Bringing the Natives Back: Identifying and Alleviating Establishment Limitations of Native Hardwood Species in a Conifer Plantation

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Thinning Treatments and Establishment of Transects
2.3. Direct Seeding of Fresh and Germinated Seeds
2.4. Controlling Seed Predation
2.4.1. Physical Barriers
2.4.2. Seed Concealment
2.4.3. Chemical Repellents
2.5. Planting Seedlings of Various Ages
2.6. Data Analysis
3. Results
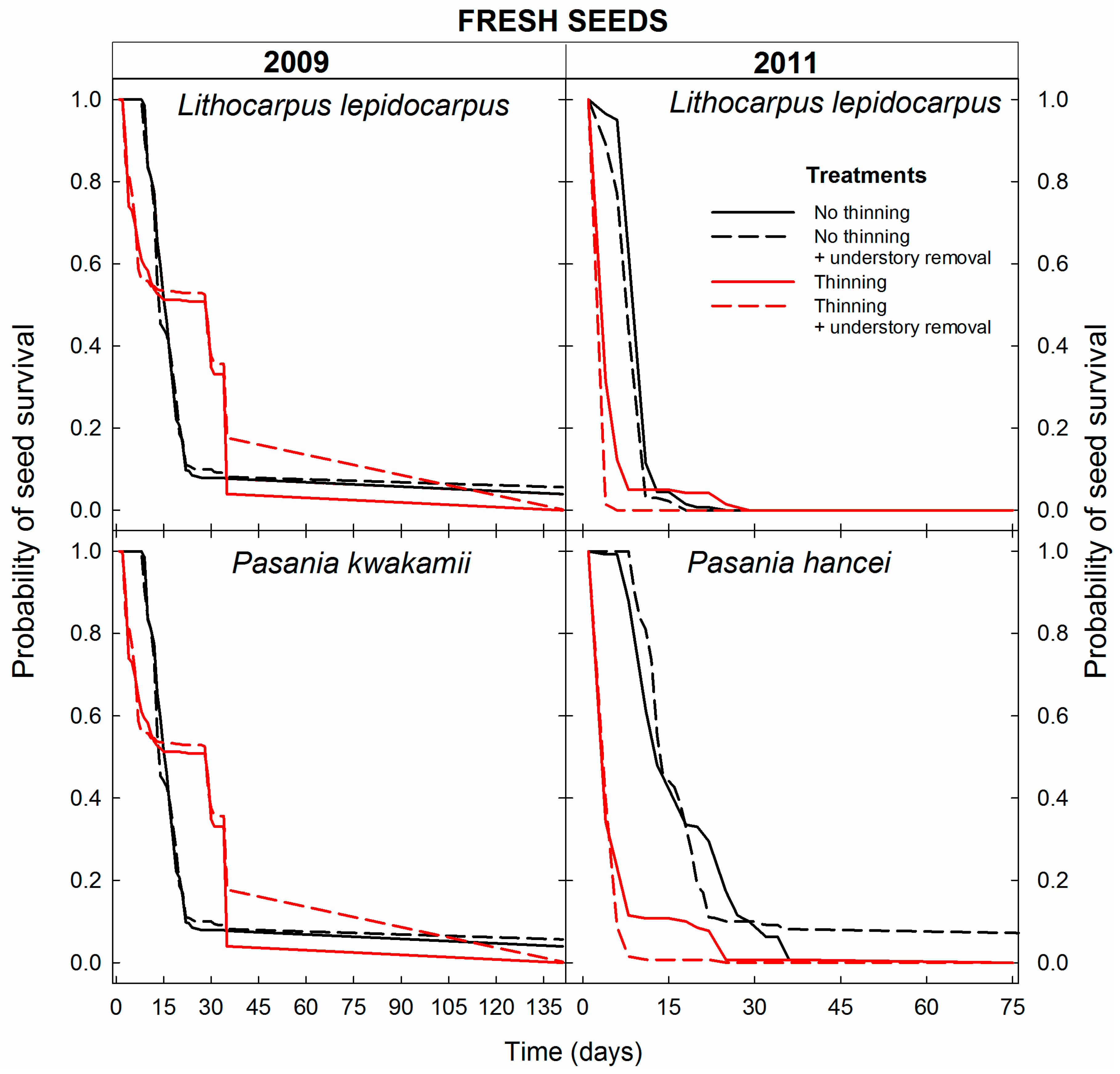
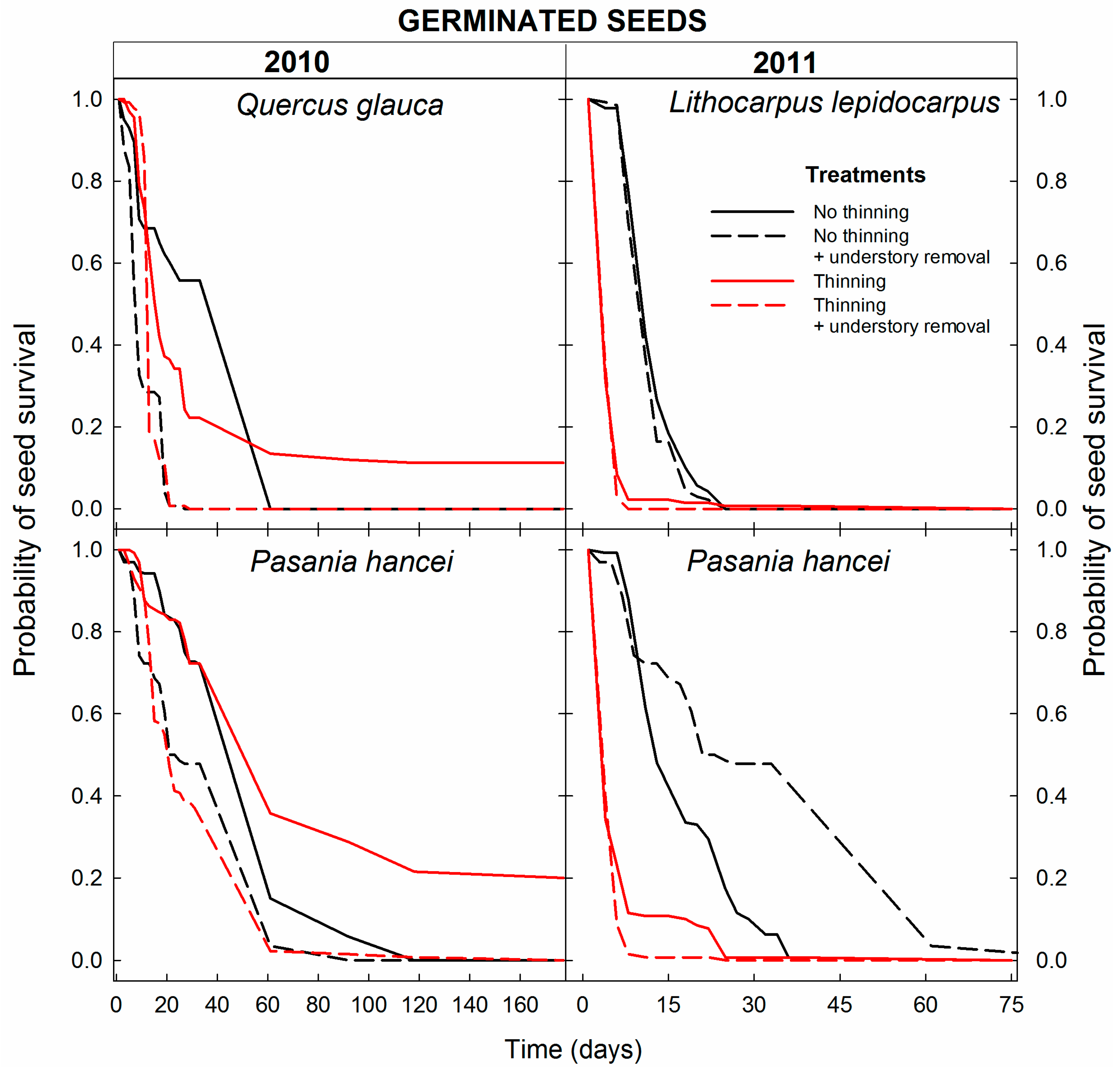
3.1. Seed Survival
3.2. Controlling Seed Predation
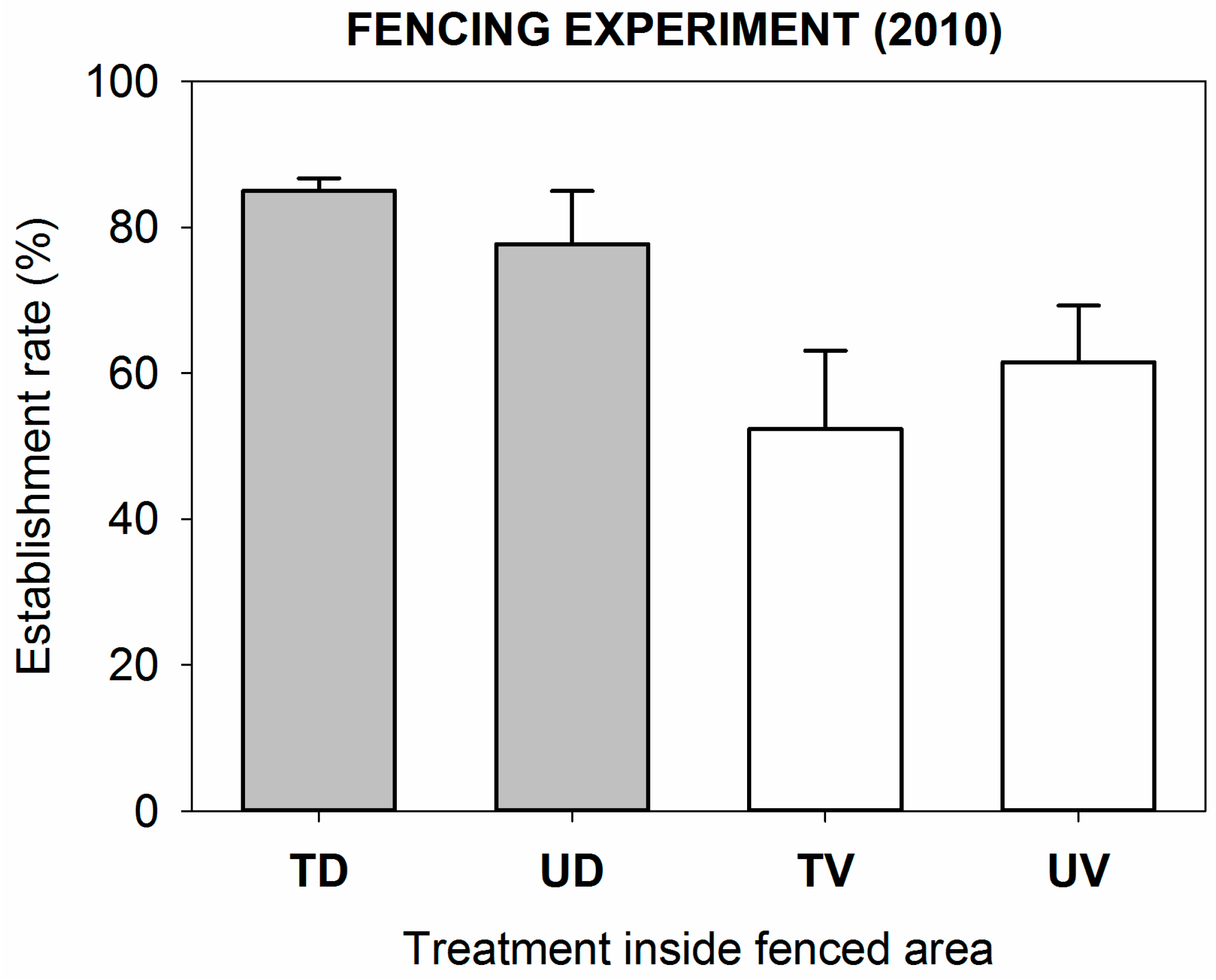
3.2.1. Physical Barriers
