
Abstract
We employ size-based theoretical arguments to derive simple analytic predictions of ecological patterns and properties of natural communities: size-spectrum exponent, maximum trophic level, and susceptibility to invasive species. The predictions are brought about by assuming that an infinite number of species are continuously distributed on a size–trait axis. It is, however, an open question whether such predictions are valid for a food web with a finite number of species embedded in a network structure. We address this question by comparing the size-based predictions to results from dynamic food web simulations with varying species richness. To this end, we develop a new size- and trait-based food web model that can be simplified into an analytically solvable size-based model. We confirm existing solutions for the size distribution and derive novel predictions for maximum trophic level and invasion resistance. Our results show that the predicted size-spectrum exponent is borne out in the simulated food webs even with few species, albeit with a systematic bias. The predicted maximum trophic level turns out to be an upper limit since simulated food webs may have a lower number of trophic levels, especially for low species richness, due to structural constraints. The size-based model possesses an evolutionary stable state and is therefore un-invadable. In contrast, the food web simulations show that all communities, irrespective of number of species, are equally open to invasions. We use these results to discuss the validity of size-based predictions in the light of the structural constraints imposed by food webs.








Similar content being viewed by others

References
Andersen KH, Beyer JE (2006) Asymptotic body size determines species abundance in the marine size spectrum. Am Nat 168:54–61
Andersen KH, Beyer JE, Lundberg P (2009) Trophic and individual efficiencies of size-structured communities. Proc R Soc B 276:109–114
Barnes C, Maxwell D, Reuman D, Jennings S (2010) Global patterns in predator-prey size relationships reveal size dependency of trophic transfer efficiency. Ecology 91:222–232
Benoît E, Rochet MJ (2004) A continuous model of biomass size spectra governed by predation and the effects of fishing on them. J Theor Biol 226:9–21
Boit A, Martinez ND, Williams RJ, Gadeke U (2012) Mechanistic theory and modelling of complex food-web dynamics in Lake Constance. Ecol Lett 15:594–602
Borgmann U (1987) Models on the slope of, and biomass flow up, the biomass size spectrum. Can J Fish Aquat Sci 44:136–140
Brose U, Williams RJ, Martinez ND (2006a) Allometric scaling enhances stability in complex food webs. Ecol Lett 9:1228–1236
Brose U, Jonsson T, Berlow EL, Warren P, Banasek-Richter C, Bersier LF, Blanchard JL, Brey T, Carpenter SR, Blandenier MF, Cushing L, Dawah HA, Dell T, Edwards F, Harper-Smith S, Jacob U, Ledger ME, Martinez ND, Memmott J, Mintenbeck K, Pinnegar JK, Rall BC, Rayner TS, Reuman DC, Ruess L, Ulrich W, Williams RJ, Woodward G, Cohen JE (2006b) Consumer-resource body-size relationships in natural food webs. Ecology 87:2411–2417
Chesson P, Kuang JJ (2008) The interaction between predation and competition. Nature 456:235–238
Cohen JE, Pimm SL, Yodzis P, Saldaña J (1993) Body sizes of animal predators and animal prey in food webs. J Anim Ecol 62:67–78
Datta S, Delius GW, Law R, Plank MJ (2011) A stability analysis of the power-law steady state of marine size spectra. J Math Biol 63:779–799
Drake JA (1990) The mechanics of community assembly and succession. J Theor Biol 147:213–233
Giacomini HC, DeMarco P, Petrere M (2009) Exploring community assembly through an individual-based model for trophic interactions. Ecol Model 220:23–39
Gomez-Canchong P, Quinones RA, Brose U (2012) Robustness of size-structure across ecological networks in pelagic systems. Theor Ecol 6:45–56
Hartvig M, Andersen KH , Beyer JE (2011) Food web framework for size-structured populations. J Theor Biol 272:113–122
Hartvig M (2011) Ecological processes yield complex and realistic food webs. In: Food web ecology—individual life-histories and ecological processes shape complex communities. ISBN 978-91-7473-080-7. Ph.D. thesis. Department of Biology. Lund University, Sweden, pp 75–126
Haskell J, Ritchie M, Olff H (2002) Fractal geometry predicts varying body size scaling relationships for mammal and bird home ranges. Nature 418:527–530
Jetz W, Carbone C, Fulford J, Brown JH (2004) The scaling of animal space use. Science 306:266–268
Kerr S (1974) Theory of size distribution in ecological communities. J Fish Res Board Can 31:1859–1862
Kramer D, Chapmanm M (1999) Implications of fish home range size and relocation for marine reserve function. Environ Biol Fish 55:65–79
Law R, Morton RD (1996) Permanence and the assembly of ecological communities. Ecology 77:762–775
Law R (1999) Theoretical aspects of community assembly. In: McGlade J (ed) Advanced ecological theory. Blackwell Science, Oxford, pp 143–171
Levine S (1980) Several measures of trophic structure applicable to complex food webs. J Theor Biol 83:195–207
Lewis HM, Law R (2007) Effects of dynamics on ecological networks. J Theor Biol 247:64–76
Lindeman RL (1942) The trophic-dynamic aspect of ecology. Ecology 4:399–417
Loeuille N, Loreau M (2005) Evolutionary emergence of size-structured food webs. Proc Natl Acad Sci USA 102:5761–5766
