 1,119 manipulative experiments on terrestrial carbon-cycling responses to global change. Nat. Ecol. Evol. 2019, 3, 1309–1320. [Google Scholar] [CrossRef] [Green Version]
- Senf, C.; Sebald, J.; Seidl, R. Increasing canopy mortality challenges the future of Europe’s forests. bioRxiv 2020. in review. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | 1101 | 1202 | 1203 | 1204 | 1205 | 1302 | 1303 |
|---|---|---|---|---|---|---|---|
| Species | pine/oak | pine | pine | pine | pine | beech | pine |
| Age 1 | 149/74 | 94 | 114 | 114 | 94 | 95 | 88 |
| Top height 2 [m] | 25.5 | 30.3 | 27.6 | 26.7 | 29.4 | 27.0 | 25.4 |
| Basal area 2 [m²] | 22.9 (34.9) | 49.8 (53.8) | 37.7 (37.8) | 32.8 (33.0) | 30.9 (32.7) | 30.4 | 31.9 |
| Stems per ha 2 [m²] | 147 (563) | 525 (1034) | 338 (381) | 422 (513) | 316 (806) | 656 | 556 |
| GV—Cover 3 [%] | 53 | 68 | 95 | 72 | 70 | 1 | 96 |
| Soil type | Brunic Areno-sol | Lamellic Areno-sol | Albic Podzol | Brunic Areno-sol | Brunic Areno-sol | Brunic Areno-sol | Albic Podzol |
| Parent material | Glacio-fluvial deposits | Glacio-fluvial deposits | Eolian sands | Glacio-fluvial deposits | Glacio-fluvial deposits | Glacio-fluvial deposits | Glacio-fluvial deposits |
| RCP | Model | T [°C] | P [mm] | PET [mm] | CWB [mm] | ET [mm] | Tr [mm] | S [mm] | SW200 [mm] | TI [d] |
|---|---|---|---|---|---|---|---|---|---|---|
| hist | 1961–1990 | 8.0 ± 0.1 a | 584 ± 16 a | 513 ± 6 a | 71 ± 20 a | 454 ± 7 a | 274 ± 4 a | 117 ± 9 a | 155 ± 2 a | 42 ± 4 a,b,c |
| hist | 1991–2019 | 9.0 ± 0.1 b | 609 ± 18 a,b | 548 ± 7 b | 61 ± 22 a | 480 ± 9 b | 296 ± 6 b | 116 ± 8 a | 154 ± 2 a | 49 ± 5 a,c |
| 2.6 | ECE/RCA4 | 9.7 ± 0.1 c | 629 ± 17 a,b | 642 ± 10 c | −13 ± 24 ab | 472 ± 8 a,b | 286 ± 4 a,b | 143 ± 8 a,b | 162 ± 3 a | 34 ± 2 a,b,c |
| 2.6 | MPI/Remo | 9.6 ± 0.1 b,c | 638 ± 17 a,b | 643 ± 8 c | −5 ± 23 ab | 457 ± 5 a,b | 275 ± 3 a,b | 180 ± 11 a,b | 169 ± 3 a | 29 ± 4 a,b |
| 8.5 | ECE/RCA4 | 12.6 ± 0.1 d | 654 ± 19 b,c | 718 ± 12 d | −64 ± 27 b | 500 ± 8 a,b | 306 ± 4 a,b | 137 ± 10 a,b | 156 ± 2 a | 51 ± 3 c |
| 8.5 | MPI/Remo | 11.6 ± 0.1 e | 716 ± 15 c | 655 ± 10 c | 61 ± 22 a | 488 ± 6 a,b | 287 ± 4 a,b | 208 ± 13 b | 176 ± 2 a | 25 ± 3 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziche, D.; Riek, W.; Russ, A.; Hentschel, R.; Martin, J. Water Budgets of Managed Forests in Northeast Germany under Climate Change—Results from a Model Study on Forest Monitoring Sites. Appl. Sci. 2021, 11, 2403. https://doi.org/10.3390/app11052403
Ziche D, Riek W, Russ A, Hentschel R, Martin J. Water Budgets of Managed Forests in Northeast Germany under Climate Change—Results from a Model Study on Forest Monitoring Sites. Applied Sciences. 2021; 11(5):2403. https://doi.org/10.3390/app11052403
Chicago/Turabian StyleZiche, Daniel, Winfried Riek, Alexander Russ, Rainer Hentschel, and Jan Martin. 2021. "Water Budgets of Managed Forests in Northeast Germany under Climate Change—Results from a Model Study on Forest Monitoring Sites" Applied Sciences 11, no. 5: 2403. https://doi.org/10.3390/app11052403