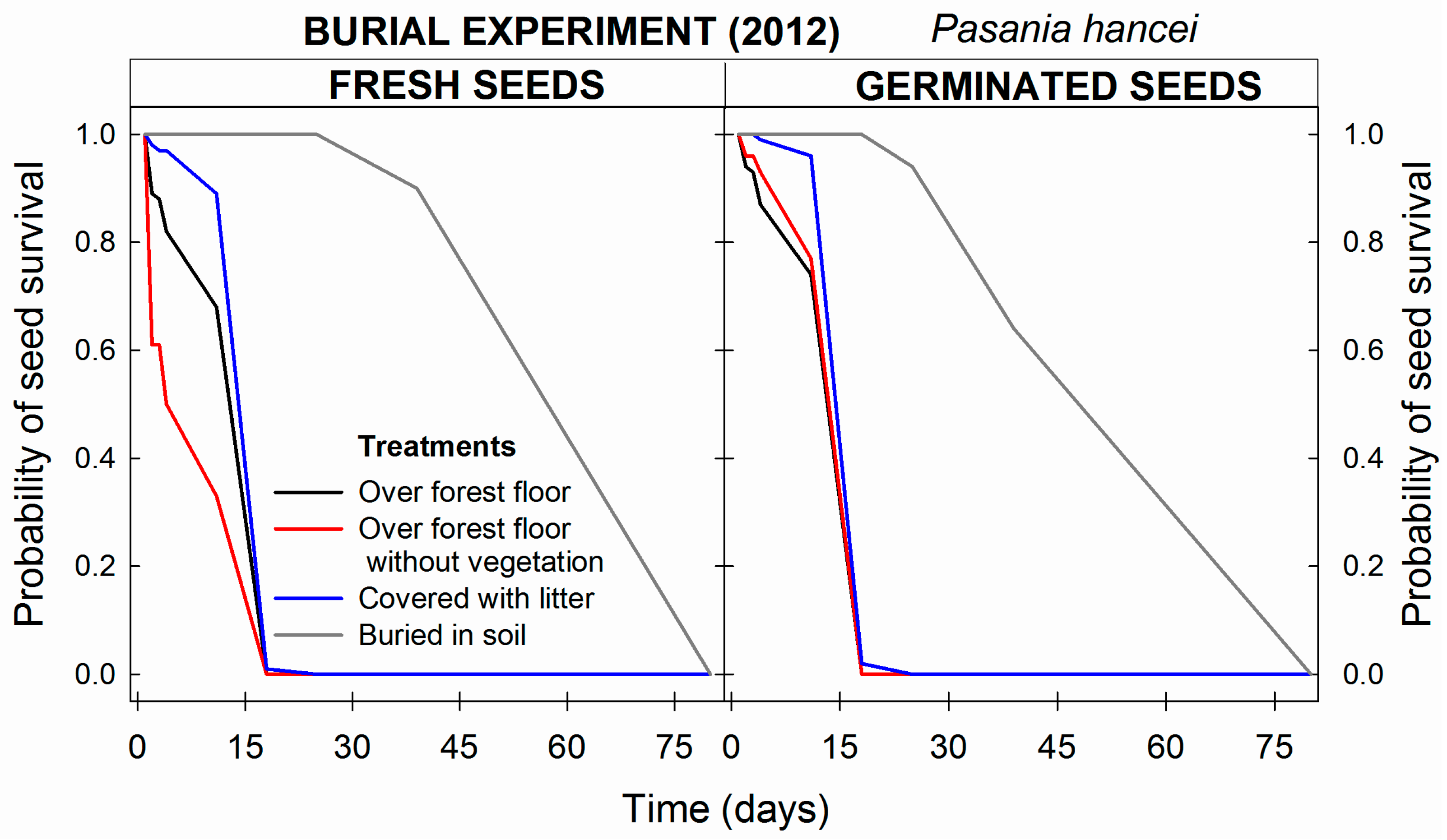
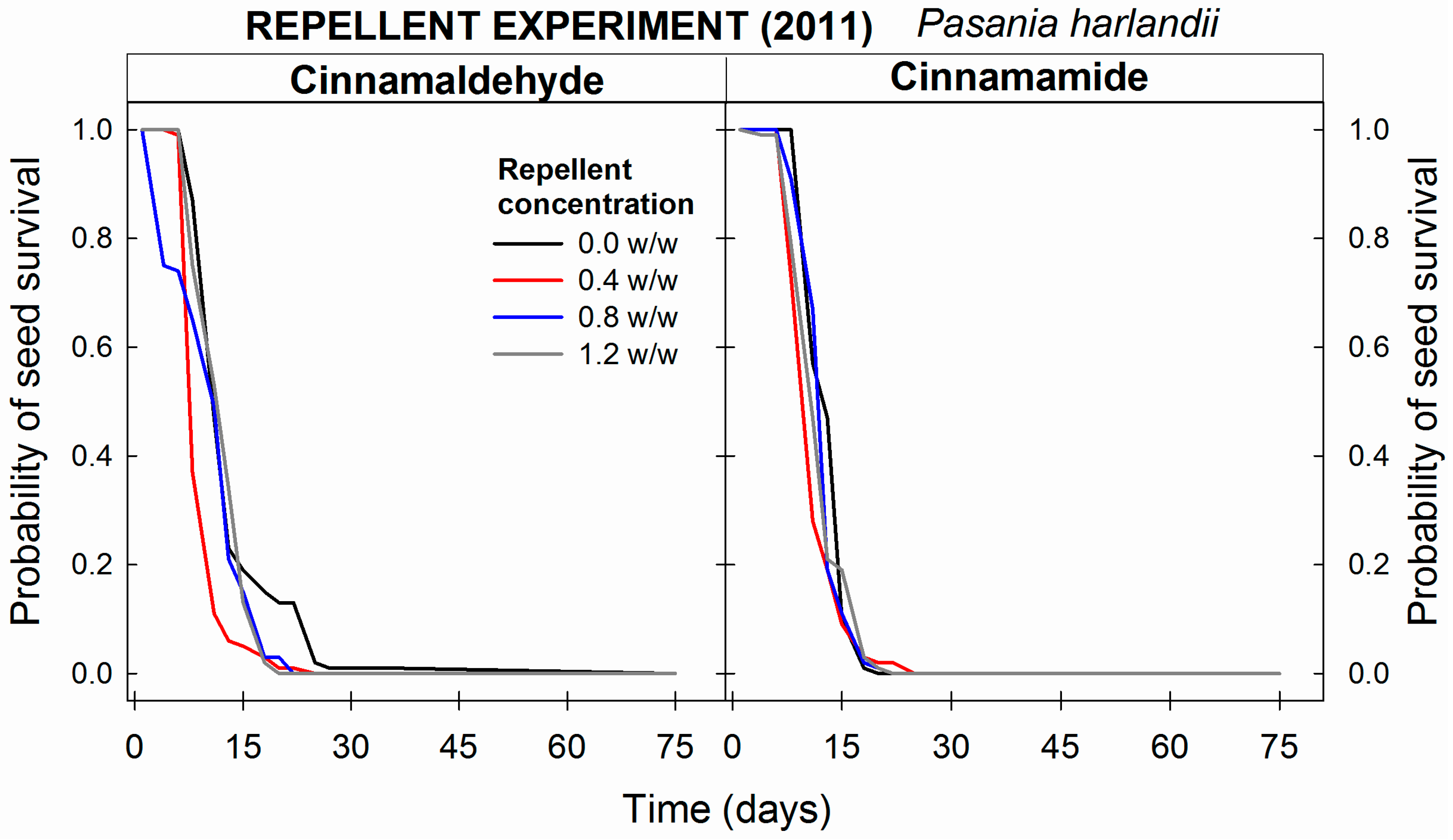
3.2.2. Seed Concealment and Chemical Repellents
3.2.3. Identification of Acorn Consumers
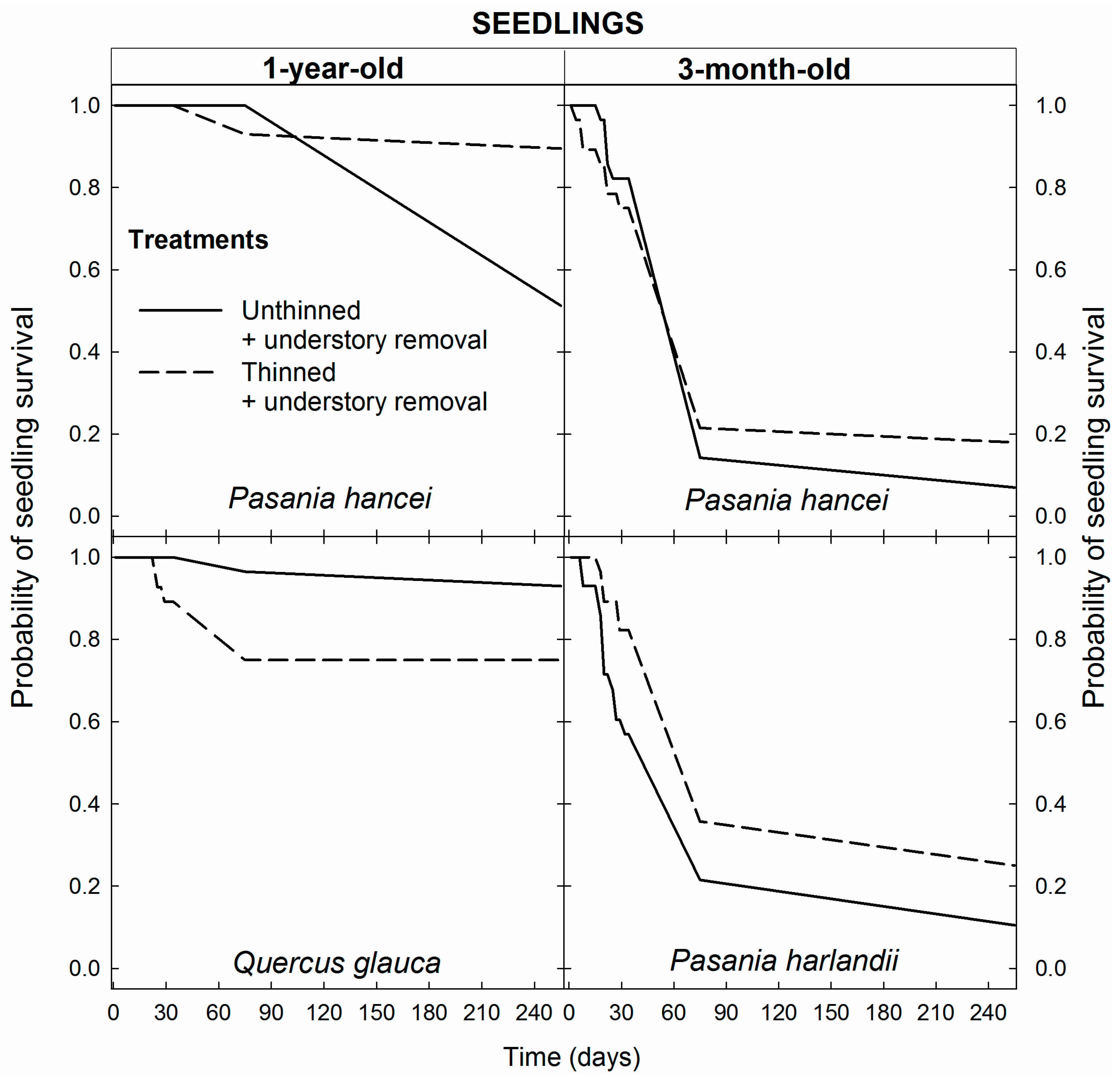
3.3. Planting Seedlings of Various Ages
4. Discussion
4.1. Seed Predation as the Main Cause of Establishment Failure
4.2. Preventing Seed Predation
4.3. Overcoming Establishment Bottlenecks: Ecosystem Management Implications
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chazdon, R.L. Beyond deforestation: Restoring forests and ecosystem services on degraded lands. Science 2008, 320, 1458–1460. [Google Scholar] [CrossRef] [PubMed]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The role of plantations in managing the world’s forests in the Anthropocene. Front. Ecol. Environ. 2010, 8, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Taiwan Forestry Bureau. Summary of the Fourth Forest Resources and Land Use Inventory in Taiwan; Taiwan Forestry Bureau: Taipei, Taiwan, 2017; p. 78.