MacArthur R, Levins R (1967) The limiting similarity, convergence, and divergence of coexisting species. Am Nat 101:377–385
Maury Q, Poggiale JC (2013) From individuals to populations to communities: a dynamic energy budget model of marine ecosystem size-spectrum including life history diversity. J Theor Biol . doi:10.1016/j.jtbi.2013.01.018
Maynard Smith J, Price GR (1973) The logic of animal conflict. Nature 246:15–18
Metz JAJ, Nisbet RM, Geritz SAH (1992) How should we define “fitness” for general ecological scenarios?Tree 7:198–202
Morton RD, Law R (1997) Regional species pools and the assembly of local ecological communities. J Theor Biol 187:321–331
Odum WE, Heald EJ (1975) The detritus-based food web of an estuarine mangrove community. In: Cronin L (ed) Estuarine research. Chemistry biology and the estuarine system, vol 1. Academic, London
Oksanen L, Fretwell SD, Arruda J, Niemela P (1981) Exploitation ecosystems in gradients of primary productivity. Am Nat 118:240–261
Post WM, Pimm SL (1983) Community assembly and food web stability. Math Biosci 64:169–182
Post DM (2002) The long and short of food-chain length. Trends Ecol Evol 17:269–277
Rooney N, McCann K, Gellner G, Moore JC (2006) Structural asymmetry and the stability of diverse food webs. Nature 442:265–269
Reuman DC, Mulder C, Raffaelli D, Cohen JE (2008) Three allometric relations of population density to body mass: theoretical integration and empirical tests in 149 food webs. Ecol Lett 11:1216–1228
Rossberg AG, Ishi R, Amemiya T, Itoh K (2008) The top-down mechanism for body-mass-abundance scaling. Ecology 89:567–580
Rossberg AG, Brännström Å, Dieckmann U (2010) How trophic interaction strength depends on traits. Theor Ecol 3:13–24
Rossberg AG (2012) A complete analytic theory for structure and dynamics of populations and communities spanning wide ranges in body size. Adv Ecol Res 46:429–522
Savage VM, Gillooly JF, Brown JH, West GB, Charnov EL (2004) Effects of body size and temperature on population growth. Am Nat 163:429–441
Sheldon RW, Prakash A, Sutcliffe WHJ (1972) The size distribution of particles in the ocean. Limnol Oceanogr 17:327–340
Sheldon RW, Sutcliffe WHJ, Paranjape MA (1977) Structure of pelagic food chain and relationship between plankton and fish production. J Fish Res Board Can 34:2344–2353
Taylor PJ (1988) The construction and turnover of complex community models having generalized Lotka-Volterra dynamics. J Theor Biol 135:569–588
Ursin E (1973) On the prey size preferences of cod and dab. Meddelelser fra Dammarks Fiskeri- og Havundersøgelser 7:84–98
Virgo N, Law R, Emmerson M (2006) Sequentially assembled food webs and extremum principles in ecosystem ecology. J Anim Ecol 75:377–386
Ware DM (1978) Bioenergetics of pelagic fish: theoretical change in swimming speed and ration with body size. J Fish Res Board Can 35:220–228
West GB, Brown JH, Enquist BJ (1997) A general model for the origin of allometric scaling laws in biology. Science 276:122–126
Yodzis P (1984) Energy flow and the vertical structure of real ecosystems. Oecologia 65:86–88
Yodzis P, Innes S (1992) Body size and consumer–resource dynamics. Am Nat 6:1151–1175
Acknowledgments
Lai Zhang gratefully acknowledges the financial support from the Swedish Kempe Foundation. Martin Hartvig acknowledges the Danish National Research Foundation for support to the Center for Macroecology, Evolution and Climate. Ken Haste Andersen was financially supported by the European FP7 program MEECE and the VKR center of excellence Ocean Life.
Author information
Authors and Affiliations
Corresponding author
Appendix: Assembly algorithm
Appendix: Assembly algorithm
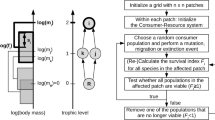
The assembly algorithm is a replicate of that by Hartvig (2011), but presented below for completeness.
Model communities are formed using sequential assembly by introducing one new species at a time in low density (\(10^{-10}\) g/vol) (Post and Pimm 1983; Drake 1990; Law 1999) from a species pool. If invasion fitness is positive, then the system is stimulated till it reaches steady state which can be a fixed point, periodic, or even chaotic, detected using heuristic algorithm. Fitness is measured using the per capita population growth rate (i.e., the evaluation of the right-hand side of Eq. (5)). A species is assumed to be in steady state if its absolute fitness is smaller than \(1/1{,}000\) year−1. During simulation, species are removed if they are going to extinction, defined as (1) population biomass falls below the extinction threshold \(10^{-20}\) g/vol; (2) fitness is smaller than \(-1/250\) year−1, while the biomass is below \(10^{-5}\) g/vol; or (3) fitness is smaller than \(-1/1{,}000\) year−1, while the biomass is below \(10^{-10}\) g/vol. The assembly proceeds to a new invader if the introduced invader has negative invasion fitness, or if the augmented community (resident community plus invader) has reached equilibrium state.
In addition, the continuous species pool is discretized evenly in the x direction with a step size \(\delta \log w = 0.1\). Results appear independent of the choice of discretization.
Rights and permissions
About this article
Cite this article
Zhang, L., Hartvig, M., Knudsen, K. et al. Size-based predictions of food web patterns. Theor Ecol 7, 23–33 (2014). https://doi.org/10.1007/s12080-013-0193-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12080-013-0193-5