- Cole, R.J.; Holl, K.D.; Keene, C.L.; Zahawi, R.A. Direct seeding of late-successional trees to restore tropical montane forest. For. Ecol. Manag. 2011, 261, 1590–1597. [Google Scholar] [CrossRef]
- Doust, S.J.; Erskine, P.D.; Lamb, D. Restoring rainforest species by direct seeding: Tree seedling establishment and growth performance on degraded land in the wet tropics of Australia. For. Ecol. Manag. 2008, 256, 1178–1188. [Google Scholar] [CrossRef]
- Löf, M.; Birkedal, M. Direct seeding of Quercus robur L. for reforestation: The influence of mechanical site preparation and sowing date on early growth of seedlings. For. Ecol. Manag. 2009, 258, 704–711. [Google Scholar] [CrossRef]
- Schmidt, L. A Review of Direct Sowing versus Planting in Tropical Afforestation and Land Rehabilitation; Development and Environment Series 10-2008; Forest & Landscape Denmark: Copenhagen, Denmark, 2008; p. 38. ISBN 978-87-7903-328-3. [Google Scholar]
- Zahawi, R.A.; Holl, K.D. Comparing the performance of tree stakes and seedlings to restore abandoned tropical pastures. Restor. Ecol. 2009, 17, 854–864. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds, 2nd ed.; Cambridge University Press: Cambridge, UK, 2005; p. 250. ISBN 0521653681. [Google Scholar]
- Leck, M.A.; Parker, V.T.; Simpson, R.L. Seedling Ecology and Evolution, 1st ed.; Cambridge University Press: Cambridge, UK, 2008; p. 534. ISBN 9780521694667. [Google Scholar]
- Beckage, B.; Clark, J.S.; Clinton, B.D.; Haines, B.L. A long-term study of tree seedling recruitment in southern Appalachian forests: The effects of canopy gaps and shrub understories. Can. J. For. Res. 2011, 30, 1617–1631. [Google Scholar] [CrossRef]
- Doust, S.J.; Erskine, P.D.; Lamb, D. Direct seeding to restore rainforest species: Microsite effects on the early establishment and growth of rainforest tree seedlings on degraded land in the wet tropics of Australia. For. Ecol. Manag. 2006, 234, 333–343. [Google Scholar] [CrossRef]
- Augspurger, C.K. Light requirements of neotropical tree seedlings—A comparative-study of growth and survival. J. Ecol. 1984, 72, 777–795. [Google Scholar] [CrossRef]
- Brokaw, N.; Busing, R.T. Niche versus chance and tree diversity in forest gaps. Trends Ecol. Evol. 2000, 15, 183–188. [Google Scholar] [CrossRef]
- Masaki, T.; Osumi, K.; Takahashi, K.; Hoshizaki, K.; Matsune, K.; Suzuki, W. Effects of microenvironmental heterogeneity on the seed-to-seedling process and tree coexistence in a riparian forest. Ecol. Res. 2007, 22, 724–734. [Google Scholar] [CrossRef]
- Blanco, J.A.; Imbert, J.B.; Castillo, F.J. Effects of thinning on nutrient pools in two contrasting Pinus sylvestris L. forests in the western Pyrenees. Scand. J. For. Res. 2006, 21, 143–150. [Google Scholar] [CrossRef]
- Smit, C.; Gusberti, M.; Mueller-Schaerer, H. Safe for saplings; safe for seeds? For. Ecol. Manag. 2006, 237, 471–477. [Google Scholar] [CrossRef]
- Li, Y.-T. Restoration of a Plantation Forest to Native Broadleaved Vegetation: Indentifying and Alleviating Establishment Limitations. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2013. [Google Scholar]
- Lo, Y.-H.; Lin, Y.-C.; Blanco, J.A.; Yu, C.-H.; Guan, B.T. Moving from ecological conservation to restoration: An example from central Taiwan, Asia. In Forest Ecosystems: More Than Just Trees; Blanco, J.A., Lo, Y.-H., Eds.; InTech: Rijeka, Croatia, 2012; pp. 339–354. ISBN 978-953-307-667-6. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC: Boca Raton, FL, USA, 2006; p. 476. ISBN 9781498728331. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 10 November 2017).
- Boman, J.S.; Casper, B.B. Differential postdispersal seed predation in disturbed and intact temperate forest. Am. Midl. Nat. 1995, 134, 107–116. [Google Scholar] [CrossRef]
- Schnurr, J.L.; Canham, C.D.; Ostfeld, R.S.; Inouye, R.S. Neighborhood analyses of small-mammal dynamics: Impacts on seed predation and seedling establishment. Ecology 2004, 85, 741–755. [Google Scholar] [CrossRef]
- Chambers, J.C.; MacMahon, J.A. A day in the life of a seed: Movements and fates of seeds and their implications for natural and managed systems. Ann. Rev. Ecol. Syst. 1994, 25, 263–292. [Google Scholar] [CrossRef]
- Den Ouden, J.; Jansen, P.A.; Smit, R. Jays, mice and oaks: Predation and dispersal of Quercus robur and Q. petraea in north-western Europe. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.M., Lambert, J.E., Hulme, P.E., vander Wall, S.B., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 223–240. ISBN 0851998062. [Google Scholar]
- Hulme, P.E.; Kollmann, J. Seed predator guilds, spatial variation in post-dispersal seed predation and potential effects on plant demography—A temperate perspective. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.M., Lambert, J.E., Hulme, P.E., vander Wall, S.B., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 9–30. ISBN 0851998062. [Google Scholar]
- Tamura, N.; Katsuki, T. Walnut seed dispersal: Mixed effects of tree squirrels and field mice with different hoarding ability. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.M., Lambert, J.E., Hulme, P.E., vander Wall, S.B., Eds.; CABI publishing: Wallingford, UK, 2005; pp. 241–252. ISBN 0851998062. [Google Scholar]
- Schupp, E.W.; Milleron, T.; Russo, S.E. Dissemination limitation and the origin and maintenance of species-rich tropical forests. In Seed Dispersal and Frugivory: Ecology, Evolution and Conservation; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CABI International: Wallingford, UK, 2002; pp. 19–34. ISBN 085199525X. [Google Scholar]
- González-Rodríguez, V.; Villar, R. Post-dispersal seed removal in four Mediterranean oaks: Species and microhabitat selection differ depending on large herbivore activity. Ecol. Res. 2012, 27, 587–594. [Google Scholar] [CrossRef]
- Tsai, J.-W.; Yuan, H.-W.; Tsai, P.-Y.; Lee, S.-Y.; Ding, T.-S.; Hong, C.-H. Effects of thinning on bird community and spinous country-rat (Niviventer coxingi) population in china-fir (Cunninghamia lanceolata) plantations. Q. J. Chin. For. 2010, 43, 367–382. [Google Scholar]
- Li, L.-L. 1981 A Study on the Behavior of Red-bellied Squirrel (Callosciurus erythraeus). Master’s Thesis, National Taiwan University, Taipei, Taiwan, 1981. [Google Scholar]
- Zhang, Z.-B.; Xiao, Z.-S.; Li, H.-J. Impact of small rodents on tree seeds in temperate and subtropical forests, China. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.M., Lambert, J.E., Hulme, P.E., Vander Wall, S.B., Eds.; CABI publishing: Wallingford, UK, 2005; pp. 269–282. ISBN 0851998062. [Google Scholar]
- Vander Wall, S.B. Seed fate pathways of antelope bitterbrush: Dispersal by seed-caching yellow pine chipmunks. Ecology 1994, 75, 1911–1926. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhang, Z.; Wang, Y. Effects of enclosure protection and seed burial on direct seeding of nut-bearing trees. Biodivers. Sci. 2005, 13, 520–526. [Google Scholar] [CrossRef]
- Chang, C.T. Microenvironmental Variation, Seed Germination and Seedling Growth under Different Canopy Openness in a Sugi (Cryptomeria japonica) Plantation at Shitou, Central Taiwan. Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2007. [Google Scholar]
- Löf, M.; Bergquist, J.; Brunet, J.; Karlsson, M.; Welander, N.T. Conversion of Norway spruce stands to broadleaved woodland-regeneration systems, fencing and performance of planted seedlings. Ecol. Bull. 2010, 53, 165–173. [Google Scholar]
- Xiao, Z.S.; Zhang, Z.B.; Wang, Y.S. Dispersal and germination of big and small nuts of Quercus Serrata in a subtropical broad-leaved evergreen forest. For. Ecol. Manag. 2004, 195, 141–150. [Google Scholar] [CrossRef]
- Xiao, Z.S.; Zhang, Z.B.; Wang, Y.S. Effects of seed size on dispersal distance in five rodent-dispersed Fagaceous species. Acta Oecol. 2005, 28, 221–229. [Google Scholar] [CrossRef]
- Birkedal, M.; Löf, M.; Olsson, G.E.; Bergsten, U. Effects of granivorous rodents on direct seeding of oak and beech in relation to site preparation and sowing date. For. Ecol. Manag. 2010, 259, 2382–2389. [Google Scholar] [CrossRef]
- Savadogo, P.; Tigabu, M.; Oden, P.C. Restoration of former grazing lands in the highlands of Laos using direct seeding of four native tree species. Mt. Res. Dev. 2010, 30, 232–243. [Google Scholar] [CrossRef]
- Huang, I.-C.; Chen, I.-Z.; Lu, S.-Y.; Chang, K.-S. Effects of chilling stratification, scarification and excised-embryo on seed germination of Lithocarpus lepidocarpus Hayata. J. Chin. Soc. Hortic. Sci. 2004, 50, 515–520. [Google Scholar]
- Crocker, D.R.; Scanlon, C.B.; Perry, S.M. Repellency and Choice: Feeding responses of wild rats (Rattus norvegicus) to cinnamic acid derivatives. Appl. Anim. Behav. Sci. 1993, 38, 61–66. [Google Scholar] [CrossRef]
- Gurney, J.E.; Watkins, R.W.; Gill, E.L.; Cowan, D.P. Non-lethal mouse repellents: Evaluation of cinnamamide as a repellent against commensal and field Rodents. Appl. Anim. Behav. Sci. 1996, 49, 353–363. [Google Scholar] [CrossRef]
- Lee, H.K.; Lee, H.S.; Ahn, Y.J. Antignawing factor derived from Cinnamomum Cassia bark against mice. J. Chem. Ecol. 1999, 25, 1131–1139. [Google Scholar] [CrossRef]
- Chao, J.-T.; Fang, K.-Y.; Koh, C.-N.; Chen, Y.-M.; Yeh, W.-C. Feeding on plants by the red-bellied tree squirrel Callosciurus erythraeus in Taipei Botanical Garden. Bull. Taiwan For. Res. Inst. 1993, 8, 39–50. [Google Scholar]
- Liu, Y.-F. A Study on the Population and Habitat Use of the Red-bellied Squirrel (Callosciurus erythraeus) in Nanjenshan Area. Master’s Thesis, National Pingtung University of Science and Technology, Pingtung, Taiwan, 2003. [Google Scholar]
- Li, H.J.; Zhang, Z.B. Effect of rodents on acorn dispersal and survival of the Liaodong oak (Quercus liaotungensis Koidz.). For. Ecol. Manag. 2003, 176, 387–396. [Google Scholar] [CrossRef]
- Birkedal, M.; Fischer, A.; Karlsson, M.; Löf, M.; Madsen, P. Rodent impact on establishment of direct-seeded Fagus sylvatica, Quercus robur and Quercus petraea on forest land. Scand. J. For. Res. 2009, 24, 298–307. [Google Scholar] [CrossRef]
- Buckley, D.A.; Sharik, T.L. Effect of overstorey and understorey vegetation treatments on removal of planted northern red oak acorns by rodents. North. J. Appl. For. 2002, 19, 88–92. [Google Scholar]
- Blanco, J.A.; Welham, C.; Kimmins, J.P.; Seely, B.; Mailly, D. Guidelines for modeling natural regeneration in boreal forests. For. Chron. 2009, 85, 427–439. [Google Scholar] [CrossRef]
- Liu, T.-Y.; Lin, K.-C.; Vadeboncoeur, M.A.; Chen, M.-A.; Huang, M.-Y.; Lin, T.C. Undersotey plant community and ligth availability in conifer plantations and natural hardwood forests in Taiwan. Appl. Veg. Sci. 2015, 18, 591–602. [Google Scholar] [CrossRef]
- Bi, J.; Blanco, J.A.; Kimmins, J.P.; Ding, Y.; Seely, B.; Welham, C. Yield decline in Chinese fir plantations: A simulation investigation with implications for model complexity. Can. J. For. Res. 2007, 37, 1615–1630. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Lithocarpus lepidocarpus (Hayata) Hayata | Quercus glauca (Thunberg) Oersted | Pasania kawakami (Hayata) Hayata | Pasania hancei (Benth.) Schottky | Pasania harlandii (Hance ex Walp.) Oerst. |
|---|---|---|---|---|---|
| Direct seeding | |||||
| Fresh seeds | 2009, 2011 | 2009 | 2011, 2012 | ||
| Germinated seeds | 2011 | 2010 | 2010, 2011, 2012 | ||
| Controlling seed predation | |||||
| Fencing | 2010 | 2011 1 | |||
| Seed concealment | 2011 | 2012 | |||
| Chemical repellent | 2012 | 2011 | |||
| Planting seedlings | |||||
| 3-month-old | 2011 | 2011 | |||
| 1-year-old | 2011 | 2011 |
| Treatment | L. lepidocarpus | P. kawakamii | ||||
|---|---|---|---|---|---|---|
| df | χ2 | p | df | χ2 | p | |
| Understory control | 1 | 0.091 | 0.763 | 1 | 0.748 | 0.387 |
| Thinning | 1 | 1.176 | 0.278 | 1 | 0.238 | 0.625 |
| Understory control × thinning | 1 | 0.856 | 0.355 | 1 | 1.308 | 0.253 |
| Treatment | Q. glauca | P. hancei | ||||
|---|---|---|---|---|---|---|
| df | χ2 | p | df | χ2 | p | |
| Understory control | 1 | 1.128 | 0.288 | 1 | 4.001 | 0.0455 |
| Thinning | 1 | 0.303 | 0.582 | 1 | 0.010 | 0.9203 |
| Understory control × thinning | 1 | 0.435 | 0.509 | 1 | 0.109 | 0.7414 |
| Treatment 1 | L. lepidocarpus | P. hancei var. ternaticupula | ||||
|---|---|---|---|---|---|---|
| df | χ2 | p | df | χ2 | p | |
| Understory control | 1 | 2.784 | 0.0952 | 1 | 2.975 | 0.0846 |
| Thinning | 1 | 0.057 | 0.8115 | 1 | 11.341 | 0.0008 |
| Type of seeds (fresh or germinated) | 1 | 0.460 | 0.4979 | 1 | 3.441 | 0.0636 |
| Treatment 1 | P. hancei var. ternaticupula | ||
|---|---|---|---|
| df | χ2 | p | |
| Method | 1 | 11.884 | 0.0078 |
| Germination phase | 1 | 0.446 | 0.5043 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-T.; Lo, Y.-H.; Lin, Y.-C.; Guan, B.T.; Blanco, J.A.; You, C.-H. Bringing the Natives Back: Identifying and Alleviating Establishment Limitations of Native Hardwood Species in a Conifer Plantation. Forests 2018, 9, 3. https://doi.org/10.3390/f9010003
Li Y-T, Lo Y-H, Lin Y-C, Guan BT, Blanco JA, You C-H. Bringing the Natives Back: Identifying and Alleviating Establishment Limitations of Native Hardwood Species in a Conifer Plantation. Forests. 2018; 9(1):3. https://doi.org/10.3390/f9010003
Chicago/Turabian StyleLi, Yu-Tsen, Yueh-Hsin Lo, Yi-Ching Lin, Biing T. Guan, Juan A. Blanco, and Chi-How You. 2018. "Bringing the Natives Back: Identifying and Alleviating Establishment Limitations of Native Hardwood Species in a Conifer Plantation" Forests 9, no. 1: 3. https://doi.org/10.3390/f9010003